肉苁蓉寄生对梭梭光合特性的影响
2021-09-24于霞霞郭晔红贾存勤马兴东马旭东
于霞霞, 郭晔红*, 杜 弢, 贾存勤, 李 欠, 马兴东, 马旭东
(1.甘肃农业大学农学院中药材研究所,甘肃省中药材规范化生产技术创新重点实验室,甘肃省药用植物栽培育种工程研究中心,甘肃省干旱生境作物学重点实验室,甘肃 兰州 730070;2.甘肃中医药大学药学院,甘肃 兰州 730000;3.甘肃汇勤生物科技有限公司,甘肃 白银 730600)
肉苁蓉CistanchedeserticolaMa是我国名贵中药材“大芸”的基原植物,寄生在梭梭的根部,其生长所需的营养物质全部由梭梭供给;梭梭Haloxylonammodendron(C. A. Mey.) Bunge属藜科梭梭属多年生灌木或小乔木,为国家濒危三级保护植物[1],具有耐旱、耐寒、抗盐碱的特性,是西北地区重要的防风固沙先锋树种。全世界约有梭梭10种,是古地中海区系的遗留种,分布于地中海至中亚地区,构成亚洲荒漠区分布最广泛的荒漠植被类型[2],中国有梭梭和白梭梭HaloxylonpersicumBunge ex Boiss. et Buhsee 2种,主要分布在内蒙古、新疆、甘肃、青海、宁夏等地,是西北荒漠半荒漠区珍贵的植物资源[3]。由于自然作用和人为因素对荒漠植被的严重破坏,使天然梭梭林大面积减少,人工梭梭林已成为当今梭梭群落存在的主要形式。
光合作用是植物的重要生理生态学特性之一[4], 它是植物生长发育的基础,也是植物长势优劣的决定性因素,植物对环境条件变化具有十分敏感的生理响应[5]。梭梭作为肉苁蓉的能量供应源,其制造有机物的光合作用至关重要,近年来有关梭梭的研究,主要集中在梭梭群落生长、林分结构、生物学特性等方面[6-10],而关于梭梭光合生理方面的研究则少有报道。