自噬在葛根素促成骨细胞分化调节中的作用
2021-09-28于冬冬赵丹阳杨鸫祥
于冬冬,赵丹阳,杨鸫祥
(1.辽宁中医药大学附属医院骨科,沈阳 110032;2.沈阳市第一人民医院神经内科,沈阳 110041)
成骨细胞的分化受到抑制是骨质疏松症发生的主要机制之一[1-2]。近年来,应用中药的活性成份调节成骨细胞分化,进而达到防治骨质疏松症的目的越来越受到人们的重视,如淫羊藿苷、丹参酮-ⅡA和骨碎补等[3-5]。葛根素作为中草药葛根的活性成分,能增加成骨细胞的增殖和碱性磷酸酶(alkaline phosphatase,ALP)的活性、促进矿化结节的形成,促进机体骨密度和骨矿物质含量增加[6]。本课题组前期研究[1,7-8]发现,葛根素能够抑制糖皮质激素诱导的成骨细胞凋亡,同时抑制破骨细胞的分化。
自噬是维持细胞正常代谢必需的一个生理过程[9]。自噬与骨质疏松症关联密切,敲除自噬关键基因的模型大鼠骨量下降,发生骨质疏松症[10]。自噬缺乏的成骨细胞矿化能力降低[11],提示自噬在骨形成和成骨细胞分化过程中发挥重要的调节作用。
葛根素能够促进成骨细胞的分化及矿化,但是其作用机制目前还不完全清楚。本研究拟探讨自噬在葛根素调节成骨细胞分化过程中的作用,为葛根素应用于临床防治骨质疏松症提供依据。
1 材料与方法
1.1 材料
1.1.1 细胞:hFOB1.19人成骨细胞(美国ATCC公司)。
1.1.2 试剂:葛根素(中国狄尔格生物医药公司,溶于DMSO,储存浓度1×10-5mol/L,-20 ℃保存);进口胎牛血清(美国Hychone公司);BCIP/NBT ALP显色试剂盒(中国Beyotime公司),茜素红S(美国Sigma公司),免疫荧光染色试剂盒、山羊抗兔、山羊抗鼠二抗HRP(美国Abcom公司),Cy3标记的二抗(中国Beyotime公司);Runt相关转录因子2(runt-related transcription factor 2,RUNX 2),骨保护素(osteoprotegerin,OPG)、微管相关蛋白轻连3-Ⅱ/Ⅰ(microtubule associated protein light chain 3-Ⅱ/Ⅰ,LC3-Ⅱ/Ⅰ)、BECLIN 1一抗(美国Abcom公司);G418(美国Sigma公司);3-甲基腺嘌呤(3-methyladenine,3-MA)。
1.1.3 仪器:细胞孵箱(日本Thermo公司);倒置相差显微镜(日本Olympus公司);免疫荧光显微镜(日本Olympus公司);AMR-100酶标仪(中国杭州奥盛仪器公司)。
