涉及簇毛麦6V染色体结构变异体的筛选与鉴定
2021-09-26张伟
摘 要:簇毛麦是小麦的一个野生近缘种,抗病性、抗逆性强,蛋白质和赖氨酸含量高,是小麦遗传改良的重要基因源。为定位、转移和利用簇毛麦有益基因,南京农业大学细胞遗传所通过花粉辐射,获得了一批涉及簇毛麦不同染色体且不同区段的结构变异体。该研究从这些结构变异体中筛选鉴定涉及簇毛麦6V染色体的结构变异体,共筛选鉴定出13种涉及簇毛麦6V染色体的结构变异体,包括2个整臂易位、5个小片段易位、4个大片段易位、1个缺失和1个中间插入易位,这些结构变异体的筛选鉴定将为进一步定位和利用簇毛麦6V染色体上的优异基因奠定基础。
关键词:小麦;簇毛麦;染色体结构变异体;基因组原位杂交;6V特异分子标记
中图分类号 S512.1 文献标识码 A 文章编号 1007-7731(2021)17-0021-06
Screening and Identification of Chromosomal Aberrations Involving Haynaldia villosa Chromosome 6V
ZHANG Wei
(Tongling Agricultural Technology Management Service Center, Tongling 244000, China)
Abstract: Haynaldia villosa (syn. Dasypyrum villosum) is a wild relative species of wheat (Triticum aestivum). It has strong ability of diseases and stress resistance, high protein and lysine contents. These characters make it an important gene pool for wheat genetic improvement. To localize, transfer and use interested genes of H. villosa, a series of involving different H. villosa chromosome and chromosomal region lines induced by irradiation have been developed at the Cytogenetics Institute, Nanjing Agricultural University, China. The aim of this study is to screen and identify structural changes involving H. villosa chromosome 6V in these chromosomal aberrations. Finally, 13 types of chromosomal aberrations involving H. villosa chromosome 6V were identified, including 2 of whole arm translocations, 5 of small alien segment