鞠强等[11]研究了梭梭光合生理生态过程与干旱环境的相互关系,有关寄生肉苁蓉后对其寄主植物光合生理的影响,郭东锋[12]、杨太新等[13]研究了管花肉苁蓉寄生对寄主柽柳光合特性的影响,而关于荒漠肉苁蓉寄生对寄主梭梭光合作用特性的影响以及环境因子与光合特性之间的相互关系则罕见报道。本实验研究了寄生荒漠肉苁蓉对梭梭光合特性的影响以及环境因子对梭梭光合生理指标影响的程度,旨在探明梭梭光合生理指标对肉苁蓉寄生的响应机制及其对环境的适应程度,为荒漠化综合治理和肉苁蓉产业持续发展提供科学依据。
1 材料与方法
1.1 试验地概况 试验点设在甘肃靖远糜滩乡,东经104°68′,北纬36°59′,海拔1 450 m,属温带干旱半干旱气候,年均气温9.2 ℃左右,年极端最高气温35.1 ℃,年均降水量240 mm,年蒸发量1 634 mm,平均日照时数2 696 h,无霜期120 d。
1.2 材料 供试植物为甘肃靖远县长势良好的4年生人工栽培梭梭,小区和管理水平与大田栽培相同。采用梅花点取样法选取寄生和未寄生肉苁蓉的梭梭植株各5株,每株梭梭分别在上、中、下(依次距地面75、50、25 cm处)部选取生长一致的各功能叶片进行测定,并每次在梭梭上、中、下测定光合的叶片处采集叶片测定叶绿素,每株重复3次。将所取样品立即置于液氮罐中贮藏,带回实验室于-80 ℃超低温冰箱中冷藏备用。
1.3 方法 试验于2018年5月4~6日(苁蓉出土期)、5月27~29日(开花结实期)、6月19~21日(枯萎期)3个时间段进行,以开始测定日期作为测定日期,每次连测3 d,求得3 d的平均值作为该测定期的值,求得3 d中每个时刻的平均值绘制光合指标日变化曲线,所选日期晴朗无风,用LI-6400型光合仪对梭梭叶片进行活体测定,每天测定时间为8:00~18:00,每隔2 h测定1次,中午加测1次,每个处理选取5株,每株重复3次,取其平均值。