瘦素及其受体在真孕和假孕母犬下丘脑-垂体-性腺轴各组织中的表达
2021-09-22狄和双王利刚韩大勇蒋珊珊孙庆蘅
狄和双,王利刚,韩大勇,蒋珊珊,孙庆蘅
(江苏农牧科技职业学院,江苏 泰州 225300)
近年来犬业经济蓬勃发展,但母犬配种后未受孕时假孕发生率达50%~70%,假孕易诱发母犬子宫疾患、不孕甚至死亡.犬假孕的原因主要是母犬发情排卵后未孕的情况下卵巢上黄体期的时间与真孕母犬黄体期时间相似甚至更久,导致犬配种后无论怀孕与否体内的孕酮水平相似,因此正常未孕母犬黄体期又被称为生理学或隐性假孕[1],母犬表现出与真孕母犬相似的怀孕征兆,增加了母犬早期妊娠诊断的难度,而B型超声波等诊断妊娠方法在犬场难以普及应用,因此,我们提出胚胎着床期可能是探寻早期鉴别犬真孕与假孕差异指标的关键时期.
研究已证实瘦素不仅与动物的体脂能量调节关系密切,瘦素及其受体对人及啮齿动物的青春期启动、性腺发育、胚胎着床及病理妊娠等繁殖活动发挥重要的作用[2-4],一定浓度的瘦素是小鼠正常妊娠时胚胎着床所必须的,但瘦素及其受体对于犬的胚胎着床的作用研究鲜见报道.我们前期的研究发现在母犬的胚胎着床期(配种后19~23 d)真孕母犬的外周血中瘦素水平极显著高于假孕母犬外周血清中的瘦素水平[5],推测瘦素可能参与了母犬的胚胎着床,瘦素可能是判别母犬真孕和假孕的激素指标.但真孕母犬外周血中高浓度瘦素水平的来源未知,为了对前期结论进行更深入的研究,本研究通过RT-PCR和免疫组织化学的方法观察胚胎着床期瘦素及其受体在真孕和假孕母犬下丘脑-垂体-性腺轴各组织中的表达差异,为验证瘦素是否参与了母犬胚胎着床以及瘦素是否可以作为判别母犬真假孕的关键因子探寻证据.
1 材料与方法
1.1 试验动物及处理
在江苏农牧科技职业学院实验犬舍选择饲养管理条件一致的中华田园犬,年龄(3~5岁)、体质量相近自然发情的健康母犬,选取用种公犬配种后20 d经B型超声波诊断妊娠的3只母犬,另外用输精管结扎的公犬进行自然配种后20 d的3只母犬作为假孕母犬模型(借鉴小鼠假孕模型的研究[6]),参照李成的方法[7],取各器官组织样品保存备用,本试验通过了我院动物实验伦理审核(另:试验犬采样后的尸体用做解剖标本的制作).
1.2 主要试剂
Trizol Reagent、cDNA第一链反转录试剂盒、PCR Mix购自HaiGene公司;兔抗犬瘦素多克隆抗体、兔抗犬瘦素受体的多克隆抗体及DAB显色试剂盒购自武汉博士德公司;SP免疫组化试剂盒购自北京博奥森生物公司;琼脂糖、氯仿、异丙醇、TAE缓冲液、无水乙醇等常规药品均购自国药集团;引物由Genscript公司合成.
1.3 RNA提取及RT-PCR
按常规方法提取各组织的RNA,溶解后-70 ℃保存.将检测完整的RNA反转录至cDNA,按以下组分配制反转录反应液:Total RNA 1 μg,5×RT MasterMix 4 μL,Oligo (dT)15 Primer 1 μL,Rnase Free H2O Up to 20 μL.在PCR仪上按以下条件进行反转录反应:30 ℃5 min, 55 ℃ 30 min,85 ℃ 5 min,4 ℃.针对犬Leptin基因(Genbank No.:NM_001003070)设计1对引物,犬Leptin-F:5′-ATACTCTAGGTGGTGGATCTGTTC-3′,犬Leptin-R:5′-TGTCTGCCGTTATGGTCTTAGG -3′,扩增片段为440 bp;针对犬leptin的受体基因(GenBank No.:AY823396)设计1对引物,犬 LeptinR-F:5′-GGTTTCACCAAAGATGCTATCG-3′,犬 LeptinR-R:5′-TGTGCCCAGGAACAATTCTTG-3′, 扩增片段为176 bp;针对犬甘油醛-3-磷酸脱氢酶(GAPDH)基因(Genbank No.:NM_001003142.1)设计1对引物, 犬GAPDH F:5′-GTGAAGGTCGGAGTCAACGG -3′,犬GAPDH R:5′-TCAGCAGAAGGAGCAGAGATG-3′ ,扩增片段为371 bp.以上引物均由Genscript公司合成.PCR反应总体积为50 μL,扩增循环条件为:95℃预变性5 min,95 ℃变性10 s,58 ℃退火10 s,72 ℃延伸30 s,共进行34个循环.取反应产物用1.5% TAE琼脂糖凝胶电泳,用凝胶成像系统(UVP,UVP -200)拍照并用软件分析条带灰度,分别以瘦素和瘦素受体基因对GAPDH的相对灰度比值作为瘦素及其受体的相对表达量.
