吡咯喹啉醌对鱼藤酮损伤细胞线粒体分裂融合的影响*
2021-09-16梁文鹏
梁文鹏,王 洁,张 琦*
(南通大学 教育部/江苏省神经再生重点实验室/神经再生协同创新中心,南通 226001)
帕金森病(Parkinson′s disease,PD)是一种常见于老年人的慢性神经系统退行性疾病,我国65 岁以上老年人患病率可达1.7%,且患病率随年龄的升高而增加。PD 的主要临床特征包括静止性震颤、运动迟缓、肌强直和自主神经功能障碍等,主要病理特征为黑质致密部多巴胺能神经元大量变性丢失以及路易小体(Lewy bodies)的出现。由于PD 病因不明,病理机制复杂,目前对于PD 的治疗只能改善和减轻症状,但不能有效地阻止病情进展[1]。
线粒体功能障碍在PD 的病理过程中具有重要的作用,线粒体功能的许多方面,包括活性氧产生、线粒体生物发生、线粒体动力学等都成为PD治疗的潜在靶点[2]。吡咯喹啉醌(pyrroloquinoline quinine,PQQ)是一种具有邻苯醌结构的氧化还原酶辅基,参与氧化还原酶的酶促反应,以往研究[3]发现PQQ 在体内能发挥清除自由基、保护神经元等作用。本课题组前期研究[4-5]也发现PQQ 对鱼藤酮损伤的神经元具有保护作用,能促进线粒体生物发生,提示PQQ可能通过调节线粒体功能发挥作用,在PD 的防治中具有潜在的应用价值。
线粒体分裂融合是维持线粒体动力学平衡的主要因素,分裂融合紊乱导致细胞内异常线粒体聚集,这是导致包括PD 在内的神经系统退行性疾病的病理机制之一[6]。线粒体的分裂融合是由动力相关蛋白1(dynamin-related protein 1,Drp1)、线粒体分裂蛋白(fission 1,Fis1)和线粒体融合蛋白1/2(mitofusin 1/2,Mfn1/2)、视神经萎缩蛋白1(optic atrophy 1,OPA1)等共同介导的[7]。鱼藤酮是线粒体复合物Ⅰ抑制剂,能选择性引起黑质多巴胺系统变性,被广泛用于PD模型的制备。鱼藤酮损伤抑制了线粒体分裂基因Drp1 和融合基因Mfn2 的表达,进而导致线粒体损伤[8-9]。慢性鱼藤酮损伤早期,线粒体分裂融合代偿性增加,损伤晚期分裂融合下降,而线粒体形态没有显著改变[10]。目前对于PQQ 如何影响线粒体分裂融合尚不清楚。
本研究采用鱼藤酮损伤SH-SY5Y 细胞模型,通过线粒体分裂融合相关基因和蛋白表达的检测以及形态学观察,分析PQQ 对线粒体分裂融合的调节,探讨PQQ 如何调节线粒体动力学对鱼藤酮损伤模型发挥保护作用,为将其开发并应用于PD 的临床治疗提供可靠的实验依据。
1 材料和方法