1.2 方法
1.2.1 hFOB1.19成骨细胞培养、诱导分化及分组:
1.2.1.1 未分化的成骨细胞培养 未分化的成骨细胞培养基含DMEM/F-12、10%胎牛血清、0.3 g/L G418,孵箱培养条件为5%CO2、33 ℃。
1.2.1.2 诱导分化培养 分化培养基是在未分化培养基的基础上,加入β-甘油磷酸(5×10-3mol/L),抗坏血酸(0.1 g/L),维生素K(10-8mol/L ),1,25-二羟维生素D3[1,25(OH)2D3](10-7mol/L),孵箱培养条件为5%CO2,39.4 ℃。
1.2.1.3 分组 将成骨细胞分为3组,即A组(成骨分化诱导),B组(成骨分化诱导+10-8mol/L葛根素3 h),C组(成骨分化诱导+10-8mol/L葛根素3 h+5 mmol/L 3-MA 1 h)。
1.2.2 ALP分析:细胞分组处理后,PBS洗涤3次,用95%乙醇固定细胞10 min,晾干,按照碧云天的BCIP/NBT碱性磷酸酶显色试剂盒所提供的相应操作步骤进行染色分析,晾干拍照。
1.2.3 茜素红染色:细胞(1×105/孔)接种于24孔板内生长致融合,分组处理后75%乙醇固定和1%的茜素红S染色,显微镜下观察并拍照。
1.2.4 免疫荧光染色检测成骨分化过程中LC3Ⅰ/Ⅱ的蛋白表达:将圆形的多聚赖氨酸载玻片至于6孔板内,细胞接种于多聚赖氨酸载玻片上24 h。分组处理后,用免疫染色固定液固定爬片10 min,免疫染色洗涤液洗玻片3次,每次3 min,封闭液封闭60 min;一抗并放入湿盒,4 ℃孵育过夜。去除一抗,加荧光二抗,湿盒中37 ℃孵育1 h,滴加DAPI避光孵育5 min,对标本进行染核,用含抗荧光淬灭剂的封片液封片,荧光显微镜下观察采集图像。
1.2.5 透射电镜观察:细胞经分组处理后消化、离心,去上清,将离心管中的细胞团块取出移入1.5 mL的EP管内,在4 ℃用PBS漂洗3次,每次10 min。再用1% 四氧化锇在4 ℃固定15~30 min,接着用PBS漂洗3次。丙酮溶液梯度脱水后浸透,吸弃瓶中脱水剂,加3 mL纯丙酮-EPON812包埋剂(1 ∶1体积比),室温下放置30 min后,弃去稀释的包埋剂,加纯包埋剂1 mL,室温放置2 h或过夜。将预先干燥的EPON明胶囊注满混合包埋剂,倒盖在单层细胞上,在60 ℃固化24 h。超薄切片,采用JEOL 2000FX透射电子显微镜观察。对自噬性溶酶体空泡样量化,自噬性溶酶体空泡样的计数在每个领域和标准化的表面积(n> 3)。