1.4 瘦素及其受体的表达
取真孕和假孕母犬的各组织固定并制备石蜡切片,按狄和双的方法[8]进行免疫组织化学染色,一抗为兔抗犬瘦素多克隆抗体(1∶500)或兔抗犬瘦素受体的多克隆抗体(1∶500),37 ℃孵育1 h,阴性对照以磷酸缓冲盐溶液(PBS)代替一抗,二抗为羊抗兔IgG,37 ℃孵育20 min,辣根酶标记链酶卵白素37 ℃孵育20 min,DAB染色,苏木精轻度复染、脱水、透明,封片.显微镜下观察,细胞浆或细胞核内见棕黄色颗粒沉着为阳性细胞.根据Kraan等[9]的方法,测量平均光密度值.
1.5 数据统计及处理

2 结果与分析
2.1 瘦素及其受体的RT-PCR产物定性及半定量分析
真孕母犬与假孕母犬的下丘脑、垂体、卵巢和子宫组织中均有瘦素mRNA的表达,输卵管中未检测出瘦素mRNA的表达.真孕和假孕母犬的下丘脑、垂体、卵巢、子宫、输卵管组织中均检测到瘦素受体mRNA的表达,扩增的瘦素、瘦素受体及GAPDH的RT-PCR产物与理论长度相符合(图1).通过mRNA灰度分析并比较得出,真孕组母犬下丘脑、卵巢和子宫中瘦素的mRNA表达水平都显著高于假孕组母犬下丘脑、卵巢和子宫中的瘦素水平(P<0.05),真孕和假孕母犬垂体中瘦素水平差异不显著(P>0.05).真孕母犬下丘脑、垂体、卵巢、子宫、输卵管组织中的瘦素受体的mRNA表达水平均显著高于假孕母犬对应各组织中的瘦素水平(P<0.05,表1).
2.2 瘦素及其受体在真孕和假孕母犬下丘脑-垂体-性腺轴各组织中的表达
免疫组织化学结果显示(图2~3),瘦素在真孕和假孕母犬的下丘脑、垂体、卵巢、子宫中均有表达,在输卵管中未检测到表达,瘦素受体在真假孕母犬的下丘脑、垂体、卵巢、子宫、输卵管组织中均有表达.其中,瘦素在真孕母犬卵巢的卵泡及卵巢间质细胞呈阳性表达,在假孕母犬卵巢阳性表达主要集中在间质细胞;瘦素受体在真假孕母犬卵泡及间质细胞均呈阳性表达,瘦素受体主要在真假孕母犬的输卵管皱襞表达阳性;瘦素及瘦素受体在真孕母犬子宫内膜、子宫腺中均呈阳性表达,而在假孕母犬子宫腺中表达阴性.