translocation, 4 of large alien segment translocation, 1 of chromosomal deletion and 1of intercalary translocation. Screening and identification of these chromosomal aberrations will be basis to localize and use interested genes of H. villosa chromosome 6V.
Key words: Wheat; Haynaldia villosa; Chromosomal aberrations; Genomic in situ hybridization; Specific molecular-markers of 6V
簇毛麥(Haynaldia villosa,又名Dasypyrum villosum,2n=14,VV)属小麦族簇毛麦属,是小麦的一个野生近缘种,它不但抗小麦白粉病、锈病、全蚀病、眼斑病等多种病害[1-3],而且具有分蘖力强、抗寒耐旱、密穗多花、籽粒蛋白质含量高等特点[4,5],是小麦遗传改良的重要基因源。目前,簇毛麦的一些基因的表型性状已初步定位,如抗白粉病[6]、眼斑病[2,7]和瘿螨[3]等基因已转移到普通小麦背景中,分别被定位在6VS、4VL和6V染色体上。但是,与簇毛麦巨大的优异基因源相比,还有许多可供小麦品种改良的性状尚未利用,甚至还未定位到具体染色体或染色体片段。因此,进一步创造小麦-簇毛麦染色体易位系、缺失系等材料,对簇毛麦有益基因的定位及有目标地向普通小麦转移显得十分必要。基于此,南京农业大学细胞遗传所以硬粒小麦-簇毛麦双倍体为基础材料,利用花粉辐射与有性杂交相结合,创制出大量的簇毛麦染色体结构变异体[8,9],并对这些材料用中国春连续回交,已得到普通小麦背景中只包含一种形式的簇毛麦染色体结构变异体的单株。准确鉴定这些单株所涉及的簇毛麦染色体身份,对于簇毛麦有益基因的定位及有目标地创造遗传种质,进而应用于育种研究,具有重要的现实意义和应用价值。
簇毛麦6V染色体除携有抗白粉病基因Pm21外,还有抗秆锈病[10]及瘿螨等有益基因。本研究旨在利用簇毛麦6V的特异分子标记筛选鉴定涉及6V的染色体结构变异体,从而为定位和利用这些优异基因提供遗传中间材料。
1 材料与方法
1.1 试验材料 普通小麦中国春(Triticum aestivum cv. Chinese Spring,CS)、簇毛麦(H. villosa,由南京植物园从英国剑桥植物园引进)、硬粒小麦(T. durum)(由中国农业科学院引种组从国际玉米小麦改良中心引进,编号“中引1286”)、硬粒小麦-簇毛麦双倍体(T. durum-H. villosa amphiploid,由南京农业大学细胞遗传研究所选育)、小麦-簇毛麦二体异附加系DA6V和硬粒小麦-簇毛麦双倍体花粉辐射的回交自交后代,均由南京农业大学细胞遗传研究所保存提供。
1.2 试验方法
1.2.1 根尖细胞染色体制片 种子在垫有湿滤纸的培养皿中置25℃培养箱中萌发,待根长1~2cm时剪取根尖,在冰水中预处理22~24h后,用卡诺氏固定液(乙醇∶冰醋酸=3∶1)固定,2~7d后在45%醋酸中压片,-20℃冷冻揭片。