测定项目包括梭梭的净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)、胞间CO2浓度(Ci)、水分利用率(WUE=Pn /Tr) 等各项光合生理指标,以及空气温度(Ta)、空气相对湿度 (RH)、叶片温度(Tl)、大气压亏缺(Vpd)、光合有效辐射(PAR)、大气CO2浓度(CO2)等环境因子。将Pn、Tr、Gs、Ci、WUE作为物种数据源;将Tl、RH、Vpd、Ta、PAR、CO2作为环境因子数据源,分析其光合生理指标与环境因子间的相关性。在每个时期进行测定光合作用的3 d中,连续3 d进行取样,测定叶绿素的含量,求得平均值作为该时期的测定值,叶绿素含量的测定采用丙酮法[14]。
1.4 统计分析 采用SPSS 19.0对寄生和未寄生肉苁蓉的梭梭叶片的光合生理指标采用One-way ANOVA中的LSD和Duncan法分析,Excel及CanoDraw法绘图,采用Canoco 4.5 软件先进行DCA分析后选择RDA分析并排序,RDA排序图中的环境因子(Tl、RH、Vpd、Ta、PAR、CO2)用带箭头的虚线表示,光合生理指标(Pn、Tr、Gs、Ci、WUE)用带箭头的实线表示。
2 结果与分析
2.1 肉苁蓉的寄生对梭梭光合生理指标的影响 3个测量时期中,每个时期寄生和未寄生肉苁蓉梭梭的净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)随时间的变化均有一致的变化规律,均呈双峰型曲线,表现出明显的光合午休现象。Pn从8:00开始逐渐升高,12:00出现第1个峰值,随后大幅度降低,在14:00降至最低,之后有所回升,在16:00达到第2次峰值,随后又逐渐下降,且每个时期寄生和未寄生肉苁蓉的梭梭第1个峰值均大于第2个峰值。