1.2.6 Western blotting检测:加入裂解液提取细胞蛋白,冰上放置40 min,4 ℃ 12 000 r/min离心40 min,取上清,BCA法定量蛋白。取20 μg蛋白样品,于10%聚丙烯酰胺凝胶上行SDS-PAGE,电泳结束后100 V电转印至PVDF膜。将PVDF膜放入封闭液中37 ℃封闭1 h,加入稀释一抗4 ℃孵育过夜。PBS洗膜3次,加入辣根过氧化物酶(horseradish peroxidase,HRP)标记的稀释二抗孵育,37 ℃震摇1 h,洗膜。ECL发光法曝光成像,扫描入电脑(n> 3)。
1.3 统计学分析
采用 SPSS 13.0软件进行统计分析,数据以表示,组间比较采用ANOVA方法,P< 0.05为差异有统计学意义。
2 结果
2.1 葛根素促进成骨细胞分化
不同浓度的葛根素(10-6、10-7、10-8mol/L)处理诱导分化组细胞3 h后,都可见不同程度的促成骨细胞分化作用。结果显示,未处理组细胞ALP染色阳性面积约占视野的40%,10-6mol/L葛根素组ALP染色阳性面积约占视野的55%,10-7mol/L葛根素组ALP染色阳性面积约占视野的65%,10-8mol/L葛根素组ALP染色阳性面积约占视野的80%以上,见图1。葛根素在10-8mol/L浓度时促成骨细胞分化效果最明显(P=0.002),因此,本研究选用该浓度进行后续实验。
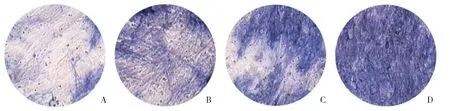
图1 葛根素促进成骨细胞分化 ×40Fig.1 Puerarin promotes osteoblast differentiation ×40
2.2 3-MA抑制葛根素的促进成骨细胞分化
ALP染色结果显示,非诱导组细胞分化不明显,A组细胞开始成骨细胞分化,ALP染色阳性面积约占视野的30%。B组成骨细胞分化效果显著提升,ALP染色阳性面积约占视野的80%(P=0.004),C组成骨细胞分化程度明显高于对照组(P=0.021),但明显低于葛根素处理的B组,ALP染色阳性面积约占视野的50%,见图2。
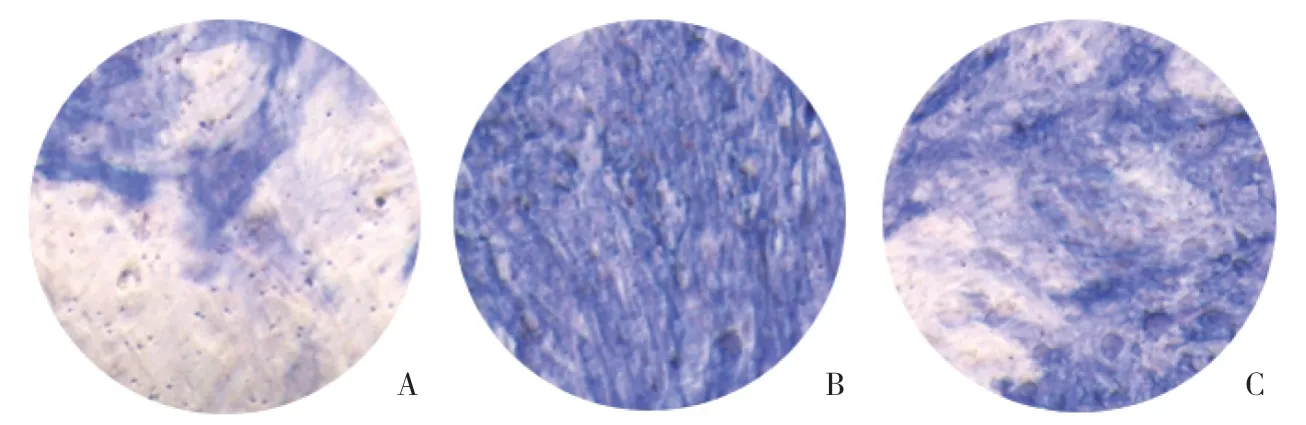
图2 3-MA抑制葛根素促进成骨细胞分化作用 ×40Fig.2 Inhibition of puerarin’s osteogenic effect by 3-MA ×40
2.3 葛根素促进成骨细胞矿化