1.2.2 荧光原位杂交 荧光原位杂交程序参照Zhang等[11]的方法,探针包括簇毛麦基因组和来自黑麦的重复序列pSc119.2[12,13]。杂交制片洗涤染色后,在Olympus BX60型荧光显微镜下观察,用Olympus DP72数码显微成像系统摄取图像。
1.2.3 引物 选取6V上共12个特异分子标记,引物信息见表1。以上引物均由上海英骏生物公司合成。
1.2.4 PCR反应 PCR体积为10mL,包含约20~30ng模板DNA,1×buffer,1.5mmol L-1 MgCl2,200mmol L-1 dNTP,左右引物终浓度各为0.2mmol L-1,0.5U Taq DNA聚合酶(以上试剂均来自Promega公司)。PCR程序为94℃变性3min;94℃ 30s、55~60℃(依引物而定)50s、72℃ 1min,33个循环;72℃延伸10min。
1.2.5 产物检测 参照Tixier等[18]的方法检测产物,其中所用Marker为DL2000(Promega)。PCR扩增产物或酶切产物中加入聚丙烯酰胺凝胶电泳专用的Loading buffer 2mL,混匀,取3mL样液用8%聚丙烯酰胺凝胶(39∶1)电泳检测结果。电泳时总电压为180V,电泳1.5h左右,银染后将胶放在凝胶成像仪上观察照相。
1.3 簇毛麦染色体结构变异体的分类 在本研究中,以簇毛麦为对象将染色体结构变异分为易位(translocation)和缺失(deletion)2种类型。易位又分为外源大片段易位(large alien segment translocation,LAST,即W-V·V)、外源小片段易位(small alien segment translocation,SAST,即W·W-V)、整臂易位(whole arm translocation,WAT)以及簇毛麦型插入易位(Ti-V,即W-V-W)。
2 结果与分析
2.1 基因组原位杂交检测 南京农业大学细胞遗传所将60Co-γ射线照射的硬粒小麦-簇毛麦双二倍体花粉,授给已去雄的普通小麦品种中国春,在M1代即用基因组原位杂交检测到了大量的涉及簇毛麦染色体不同区段的结构变异(易位或缺失)。但由于选用硬粒小麦-簇毛麦双二倍体作为辐射亲本,涉及整套簇毛麦染色体组,因此在早期世代,多种类型的结构变异往往一起出现或伴有完整的簇毛麦染色体出现(图1a,b)。
这些含有多种类型的簇毛麦染色体结构变异体或伴有多条完整簇毛麦染色体的单株,给分子标记分析带来不便。对这些单株连续用普通小麦中国春回交2~3次,加上GISH的定向选择,可以筛选得到在小麦背景中只存在有1种形式的簇毛麦染色体结构变异材料(图2a,b)。
目前,经GISH检测共得到各类簇毛麦染色体结构变异体材料140份,其中包括:小片段易位(SAST)41份、大片段易位(LAST)32份、整臂易位(WAT)44份、簇毛麦染色体缺失(Del)18份和簇毛麦型插入易位(Ti-V)5份(图3,其中整臂易位只给出部分示例)。
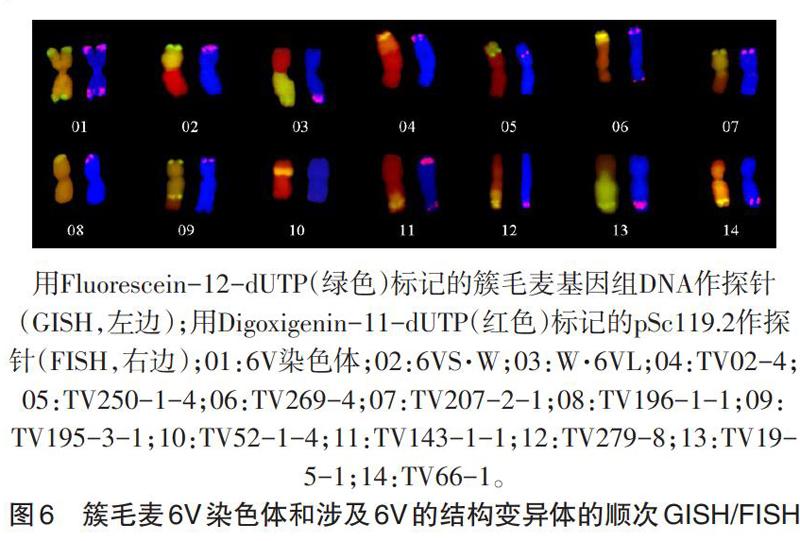
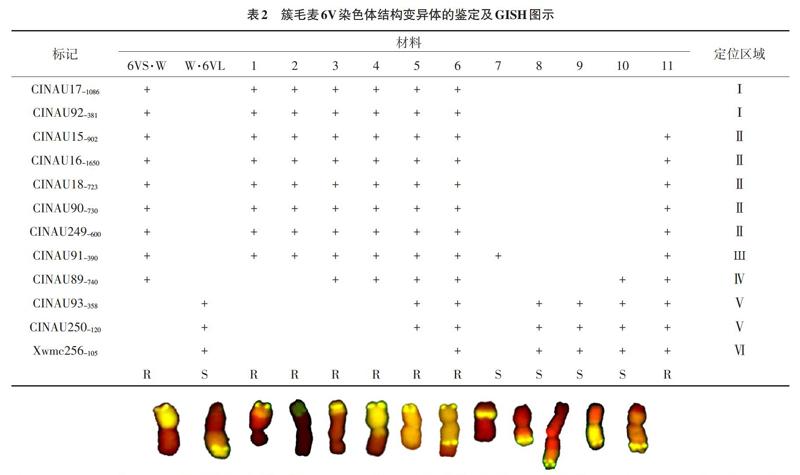