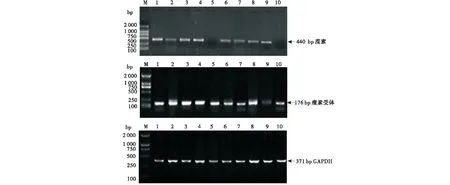
1~10分别代表:真孕母犬下丘脑、垂体、卵巢、子宫、输卵管、假孕母犬下丘脑、垂体、卵巢、子宫、输卵管.内参采用GAPDH (371 bp),Marker:DL2000 (100、250、500、750、1 000、2 000).Lane 1~5 represents hypothalamuses,pituitaries,ovaries,uteruses and fallopian tubes of pregnant bitches respectively,and lane 6~10 represents hypothalamuses,pituitaries,ovaries,uteruses and fallopian tubes of pseudo-pregnant bitches respectively.GAPDH was used as a control,whose length was 371 bp.DL2000 was used as DNA Marker.图1 瘦素(440 bp)和瘦素受体 (176 bp)及内参对照GAPDH(371 bp)在真孕和假孕母犬下丘脑-垂体-性腺轴各组织中的表达情况Figure 1 The expression of leptin and its receptor in various tissues of the hypothalamus-pituitary-gonad axis

表1 真孕组和假孕组母犬的各组织器官中瘦素及其受体mRNA的表达水平比较

A:真孕组下丘脑;B:假孕组下丘脑;C:真孕组垂体;D:假孕组垂体;E:真孕组卵巢;F:假孕组卵巢;G:真孕组输卵管;H:假孕组输卵管;I:真孕组子宫;J:假孕组子宫.A,C,E,G and I represent the tissues of pregnant bitches while B,D,F,H and J represent the tissues of pseudo-pregnant bitches.The images were magnified as 40×.A & B:hypothalamuses,C & D:pituitaries,E & F:ovaries,G & H:fallopian tubes,I & J:uteruses.图2 瘦素在真孕和假孕母犬下丘脑-垂体-性腺轴各组织中的表达(40×)Figure 2 The expression of leptin in various tissues of the hypothalamus-pituitary-gonad axis(40×)

A:真孕组下丘脑;B:假孕组下丘脑;C:真孕组垂体;D:假孕组垂体;E:真孕组卵巢;F:假孕组卵巢;G:真孕组输卵管;H:假孕组输卵管;I:真孕组子宫;J:假孕组子宫.A,C,E,G and I represent the tissues of pregnant bitches while B,D,F,H and J represent the tissues of pseudo-pregnant bitches.The images were magnified as 40×.A & B:hypothalamuses,C & D:pituitaries,E & F:ovaries,G & H:fallopian tubes,I & J:uteruses.图3 瘦素受体在真孕和假孕母犬下丘脑-垂体-性腺轴各组织中的表达(40×)Figure 3 The expression of leptin receptor in various tissues of the hypothalamus-pituitary-gonad axis
真孕母犬下丘脑、卵巢和子宫中的瘦素及其受体的表达量均显著高于假孕母犬(P<0.05),真孕母犬垂体和输卵管中瘦素受体的表达量显著高于假孕母犬(P<0.05).真孕母犬垂体中的瘦素表达水平与假孕母犬差异不显著(P>0.05,表2)

表2 瘦素及其受体在真孕和假孕母犬下丘脑-垂体-性腺轴各组织中的表达比较
3 讨论
瘦素已被证实是小鼠、牛等动物体内胚胎着床的关键因子[10-12],在妊娠小鼠及孕妇外周血中瘦素水平显著高于妊娠前和分娩后,且瘦素水平在孕妇体内下降有可能是妊娠即将终止的信号,测定孕妇血清中瘦素水平可以预测流产[13-14].对中国美利奴新疆军垦型细毛羊(单胎品种)和江苏湖羊(多胎品种)妊娠1~43 d外周血中的瘦素水平进行比较,结果显示江苏湖羊外周血中瘦素水平极显著高于中国美利奴新疆军垦型细毛羊,2种妊娠母羊的瘦素水平均高于空怀母羊,说明瘦素水平的含量不同直接影响绵羊是否怀孕以及怀单胎还是双胎[15].与我们前期研究结果相似近期有学者也发现妊娠母犬从排卵到分娩外周血中瘦素水平的浓度均高于未妊娠的母犬[16],多种动物妊娠与非妊娠期瘦素水平的对比结果显示瘦素对于母犬的妊娠具有一定的调节作用.
大多数的哺乳动物妊娠前期由卵巢上的黄体分泌孕酮维持妊娠,黄体退化后胎盘分泌孕酮继续维持妊娠,但目前尚无证据支持犬的胎盘能分泌孕酮,而且在母犬怀孕的任何阶段切除卵巢或黄体都会导致母犬流产[17].因此,犬的妊娠调控是由下丘脑-垂体-卵巢轴分泌激素相互作用的结果.在灵长类动物及小鼠下丘脑和垂体均发现了瘦素受体,瘦素通过直接或间接作用于下丘脑促性腺激素释放激素(GnRH)神经元或GnRH神经元主要调节因子kisspeptin (Kiss1)促进动物初情期启动及生殖系统的发育[18],瘦素缺乏会导致下丘脑-垂体-性腺轴功能异常[19-20].在小鼠垂体离体培养试验中[21]发现外源瘦素可以促进垂体促卵泡素(FSH)和促黄体素(LH)的释放且呈剂量依赖性,当FSH和LH达峰值后再提高瘦素浓度FSH和LH又下降,类似于垂体的负反馈作用,所以推测瘦素可以直接调控垂体功能也可能是先作用于下丘脑再促进垂体分泌促性腺激素.本研究发现在真孕母犬下丘脑和垂体均有瘦素及其受体的表达,且真孕组瘦素及其受体在下丘脑中表达显著高于假孕母犬,瘦素受体在垂体的表达显著高于假孕母犬,说明瘦素及其受体在犬胚胎着床期可能也是通过直接或间接作用介导了下丘脑-垂体-卵巢轴对于胚胎着床的调控.