除5月4日未寄生肉苁蓉梭梭的Gs、5月27日寄生和未寄生肉苁蓉梭梭的Tr,其他时间寄生和未寄生肉苁蓉梭梭的Gs、Tr与Pn的峰值时间一致,但2个峰值的大小则无明显的规律,见图1。每个时期梭梭Pn日均值都表现为未寄生肉苁蓉的梭梭>寄生肉苁蓉的梭梭,其中5月4日和5月27日未寄生肉苁蓉梭梭的Pn日均值高于寄生肉苁蓉的梭梭(P<0.05);各个时期梭梭的Gs和Tr日均值都表现为未寄生肉苁蓉的梭梭>寄生肉苁蓉的梭梭,但未达到显著性水平,见表1。
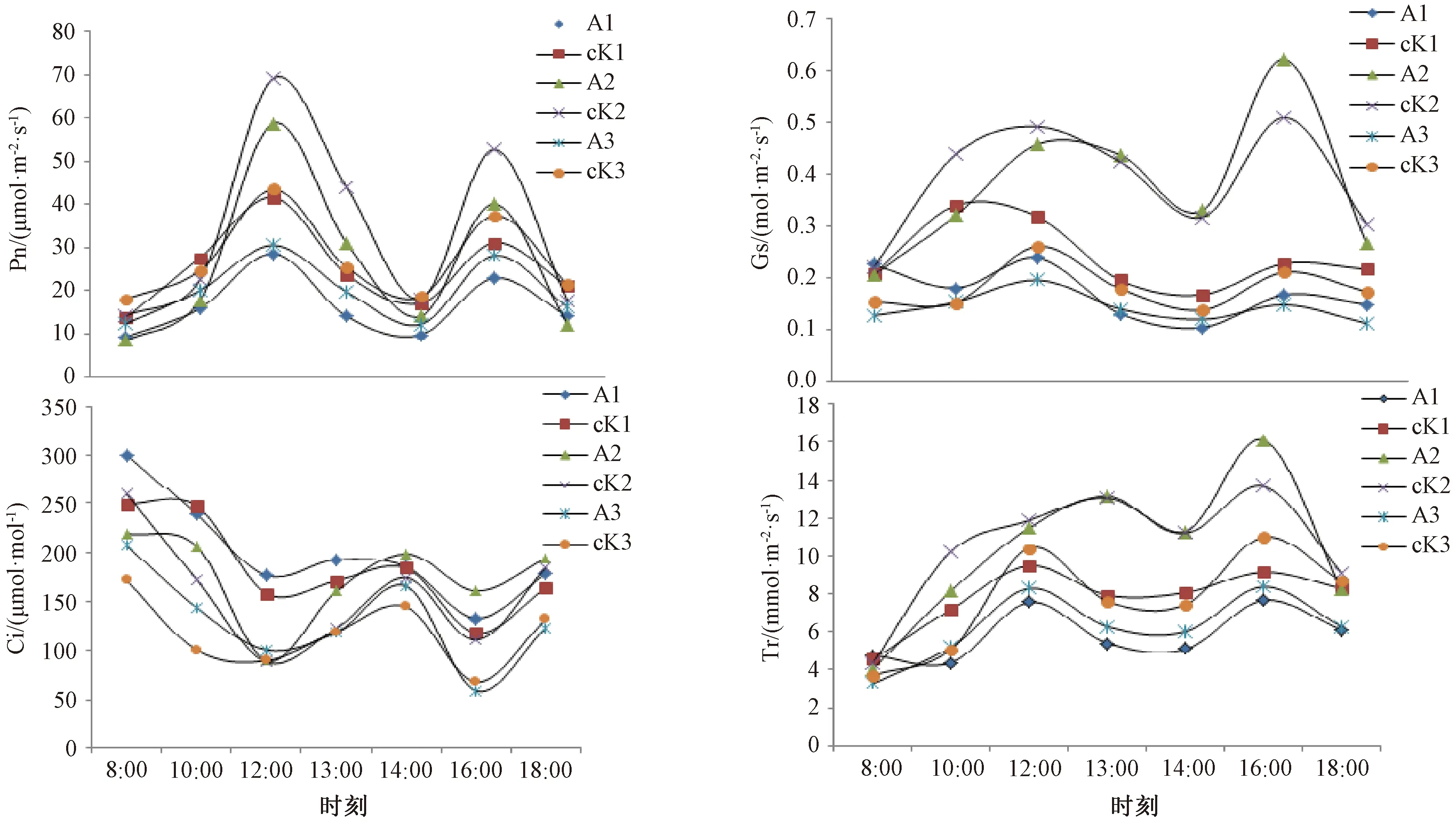
注:1、2、3分别代表5月4日、5月27日、6月19日。A为寄生了肉苁蓉的梭梭,cK为未寄生肉苁蓉的梭梭。图1 肉苁蓉不同寄生时期梭梭光合生理特性日变化
3个测量时期中,每个时期寄生和未寄生肉苁蓉梭梭的Ci随时间的变化趋势一致,且与各自Pn、Gs、Tr变化趋势相反,呈倒双峰型曲线,在8:00、14:00、18:00 Ci值较高,Ci出现峰值的时间与Gs、Tr、Pn出现谷值的时间一致。除5月4日外,其他2个时期梭梭Ci日均值与Gs、Tr、Pn日均值相反,为寄生肉苁蓉的梭梭>未寄生肉苁蓉的梭梭,5月4日未寄生肉苁蓉的梭梭Ci仅高于寄生肉苁蓉梭梭 1.20 μmol/mol,见图1C、表1。
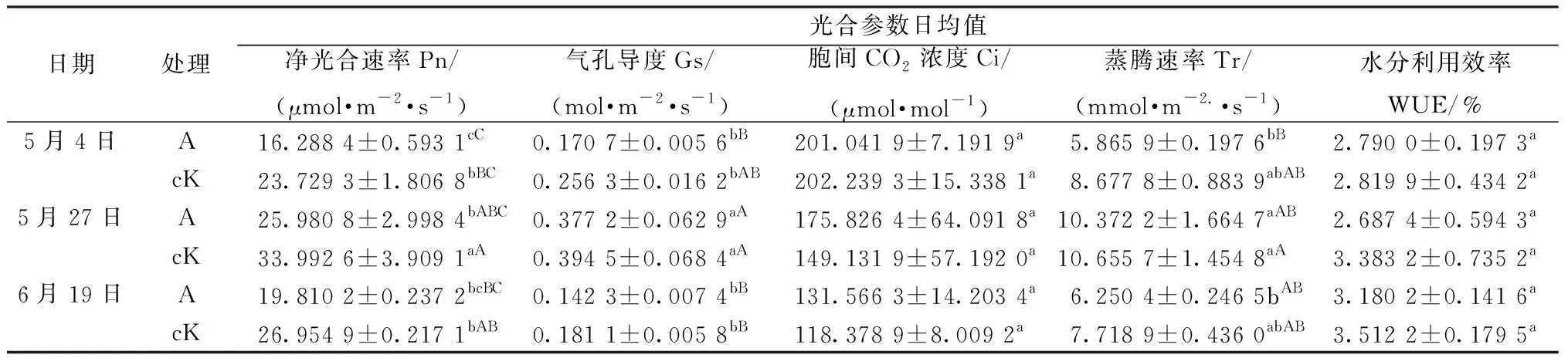
表1 肉苁蓉寄生对梭梭光合参数日均值的影响
2.2 肉苁蓉的寄生对梭梭水分利用效率及叶绿素含量的影响 水分利用效率是反映植物水分利用特性的重要参数,荒漠植物具有较高的水分利用效率,尤其是在水分亏缺时能以高水分利用效率来抵御干旱胁迫[15],逆境胁迫会影响植物光合色素的合成[16]。梭梭WUE测量的3个时期中,各自的寄生肉苁蓉梭梭和未寄生肉苁蓉梭梭日变化趋势一致,不同生长期,WUE日变化趋势有差异。