2.2 分子标记鉴定 仅通过基因组原位杂交无法确定这些结构变异体中所涉及的簇毛麦染色体身份,为此我们利用6V上共12个特异分子标记对这些结构变异体进行了分析鉴定,图4a-b是6V长、短臂标记CINAU93-358、CINAU91-390的扩增结果,从图中可以看出共有13份结构变异体材料可扩增出相应的特异条带,对照它们的GISH图发现这13份结构变异体材料分别包括:2个整臂易位(6VS·W和W·6VL)、5个小片段易位(TV02-4、TV250-1-4、TV269-4、TV143-1-1和TV279-8)、4个大片段易位(TV207-2-1、TV195-3-1、TV19-5-1和TV66-1)、1个缺失(TV196-1-1)和1个中间插入易位(TV52-1-4)。总的扩增结果见表2。
注:“+”表示对应列中的标记可以在相应行中扩增出特异带;“空白”表示未能扩增出特异带;“Ι-Ⅵ”表示标记在染色体上的定位区域;“R”表示抗白粉病,“S”表示感白粉病。1: TV02-4; 2: TV250-1-4; 3: TV269-4; 4: TV207-2-1; 5: TV196-1-1; 6: TV195-3-1; 7: TV52-1-4; 8: TV143-1-1; 9: TV279-8; 10: TV19-5-1; 11: TV66-1。
标记结果(表2)表明:单株TV02-4和TV250-1-4的扩增结果相同,都涉及6VS部分区段的小片段易位,但尚不能区分两者之间易位片段的大小;单株TV269-4也是涉及6VS部分区段的小片段易位,但其易位的片段较TV02-4和TV250-1-4的大;单株TV207-2-1是涉及6VS整臂及6VL部分区段的大片段易位,所涉及的6VL区域不含任何6VL的标记;单株TV196-1-1是涉及6VL末端的缺失染色体;单株TV195-3-1能扩增出6V所有的标记,易位的片段最大;TV52-1-4是一個中间插入易位,它仅能扩增出CINAU91-390这个标记,插入的是6VS片段;单株TV143-1-1和TV279-8的扩增结果相同,都是涉及6VL的小片段易位,但无法区分两者之间易位片段的大小;单株TV19-5-1能扩增出长臂的3个标记,但只扩增出短臂的近着丝粒区域的1个标记,而TV66-1除扩增出长臂所有标记外,还能扩增出位于短臂不同区域的7个标记,所涉及的6VS区域较TV19-5-1的大。
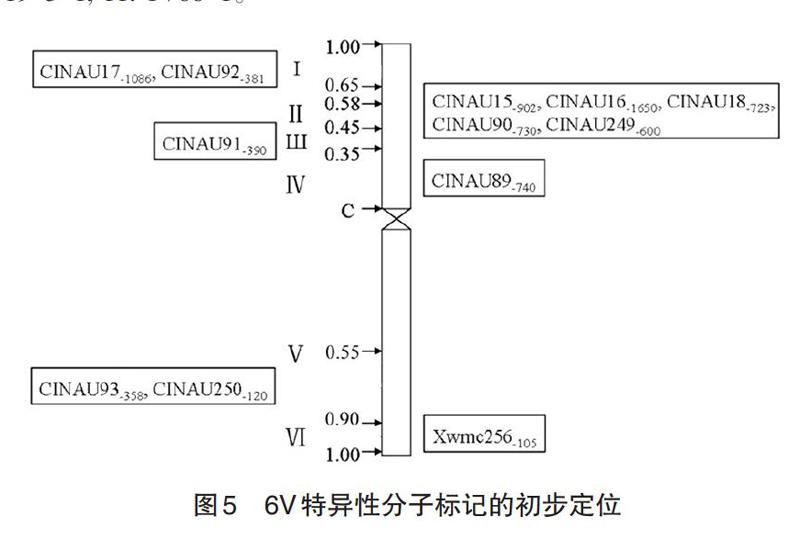
根據表2中标记的扩增结果和引物设计所用EST序列的定位信息,对6V染色体的标记进行了初步的物理定位(图5),短臂9个标记被分为4个区域:Ι区2个标记定位于FL:0.65~1.00,Ⅱ区5个标记定位于FL:0.45~0.58,Ш区1个标记定位于FL:0.35~0.45,Ⅳ区1个标记定位于FL:0.00~0.35;长臂3个标记被分为2个区域:Ⅴ区2个标记定位于FL:0.55~0.90;Ⅵ区1个标记定位于FL:0.90~1.00。
2.3 荧光原位杂交分析 来自黑麦的重复序列pSc119.2可以在簇毛麦6V染色体的末端产生杂交信号(张伟,私人通讯),利用这一特点对涉及6V的13份结构变异体进行了分析。对同一张染色体制片,首先进行基因组原位杂交,将信号洗掉后再利用Digoxigenin-11-dUTP标记的pSc119.2作为探针进行荧光原位杂交(图6)。从图6可以看出,pSc119.2在6V染色体的2个末端都有红色的杂交信号,TV52-1-4是一个中间插入易位,没有pSc119.2的杂交信号,表明是插入了6VS染色体的中部区域;其他12份材料都有红色的pSc119.2的杂交信号,表明这些结构变异体都保留了6V的1个原始末端。材料TV250-1-4的pSc119.2的信号出现在了易位染色体的近末端,可能是由于辐射引起的断裂-重接的原因,6VS的末端连接到了小麦染色体上。