给肥胖的雌鼠外源注射瘦素可明显增加卵巢重量及发育卵泡数量,而离体卵泡颗粒细胞与卵泡膜细胞培养时瘦素可以抑制颗粒细胞分化和卵细胞的成熟,说明不同的生理环境下瘦素对于卵巢具有不同的调节机制[22-23].本研究中瘦素在真孕母犬卵巢的卵泡及卵巢间质细胞呈阳性表达,在假孕母犬卵巢阳性表达主要集中在间质细胞,瘦素受体在真假孕母犬卵泡及间质细胞均呈阳性表达,说明瘦素可以直接作用于卵巢调控卵泡与间质细胞的功能.真孕组瘦素及受体在卵巢上表达量显著高于假孕组可能是瘦素直接或通过下丘脑垂体间接地促进卵巢分泌胚胎着床需要的激素.
瘦素促进人、小鼠、牛等动物胚胎着床的功能与瘦素受体在子宫内膜的表达密切相关,瘦素与子宫内膜上的受体结合,刺激子宫内膜发生变化为胚胎植入做好准备[10-13],也可能是瘦素先促进卵巢雌激素、孕激素的分泌,然后雌激素、孕激素又作用于子宫促进胚胎的着床.有学者发现瘦素及其受体的mRNA在妊娠期母犬的子宫和胎盘中有不同程度的表达,指出瘦素及其受体通过自分泌/旁分泌参与了犬胚胎着床的调控[24],与之相似,本研究中犬胚胎着床期瘦素及其受体在真孕母犬子宫内膜、子宫腺中表达阳性,且表达水平显著高于假孕母犬,说明瘦素在犬胚胎着床期可能也是通过自/旁分泌作用与子宫内膜上受体结合促进子宫与胚胎的结合.妊娠早期在雌、孕激素共同作用下子宫内膜增生,子宫腺分泌物增加为胚胎着床做好准备,胚胎着床期瘦素及其受体在假孕母犬子宫内膜腺中表达阴性说明假孕子宫内没有发生胚胎着床时子宫腺分泌功能增强等变化.Bartel等在母犬发情后期泡沫状子宫内膜上皮细胞(FEECs)发现了瘦素及其瘦素受体的存在,认为犬子宫内的泡沫状子宫内膜上皮细胞及表达的瘦素及其受体在早期妊娠中有潜在的作用[25].可见,瘦素及其受体通过作用于子宫对于母犬妊娠及胚胎着床具有一定的调节作用.
瘦素及其受体在人的输卵管中均有表达[26],但瘦素在真孕和假孕母犬输卵管中没有检测到表达,而瘦素受体主要在输卵管的黏膜皱襞上有表达,说明瘦素受体与犬输卵管的相关功能密切相关.本研究中瘦素及其受体在假孕母犬各组织中也有一定的表达,说明瘦素在假孕母犬体内也发挥着特定的生理作用.在非妊娠母犬黄体形成期瘦素及其受体的mRNA在卵巢中的表达有显著变化,表明瘦素对于非妊娠母犬黄体分泌孕酮及黄体退化有调控作用[27].可见,瘦素及其受体在母犬不同生理状态下发挥不同的功能.
通过比较瘦素及其受体在胚胎着床期真孕与假孕母犬下丘脑-垂体-性腺轴各组织中的表达差异,课题组推测真孕母犬外周血中高浓度的瘦素水平可能来自于下丘脑-垂体-性腺轴各组织的表达,外周血中瘦素水平能否作为判别真假孕的检测指标,其定量阈值是多少将在未来继续进行研究.
4 结论
胚胎着床期瘦素及其受体在真孕和假孕母犬下丘脑-垂体-性腺轴大部分组织中存在显著表达差异,表明瘦素及其受体对于犬的胚胎着床具有一定调节作用.本研究为瘦素在多种动物体内参与了下丘脑-垂体-性腺轴对于生殖的调控[28-30]提供了新证据.