WUE日变化趋势在5月4日和5月27日大体相同,均为双峰型曲线,峰值时间与Pn的一致,12:00达到一天中的最大值,14:00为一天中的最小值。与其不同,6月19日梭梭WUE日变化曲线呈降-升-降的趋势,在8:00~14:00逐渐降低,14:00后WUE有所回升,到16:00达到一个峰值,随后又下降。最大值出现在8:00,最小值出现的时间与其他2个时期一致,为14:00。同一测量时期梭梭WUE日均值表现为未寄生肉苁蓉梭梭>寄生肉苁蓉梭梭,但未达到显著性水平,表明肉苁蓉的寄生降低了梭梭的WUE,但不改变日变化趋势,见图2、表1。3个测量时期,梭梭叶绿素a和叶绿素b含量在每个测量期均为未寄生肉苁蓉梭梭>寄生肉苁蓉梭梭,但未达到显著性水平。6月19日,总叶绿素含量寄生肉苁蓉梭梭(0.10)略大于未寄生肉苁蓉梭梭(0.09),叶绿素a/b在5月4日为寄生肉苁蓉梭梭>未寄生肉苁蓉梭梭,其他2个时期,总叶绿素与叶绿素a/b均表现为未寄生肉苁蓉梭梭>寄生肉苁蓉梭梭,但未达到显著性水平,见表2。由于梭梭的叶绿素含量较低,且在不同时期寄生和未寄生肉苁蓉梭梭的总叶绿素含量大小比较不一致,因此肉苁蓉的寄生是否降低了梭梭的总叶绿素含量还需做进一步的研究。
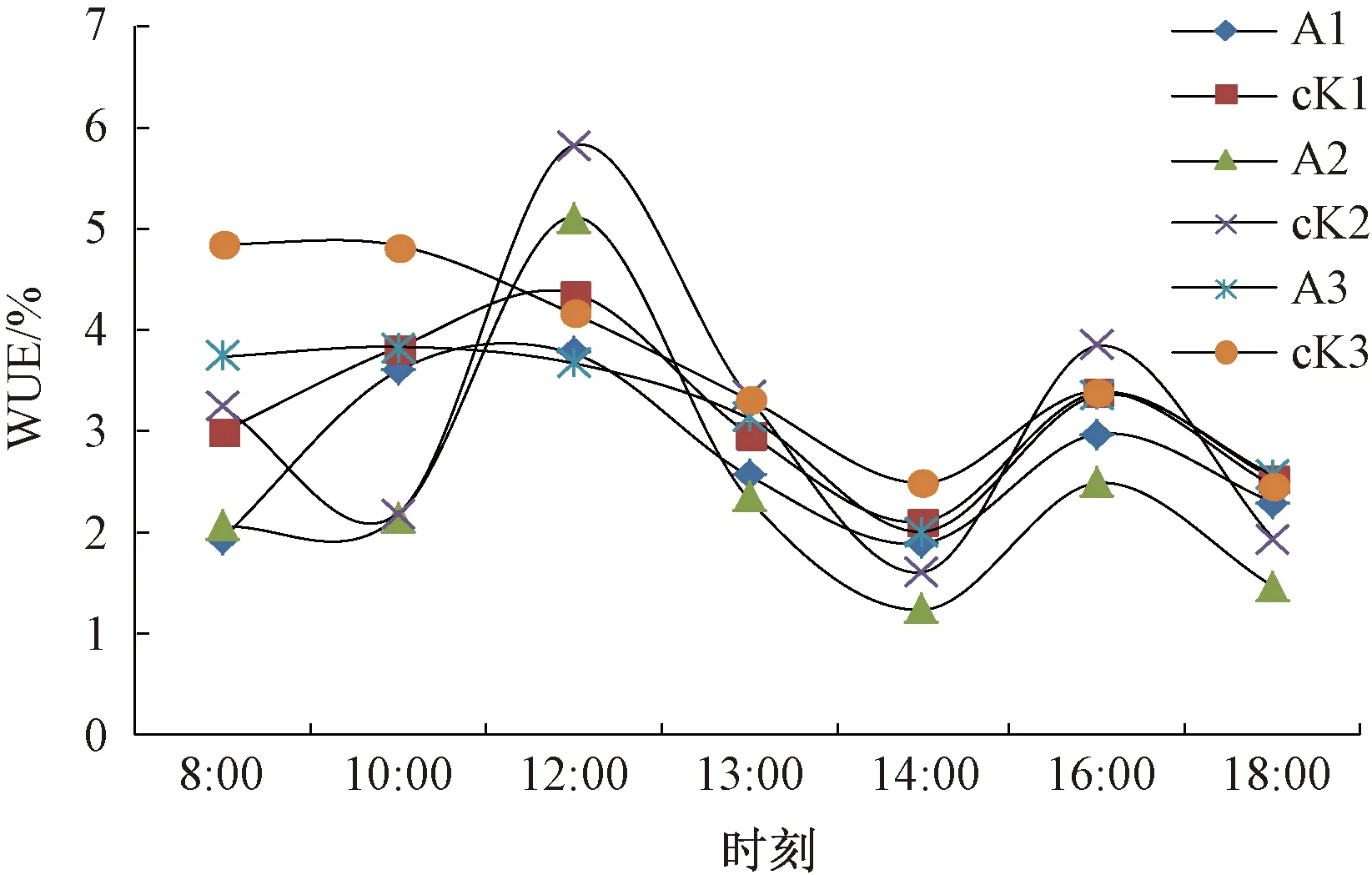
注:1、2、3分别代表5月4日、5月27日、6月19日。A为寄生了肉苁蓉的梭梭,cK为未寄生肉苁蓉的梭梭。图2 肉苁蓉不同寄生时期梭梭WUE日变化

表2 肉苁蓉寄生对梭梭叶绿素含量的影响
2.3 肉苁蓉寄生对梭梭光合生理指标与环境因子相关性的影响 梭梭作为强旱生盐生植物,对环境有着超强的适应能力[17]。本研究中第一排序轴对物种的解释量最大,故以第1排序轴为主分析环境因子与光合生理指标的相关性。