1.1 细胞模型构建 人神经母细胞瘤细胞(SH-SY5Y)细胞采用含有10%胎牛血清(fetal bovine serum,FBS)的DMEM 培养基,放于37 ℃、饱和湿度、5%CO2细胞培养箱中培养,隔天换液。细胞密度达90%以上即可进行传代。采用1、10、100 μmol/L PQQ(Sigma 公司)预处理细胞24 h 后,加入100 μmol/L 鱼藤酮(Sigma公司)损伤细胞,继续培养24 h 后收集细胞。分为正常对照组(Control)、单独PQQ 处理组(PQQ 100 μmol/L)、鱼藤酮损伤组(Rot 100 μmol/L)、PQQ 低剂量组(Rot+PQQ 1 μmol/L)、PQQ 中剂量组(Rot+PQQ 10 μmol/L)、PQQ 高剂量组(Rot+PQQ 100 μmol/L)。
1.2 总RNA 提取及定量聚合酶链式反应(polymerase chain reaction,PCR)采用Trizol 法提取SH-SY5Y 细胞总RNA 并进行逆转录。利用Primer 5软件设计Drp1 和Mfn2 的引物,分别为Drp1-F-gagtaagccctgaaccaa,Drp1-R-tgatgaaccgaagaatgag,Mfn2-F-accgccacatagaggaag,Mfn2-R-gcacagacacaggaagga。以18SrRNA 作为内参,引物序列18S-F-tagagggacaagtggcgttc,18S-R-cgctgagccagtcagtgt。定量PCR 反应体系为:SYBR green MIX 5 μL,上、下游引物各0.5 μL,cDNA 1 μL,ddH2O 3 μL,每个样本设3 个重复孔,生物学重复3 次,采用2-ΔΔCT法计算Drp1 和Mfn2 的相对表达量。
1.3 线粒体和胞质蛋白分离 采用线粒体分离试剂盒(碧云天公司)分离线粒体和胞质蛋白。细胞处理后用预冷的磷酸缓冲盐溶液(phosphate buffered saline,PBS)重悬、沉淀,加入1 mL 线粒体分离试剂进行悬浮、匀浆、离心等,获得的沉淀为线粒体,上清为胞质蛋白。
1.4 Western Blot 将上述收集获得的线粒体和胞浆蛋白采用聚氰基丙烯酸正丁酯(bicinchoninic acid,BCA)试剂盒(碧云天公司)进行浓度测定,调整蛋白质量浓度为1 μg/μL 后采用聚丙烯酰胺凝胶电泳(sodium dodecyl sulfatepolyacrylamide gel electrophoresis,SDS-PAGE)分离蛋白样品,转至聚偏氟乙烯(polyvinylidene fluoride,PVDF)膜后封闭,分别加入抗Drp1(1∶500)、Mfn2(1∶500)抗体4 ℃孵育过夜,二抗辣根过氧化物酶(horseradish peroxidase,HRP)-羊抗兔IgG(1∶5 000)、HRP-羊抗鼠IgG(1∶5 000)(Abcam 公司)室温孵育2 h,增强化学发光(enhanced chemiluminescence,ECL)显色、扫描,测量灰度值进行统计分析。分别采用电压依赖阴离子通道(voltage-dependent anion channel,VDAC)和β-actin(Sigma 公司)作为线粒体和胞浆蛋白的内参。