茜素红染色结果显示,A组细胞有钙化结节的形成,茜素红染色阳性面积约占视野的25%。B组钙化结节的形成更加显著(P=0.012),茜素红染色阳性面积约占视野的50%。C组促成骨细胞分化受到抑制(P=0.034),茜素红染色阳性面积约占视野的30%,见图3。
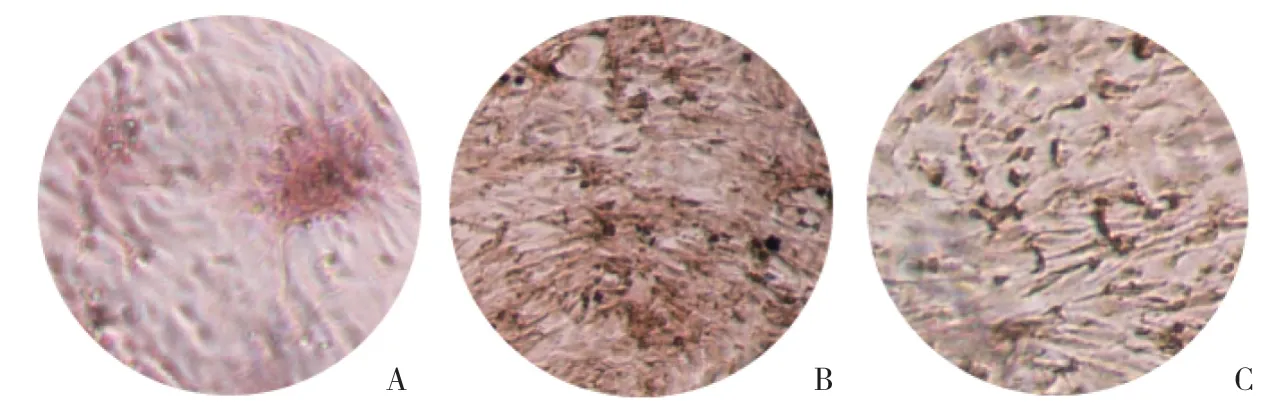
图3 葛根素促进成骨细胞矿化 ×40 Fig.3 Puerarin promotes osteoblast mineralization ×40
2.4 葛根素促进成骨细胞分化过程中自噬体的形成
A组有少量自噬体形成,B组自噬体形成较A组增多(P=0.002)。C组用3-MA自噬抑制剂处理后,自噬体明显减少,在自噬受到抑制的同时,葛根素的促成骨细胞分化效果也受到抑制(P=0.024),见图4。

图4 葛根素促进成骨细胞分化过程中自噬体的形成Fig.4 Puerarin promotes autophagy during osteoblast differentiation
2.5 葛根素促进LC3-Ⅱ蛋白表达
免疫荧光染色结果显示,未加葛根素处理时,LC3-Ⅱ散在分布于细胞质中,加入葛根素后,LC3-Ⅱ增多并在细胞核周围累积(染色红色),说明LC3-Ⅱ的表达和分布通过葛根素给药明显放大。用3-MA自噬抑制剂(5 mm,1 h)处理后,LC3-Ⅱ表达减少,见图5。
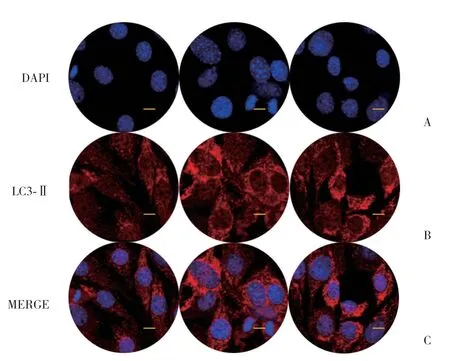
图5 葛根素促进LC3-Ⅱ蛋白表达 Fig.5 Puerarin promotes LC3-Ⅱ protein expression
2.6 葛根素促进成骨细胞分化相关蛋白表达
Western blotting结果显示,成骨细胞分化过程中,A组OPG、RUNX 2蛋白表达略增加,B组OPG、RUNX 2蛋白表达明显增加(P=0.012和P=0.38)。C组葛根素的促成骨细胞分化作用被明显减弱,其蛋白表达与A组相似。提示葛根素在促进成骨细胞分化过程中,成骨细胞分化相关蛋白发挥了重要的调节作用,见图6。
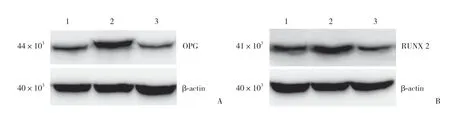
图6 葛根素促进成骨细胞分化相关蛋白表达Fig.6 Puerarin promotes autophagy during osteoblast differentiation
2.7 葛根素促进成骨细胞分化过程中自噬相关蛋白表达