3 讨论
南京农业大学细胞遗传所一直致力于簇毛麦有益基因的利用和转移研究,培育出的含有抗白粉病基因Pm21的6VS·6AL易位系已在小麦育种中得到了广泛应用,并育成了南农9918[19]、石麦14、内麦9号等高抗白粉病的小麦品种。但相对于簇毛麦庞大的优良基因资源来说,目前应用到实际生产中的并不多,一些基因甚至还未定位到具体的染色体或染色体片段上。这一方面是因为没有合适的群体对这些基因进行标记定位;另一方面是由于涉及相关染色体不同断裂位点的结构变异体的数量偏少。因此,开发高效诱导和高效筛选结构变异体的方法更具现实意义。Bie等[8]和Cao等[9]将60Co-γ射线照射的硬粒小麦-簇毛麦双倍体花粉,授给已去雄的普通小麦品种中国春,在M1代即用基因组原位杂交检测到了大量涉及簇毛麦染色体不同区段的结构变异(易位或缺失),在对这些结构变异体的传递率进行统计时发现,多数结构变异体能在后代中稳定地传递。对这些结构变异体单株,用中国春连续回交,经GISH检测已得到了140份在小麦背景中只存在一种形式的簇毛麦染色体结构变异体。用簇毛麦各条染色体的特异分子标记对这些结构变异体分析,可以快速地确定其涉及的簇毛麦染色体身份。本研究利用簇毛麦6V染色体的标记,共筛选鉴定出13份涉及6V的结构变异体。另外,利用pSc119.2在簇毛麦6V染色体末端产生的杂交信号来判断辐射造成的断裂-重接过程中,其末端是否发生了变化,本研究中材料TV250-1-4就是末端连接到小麦染色体上形成的易位染色体。
簇毛麦6V染色体除含抗白粉病基因Pm21外,还有抗秆锈病及瘿螨等基因,Qi等[10]利用着丝粒断裂-融合在中国春-簇毛麦6V二体附加系与中国春6D单体杂交后代中选育出补偿性易位系T6AS·6VL,该易位系在温度16℃条件下对由小种Ug99(TTKSK)引起的小麦秆锈病有很好抗性,并将抗性基因定位于6V长臂上,命名为Sr52。本研究筛选鉴定的涉及6VL区域的结构变异体共8份,目前这些变异体的抗性鉴定正在进行之中,结合抗性结果可以进一步定位该基因。
目前,所筛选的13份结构变异体中多数仍处于杂合状态,遗传上仍不稳定,除了在自交过程中发生丢失外,还会出现着丝粒错分裂。在自交过程中选育到纯合的结构变异体可能需要较多世代,而利用花药培养并诱导加倍单倍体有可能加快选育的进程,这项工作本实验室正在开展之中。
作者简介:张伟(1980—),男,安徽铜陵人,博士,高级农艺师,从事农业技术推广工作。 收稿日期:2021-05-12
参考文献
[1]Qi L L,Chen P D,Liu D J,et al.The gene Pm21-a new source for resistance to wheat powdery mildew[J]. Acta Agron Sin,1995,21(3): 257-262 (in Chinese with English abstract).