1.5 电镜检测 将上述处理的细胞置于4%戊二醛中于4 ℃固定2 h 以上,采用锇酸水溶液进行后固定2 h,乙醇梯度脱水后采用环氧丙烷和包埋剂包埋,半薄切片后用1%甲苯胺蓝溶液进行染色,在光镜下观察定位后进行超薄切片,厚度约70 nm,采用0.5%醋酸双氧铀染色2~3 min,柠檬酸铅复染2~3 min,最后在透射电子显微镜(Hitachi 公司)下观察线粒体形态。
1.6 统计学方法 采用GraphPad Prism 5 统计软件进行统计分析,多组比较采用单因素方差分析,并通过Tukey′s Multiple Comparison Test 进行两两比较,P<0.05 为差异有统计学意义。
2 结果
2.1 PQQ 对鱼藤酮损伤SH-SY5Y 细胞Drp1 和Mfn2 基因表达的影响 在不同浓度PQQ 预保护SH-SY5Y 细胞后,加入100 μmol/L 鱼藤酮损伤24 h,提取细胞总RNA,采用实时定量PCR 分析线粒体分裂和融合相关基因Drp1 和Mfn2 的表达变化。结果发现,单纯PQQ 作用不影响以上两个基因的表达,鱼藤酮损伤显著降低细胞中Drp1 和Mfn2 基因的表达,中(10 μmol/L)、高(100 μmol/L)剂量组PQQ 预保护均能拮抗鱼藤酮诱导的Drp1 和Mfn2 表达降低(图1A~B)。
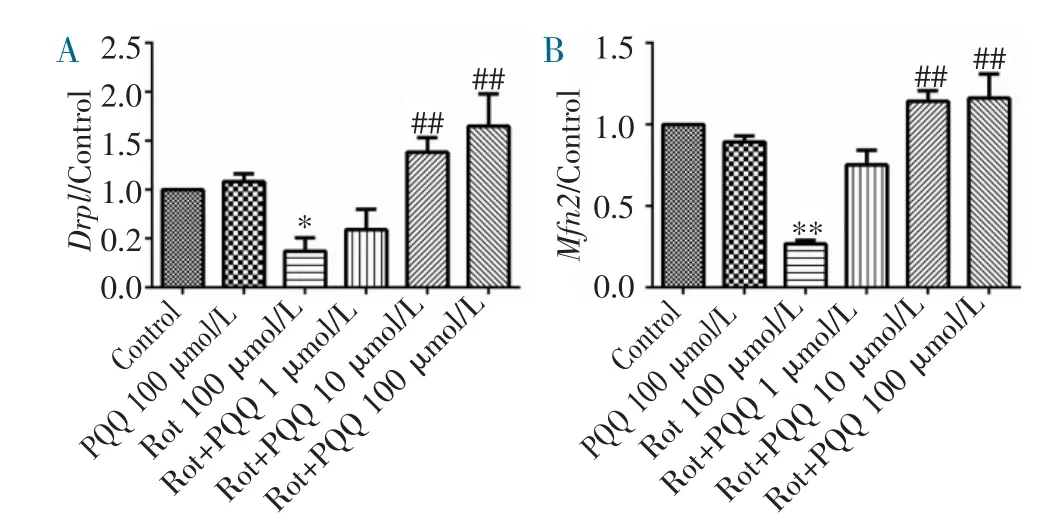
图1 定量PCR 检测PQQ 对鱼藤酮损伤SH-SY5Y 细胞Drp1 和Mfn2 基因表达的影响
2.2 PQQ 对鱼藤酮损伤SH-SY5Y 细胞Drp1 蛋白表达的影响 不同浓度PQQ 预处理鱼藤酮损伤的SH-SY5Y 细胞后,分离胞质和线粒体蛋白,采用Western Blot 检测发现,鱼藤酮损伤细胞的胞质中Drp1 表达增加,而在线粒体中表达显著下降(图2)。低、中、高浓度PQQ 预保护24 h 均能增加胞质中Drp1 的表达(图2A、C),高浓度PQQ 能同时显著增加线粒体中Drp1 的表达(图2B、D)。
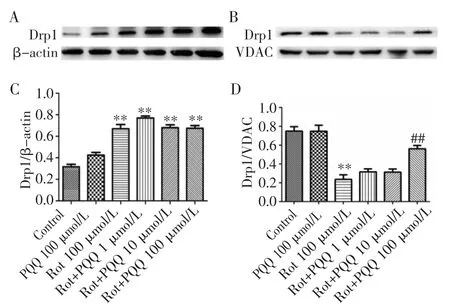
图2 PQQ 对鱼藤酮损伤SH-SY5Y 细胞Drp1 蛋白表达的影响
2.3 PQQ 对鱼藤酮损伤SH-SY5Y 细胞Mfn2 蛋白表达的影响 不同浓度PQQ 预处理鱼藤酮损伤的SH-SY5Y 细胞后,分离胞质和线粒体蛋白,采用Western Blot 检测发现,鱼藤酮损伤细胞的胞质中Mfn2表达没有显著变化,而在线粒体中表达显著下降(图3)。低、中、高浓度PQQ 预保护24 h 均能增加胞质(图3A、C)和线粒体(图3B、D)中Mfn2 的表达。
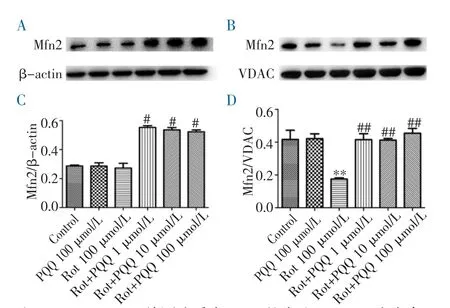
图3 PQQ 对鱼藤酮损伤SH-SY5Y 细胞Mfn2 蛋白表达的影响