A组LC3-Ⅱ蛋白的表达略有增加(P=0.001),B组LC3-Ⅱ的表达显著增加。BECLIN 1在自噬开始的信号传导中起重要作用,与A组相比,B组BECLIN 1的表达也显著增加(P=0.004)。C组葛根素的促成骨细胞分化作用被明显减弱,自噬相关蛋白表达亦减弱。提示在葛根素促进成骨细胞分化过程中,自噬发挥了重要的调节作用(P=0.014和P=0.028),见图7。
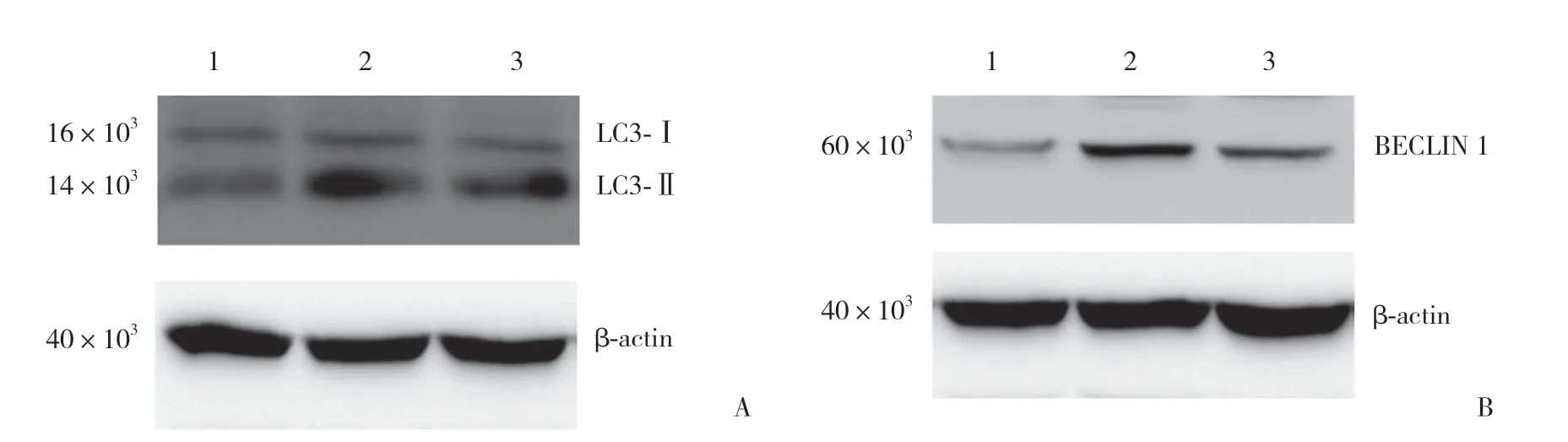
图7 葛根素促进成骨细胞分化过程中自噬相关蛋白表达Fig.7 Puerarin promotes expression of autophagy-related proteins during osteoblast differentiation
3 讨论
骨质疏松症的发病过程中伴随着成骨细胞和破骨细胞的数量和活性的不平衡[12]。其中,成骨细胞的分化能力降低是导致骨质疏松症的主要原因之一。成骨细胞是骨形成的主要细胞,因此,增加成骨细胞的分化能力是防治骨质疏松症的主要策略[13]。前期相关研究[14]发现,葛根素能够促进成骨细胞的分化。但是,葛根素对成骨细胞分化的影响以及自噬在成骨细胞分化过程中的作用目前还不清楚。
自噬参与骨组织的代谢过程,适当水平的自噬使成骨细胞能够存活于低氧、营养缺乏甚至高渗的环境[15]。此外,自噬在维持骨稳态方面起着至关重要的作用,与骨质疏松症的发生密切相关[11]。研究[16-17]表明,自噬相关蛋白参与成骨细胞分化、矿化和骨形成。敲除FIP200(参与组成fip200-atg13-ulk1复合体)的基因鼠模型中,由于骨形成严重减少导致骨质疏松症的发生,导致成骨细胞多种自噬缺陷,包括p62的过表达、LC3-Ⅱ 的转换不充分、自噬流的缺乏等,影响骨的形成[18-19]。敲除自噬相关基因7可导致大鼠松质骨的骨量和皮质骨的厚度下降,皮质骨孔隙度增加,诱导骨质疏松症的发生[20]。
本研究结果表明,成骨细胞在分化过程中表现出一定的基础自噬水平,葛根素在促进成骨细胞分化的同时,也明显增强细胞的自噬能力,表现为细胞内自噬小体的数量增加和自噬特异性蛋白LC3-Ⅱ和BECLIN 1的表达增强。同时还发现,加入自噬抑制剂3-MA后,葛根素的促成骨细胞分化作用受到明显抑制,说明自噬在葛根素促进成骨细胞分化的过程中扮演着重要的角色,葛根素可能通过自噬调节成骨细胞分化。
本研究深入分析了自噬在葛根素促成骨细胞分化中的分子生物学作用,明确了葛根素通过自噬调节成骨细胞分化潜在的机制,为临床应用中药单体防治骨质疏松症提供理论基础及实验依据。