5月4日和6月9日,影响寄生和未寄生肉苁蓉梭梭的主要环境因子排序一致,为CO2>Tl>Ta,见图3。对CO2与光合生理指标的相关性(相关系数)进行分析,5月4日,寄生肉苁蓉梭梭为0.444,未寄生肉苁蓉梭梭为0.698(P<0.01),6月19日,寄生肉苁蓉梭梭和未寄生肉苁蓉梭梭依次为0.906、0.927(P<0.01),见表3,表明肉苁蓉的寄生使梭梭主要光合生理指标与环境因子的相关性减弱。5月27日,寄生肉苁蓉梭梭和未寄生肉苁蓉梭梭的光合生理指标与环境因子的相关性相反,其中Tl对寄生和未寄生肉苁蓉梭梭的光合生理指标的影响作用最大,相关系数依次为0.415、-0.502(P<0.01),见表3,表明肉苁蓉的寄生使梭梭光合生理指标与环境因子的促进与抑制作用变得相反,且每个时期种—环境的相关性都表现为未寄生肉苁蓉的梭梭>寄生肉苁蓉的梭梭。
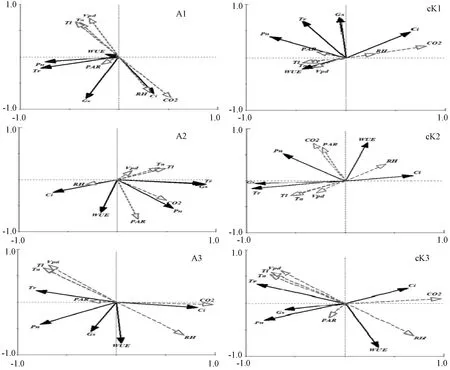
注:A、cK分别代表寄生和未寄生肉苁蓉的梭梭。1、2、3分别代表5月4日、5月27日、6月19日。图3 寄生与未寄生肉苁蓉梭梭的光合生理指标与环境因子的RDA二维排序图
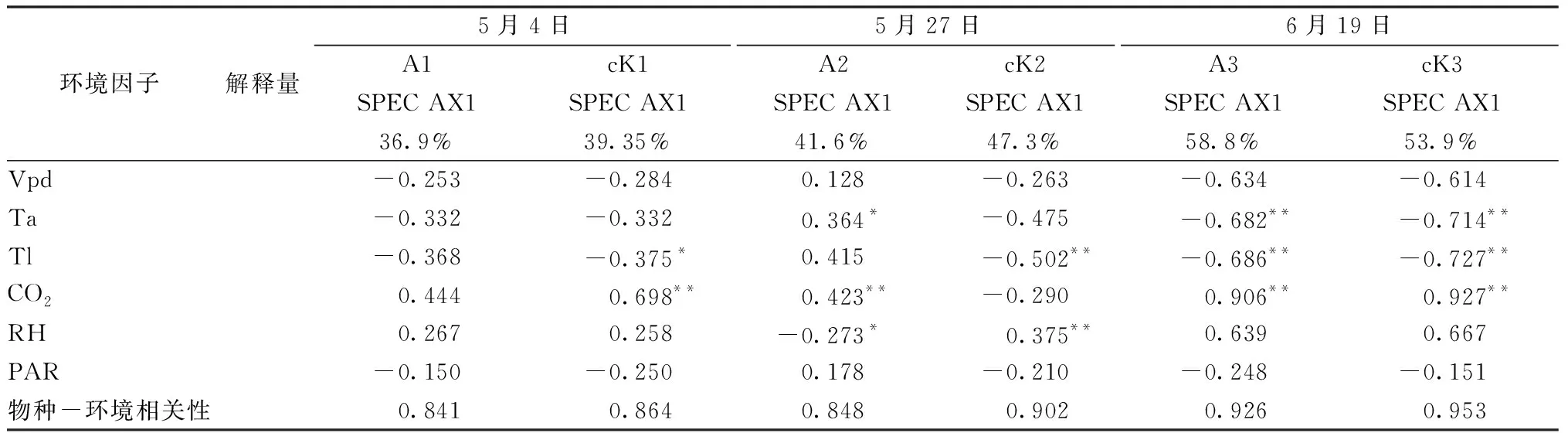
表3 第一排序轴与环境因子的冗余分析
2.4 梭梭光合指标与叶绿素含量的相关性分析 Pearson相关性分析(表4)表明,梭梭Pn与Ci、叶绿素a及叶绿素a/b呈负相关,但未达到显著性水平;与Tr呈显著正相关(r=0.859,P<0.05),其他指标中与Pn相关性较大的依次是总叶绿素、Gs、WUE,相关系数依次为0.764、0.759、0.517,表明蒸腾速率对净光合速率具有强烈的促进作用;梭梭Gs与Tr及总叶绿素、Tr与总叶绿素、总叶绿素与叶绿素a/b呈极显著正相关(P<0.01),相关系数依次为0.961、0.969、0.935、0.976,表明这些指标之间相互促进的程度最大。