2.4 PQQ 对鱼藤酮损伤SH-SY5Y 细胞线粒体形态的影响 采用透射电镜检测线粒体的超微结构,发现正常对照组线粒体数量较多,膜结构完整,线粒体大小在正常范围以内,单独使用PQQ 对细胞线粒体形态无显著影响,而鱼藤酮损伤组线粒体数量减少,可见异常肿胀和增大的线粒体,呈现病理性表型;不同浓度PQQ 处理后能减少病理性线粒体的发生(图4)。以上结果提示PQQ 能改善鱼藤酮损伤SH-SY5Y细胞线粒体动力学紊乱。
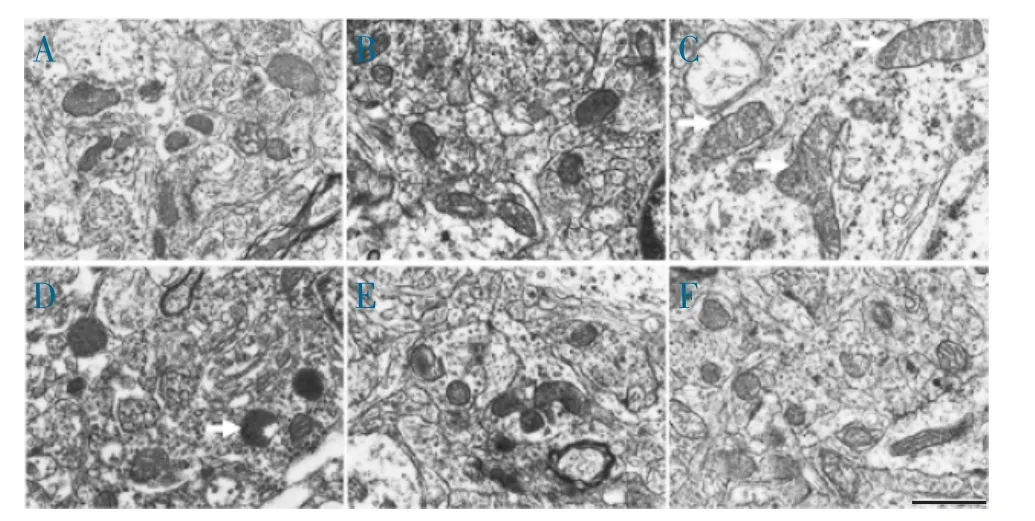
图4 透射电镜检测PQQ 对鱼藤酮损伤SH-SY5Y 细胞线粒体形态的影响
3 讨论
PD 是发病率仅次于阿尔茨海默病的一种神经退行性疾病,其病因可能与环境、遗传、氧化应激等相关。尽管目前临床上PD 的治疗方法不断进步,但由于发病机制不清,仍无法根治,开发能针对PD 发病机制的治疗方法迫在眉睫。
线粒体是一种处于融合与分裂动态平衡中的细胞器,是线粒体重要的质量控制机制,通过融合保留膜电位较高、质量相对较好的线粒体,通过裂解及自噬清除膜电位较低、质量相对较差的线粒体,细胞存活和分化过程中通过线粒体这种适应性改变在维持细胞功能和细胞活力方面起着重要作用[6]。家族性PD相关基因的致病性突变通常会导致线粒体动力学异常,在PD 动物模型中也发现神经元中存在肿胀和增大的线粒体,提示PD 的发病机制可能与线粒体分裂融合紊乱有关[11-12]。因此,线粒体动态失衡可能作为PD治疗的一个新靶点[13]。
研究[14]发现在小鼠多巴胺能神经元中敲除Drp1导致尾状壳核多巴胺能神经终末消失,中脑神经元胞体退行性改变,同时线粒体含量显著下降,尤其在轴突中线粒体动态失衡。本研究发现在鱼藤酮损伤细胞模型中Drp1 表达下降,并发生了从线粒体到胞质的转位,推测其不能及时有效分裂受损线粒体,而PQQ 预保护则能显著增加Drp1 的表达,尤其在线粒体中发挥清除损伤线粒体的功能。
研究[11]表明,过表达Mfn2 能阻断线粒体断裂,在体外保护神经元免受百草枯诱导的线粒体功能障碍,抑制黑质和纹状体轴突终末多巴胺能神经元选择性丢失。在不同细胞系中降低Mfn2 可导致线粒体增大和管状网络的破坏[15],Mfn2 可通过改善细胞凋亡来阻止鱼藤酮诱导的细胞死亡,从而发挥保护作用[16]。本研究也表明,PQQ 可显著增加Mfn2 的表达,并由此调节线粒体动力学,发挥神经保护作用。
此外,促进线粒体生物合成也能调节线粒体分裂融合,对鱼藤酮引起的神经毒性具有保护作用[9]。以往研究[4]发现PQQ 可以促进线粒体生物发生,并由此拮抗鱼藤酮损伤,可见线粒体生物发生和分裂融合之间的相互作用也参与了神经系统退行性变的发生。PQQ 有可能通过促进线粒体生物发生影响线粒体分裂融合过程,从多方面发挥神经保护作用,但需要进一步研究证实。
综上所述,本研究在体外采用鱼藤酮损伤SHSY5Y 细胞模型,初步探究了PQQ 对鱼藤酮诱导PD模型中线粒体分裂融合的调控,提示PQQ 有可能通过调节线粒体动态平衡对鱼藤酮诱导的PD 模型发挥保护作用,为进一步将PQQ 用于临床治疗提供了基础理论依据,也使线粒体分裂融合成为PD治疗新的靶点。