[2]Murray T D,De La Pena R C,Yildrim A,et al. A new source of resistance to Pseudocercosporella herpotrichoides,cause of eyespot disease of aheat,located on chromosome 4V of Dasypyrum villosum[J]. Plant Breed,1994,113: 281-286.
[3]Chen Q,Conner R L,Laroche A. Molecular characterization of Haynaldia villosa chromatin in wheat lines carrying resistance to wheat curl mite colonization[J]. Theor Appl Genet,1996,93: 679-684.
[4]Blanco A,Resta P,Simeone R,et al. Chromosomal location of seed storage protein genes in the genome of Dasypyrum villosum (L.) Candargy[J]. Theor Appl Genet,1991,82: 358-362.
[5]De Pace C,Snidaro D,Ciaffi M,et al. Introgression of Dasypyrum villosum chromatin into common wheat improves grain protein quality[J]. Euphytica,2001,117: 67-75.
[6]Liu D J,Chen P D,Pei G Z,et al. Transfer of Haynaldia villosa chromosomes into Triticum aestivum. In: Miller T E,Koebner R M D. (eds). Proc 7th Int Wheat Genet Symp,Cambridge,U.K. 1988:355-361.
[7]Yildirim A,Jones S S,Murray T D. Mapping a gene conferring resistance to Pseudocercosporella herpotrichoides on chromosome 4V of Dasypyrum villosum in a wheat background[J]. Genome,1998,41: 1-6.
[8]Bie T D,Cao Y P,Chen P D. Mass production of intergeneric chromosomal translocatons through pollen irradiation of Triticum durum-Haynaldia villosa amphiploid[J]. J Integr Plant Biol,2007,49: 1619-1626.
[9]Cao Y P,Bie T D,Wang X E,et al.Induction and transmission of wheat-Haynaldia villosa chromosomal translocations[J]. J Genet Genomics,2009,36: 313-320.
[10]Qi L L,Pumphrey M O,Friebe B,et al. A novel Robertsonian translocation event leads to transfer of a stem rust resistance gene (Sr52) effective against race Ug99 from Dasypyrum villosum into bread wheat[J]. Theor Appl Genet,2011,123: 159-167.
[11]Zhang W,Gao A L,Zhou B,et al. Screening and applying wheat microsatellite markers to trace individual Haynaldia villosa chromosomes[J]. Acta Genetica Sinica,2006,33(3): 236-243.
[12]Bedbrook JR,Jones J,?Neill M,et al. A molecular description of telomeric heterochromatin in Secale species[J]. Cell,1980,19: 545-560.
[13]McIntyre CL,Pereira S,Moran LB,et al. New Secale cereale (rye) DNA derivatives for the detection of rye chromosome segments in wheat[J]. Genome,1990,33: 635-640.
[14]Cao A Z,Wang X E,Chen Y P,et al. A sequence-specific PCR marker linked with Pm21 distinguishes chromosome 6AS,6BS,6DS of Triticum aestivum and 6VS of Haynaldia villosa[J]. Plant Breed,2006,125:201-205.
[15]Chen Y P,Wang H Z,Cao A Z,et al. Cloning of a resistance gene analog from wheat and development of a co-domninant PCR marker for Pm21[J]. J Integr plant Biol,2006,48(6):715-721.
[16]王春梅,別同德,陈全战,等.簇毛麦6V染色体短臂特异分子标记的开发和应用[J].作物学报,2007a,33(10):1595-1600.
[17]陈升位,陈佩度.簇毛麦6V染色体短臂特异性EST标记的开发及缺失定位[J].麦类作物学报,2010,30(5): 789-795.
[18]Tixier M H,Sourdille P M S. Detection of wheat microsatellite using no-radioactive silvermitrate staining method[J].Genet Breed,1997,51: 175-177.
[19]Chen PD,Zhang SZ,Wang XE,et al. New wheat variety Nannong 9918 with high yield and powdery mildew resistance[J].J Nanjing Agric Univ,2002,25(4):105-106(in Chinese with English abstract).
(责编:张宏民)