表4 梭梭光合指标及叶色素含量的相关性分析
2.5 肉苁蓉寄生对梭梭光合特性影响的综合排序 对不同生长期所测梭梭光合及其相关指标用下式求各指标的具体隶属函数值。
(1)
或
(2)
式中,X为梭梭某一光合及其相关指标测定值,Xmax、Xmin分别为梭梭该指标测定值内的最大值和最小值。若所测指标与光合作用呈正相关,用式(1);若所测指标与光合作用呈负相关,用式(2)。把每个时期每个处理梭梭光合及其相关指标的隶属值进行累加,并求其平均值,根据各处理平均值大小确定光和能力的强弱。
光合能力的强弱一般用净光合速率表示,本研究中通过对梭梭光合及其相关指标进行相关性分析,见表4,将与净光合速率呈正相关的指标用式(1)求隶属函数值,将与净光合速率呈负相关的指标用式(2)求隶属函数值,即Ci、叶绿素a、叶绿素a/b用式(2),其他指标用式(1)。
光合特性是多个光合生理指标综合作用的结果,根据寄生与未寄生肉苁蓉梭梭的各光合生理及其相关指标的隶属函数值求得平均隶属函数值并进行排序,可得不同生长期不同处理梭梭的光合能力的强弱顺序。表5显示,同一生长期,光合及其相关指标的平均隶属函数值均表现为未寄生肉苁蓉的梭梭>寄生肉苁蓉的梭梭,表明肉苁蓉的寄生使梭梭光合能力减弱。不同生长期的同一处理梭梭的平均隶属函数值的综合排序为5月27日>6月19日>5月4日,与Pn日均值的排序一致,进一步表明Pn是光合生理的代表性指标。
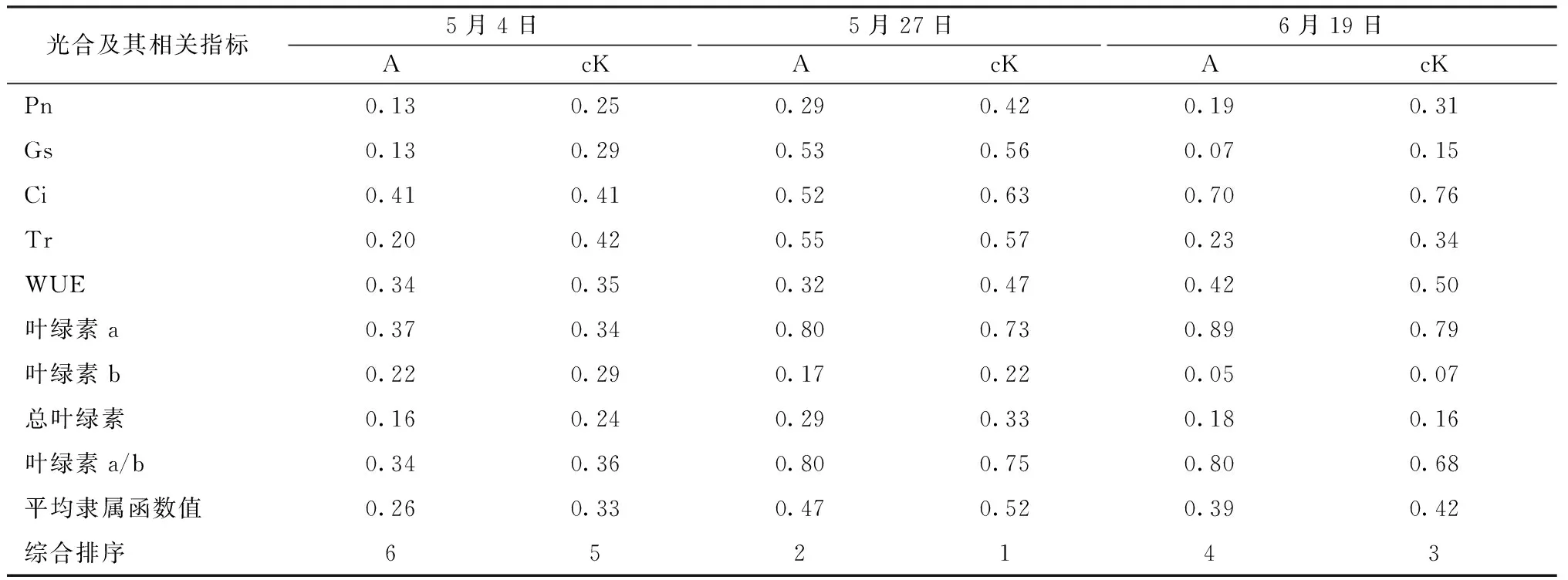
表5 肉苁蓉不同寄生时期梭梭光合作用综合排序
3 讨论
植物在受到胁迫后,会产生逆境生理[18-19],肉苁蓉的寄生给寄主梭梭造成了胁迫的环境,对寄主保护酶系统及渗透调节系统产生了明显的伤害,并降低了寄主梭梭的抗旱性[20]。肉苁蓉寄生对梭梭产生的逆境生理使类囊体膜系统及酶蛋白受损,导致叶绿素分解大于合成,表现为叶绿素(叶绿素a、叶绿素b)含量降低[21-22],本研究得出,肉苁蓉的寄生使寄主梭梭叶绿素a、叶绿素b含量降低。
植物在受到胁迫时,植物的光合作用场所类囊体薄膜受损和叶绿体基质代谢紊乱[23],光合作用减弱,表现为单位时间、单位叶面积吸收CO2的量或放出O2的量减小[24-25],当光合过程受阻,气孔导度降低,水分代谢失调,植物的蒸腾速率下降[26],因此,认为有效的形态调节和较强的气孔控制是其维持光合作用能力以及适应干旱的主要机制[27]。水分利用效率可以反映植物对干旱环境适应能力的强弱[28-29],植物在受到胁迫后,水分利用效率和生物产量降低,甚至出现由于含水量下降而萎蔫的现象[30]。本研究中,梭梭在受到肉苁蓉的寄生胁迫后,净光合速率,蒸腾速率、气孔导度、水分利用效率降低,胞间CO2升高,但肉苁蓉的寄生未改变梭梭光合特性的日变化趋势。相关性分析表明,蒸腾速率与净光合速率之间相互促进,总叶绿素含量最大程度的促进气孔导度、蒸腾速率和叶绿素a/b。
光合能力是多指标综合作用的结果,其光合指标之间具有相关性,因此需用光合生理及其相关指标综合评价植物的光合能力[31],环境中的光、温、水、气等生态因子变化可引起植物生理因子变化,外在生态环境条件和内在生理因子共同影响植物光合作用特性[32]。本实验结果与前人有关棉花、菠菜大豆和酸枣的研究一致[33-36],经冗余分析(RDA)寄生与未寄生肉苁蓉梭梭的光合指标与环境因子的相关性,表现为肉苁蓉的寄生使梭梭光合指标与环境因子的相关性减弱,寄生肉苁蓉的梭梭植株对环境因子表现出“迟钝型”响应。多指标综合评价寄生与未寄生肉苁蓉梭梭的光合能力强弱,其结果与单一指标评价的结果一致,表现为肉苁蓉的寄生使梭梭的光合能力减弱。
