不同连作年限菠萝园土壤差异代谢物和细菌群落结构 分析
2021-09-14刘传和贺涵何秀古刘开邵雪花赖多匡石滋肖维强
刘传和 贺涵 何秀古 刘开 邵雪花 赖多 匡石滋 肖维强
(1. 广东省农业科学院果树研究所/ 农业农村部南亚热带果树生物学与遗传资源利用重点实验室/ 广东省热带亚热带果树研究重点实验室, 广州 510640;2. 广东省农业科学院,广州 510640)
连作障碍制约着果树产业的可持续发展,化感自毒作用是引起果树连作障碍的主要原因之一[1]。有机酸、直链醇、酚酸、萜类和生物碱等代谢物多数属于自毒物质[2],其中萜类和酚酸类物质是导致土壤化感自毒作用的关键代谢物[3]。作物长期连作后根际分泌的苯甲酸、羟基苯甲酸和肉桂酸等酚酸类代谢物通过与土壤根际微生物互作而不断积累,阻碍土壤有机质的分解和矿化,妨碍了根系对土壤养分的吸收[4-7],导致自毒作用加剧。
土壤细菌群落作为土壤生态环境的重要组成部分[8],其群落结构的变化与连作密切相关。在大豆、烟草、茶树、香榧等作物连作后土壤研究表明,连作后以糖类、氨基酸类碳源利用为主的细菌数量显著减少,根际土壤细菌碳源利用率下降[9]。连作导致土壤中细菌种类减少、细菌多样性降低,土壤细菌群落多样性和丰富度指数下降,土壤微生物类型逐渐由“细菌型”向“真菌型”转变,土壤微生物出现失衡[8,10-14]。在门水平上分析表明连作后土壤的优势菌门厚壁菌门(Firmicutes),以及有益菌门变形菌门(Proteobacteria)、拟杆菌门(Bacteroidetes)和放线菌门(Actinobacteria)等所占比例逐渐下降[12,15]。土壤细菌根瘤菌目和酸微菌目的相对丰度,以及分枝杆菌属(Mycobacterium)、藤黄单胞菌属(Luteimonas)、芽单胞菌属(Gemmatimonas)、浮霉菌属(Planctomyces)细菌的多样性因连作而减少[13-14]。
菠萝(Ananas comosus(L.)Merr.)又称凤梨,是著名的热带水果。据统计,我国现有菠萝种植约97.1万亩,产量约173.3万t[16]。我国菠萝消费量约以年均7.5%速度在增长,市场需求量大,菠萝产业为我国热区经济社会发展做出了重要贡献。菠萝连作在我国各产区十分普遍。果农在菠萝果实收获后,将茎叶粉碎还田翻松土壤后再种植菠萝[17-18],并施用大量化肥,循环往复,常连续种植10余年甚至更长。随着连作年限的增加,菠萝果实产量、品质明显降低,产业发展受到影响。有关菠萝连作对土壤代谢物及细菌群落结构的影响变化等仍不清楚。
本研究基于非靶向代谢组学及16S rDNA高通量测序技术,以菠萝园边角地未耕作过的土壤为对照,对连续种植5 a和15 a菠萝的土壤代谢物和土壤细菌群落结构进行比较分析,筛选出不同连作年限菠萝园土壤中的差异代谢物及代谢通路,分析细菌群落组成及丰度变化。本研究旨在探讨菠萝园土壤代谢物及细菌群落结构的变化与菠萝连作年限的关系,为菠萝连作相关研究奠定基础,也为菠萝的田间管理、土壤环境保护等提供参考。
1 材料与方法
1.1 材料
本研究取样区域选择在我国最大的菠萝主产区广东省湛江市徐闻县。本次采集的土壤样品来自徐闻县菠萝的主要种植区曲界镇,土壤类型为砖红壤。
选择连续种植了菠萝5 a、15 a的菠萝园土壤(分别简写成T5、T15)及菠萝园边角地未耕作过的土壤(T0,对照)作为研究对象。菠萝种植园每造均用吸芽苗种植,种植品种为“巴厘”,亩植4 000棵,果实收获后菠萝茎叶粉碎还田。每造菠萝亩施尿素100 kg、复合肥(N15-P2O515-K2O15)150 kg,分别于种植后的第4个月(尿素50 kg、复合肥50 kg)和第8个月(尿素50 kg、复合肥100 kg)分两次施完。
本研究土壤取样于2020年3月中旬进行。每个种植年限选择3个园区(园区间隔 100 m 以上)。所有种植菠萝的园区坡向、坡度和管理措施基本一致。取样时菠萝种植园当造菠萝未施首次大肥,每个园区随机选择东部、西部2个位置作为取样对象。每个位置随机选取两个取样点(相距5 m)。取样时先除去地表5 cm厚的表层土壤,采用内径为5 cm土钻采集0-20 cm土层样品,每个取样点平行钻取2钻;两个取样点的土样混为1个样品,共18个样品,每处理6次重复。取样后用离心管装好立即置于液氮中速冻2 h后在-70℃超低温冰箱中暂存。
1.2 方法
1.2.1 土壤代谢物的测定
1.2.1.1 土壤代谢物的提取 (1)取样本1 000 mg于5 mL EP管中,加入1 000 μL提取液(甲醇∶水=3∶1,V/V),再加入1 000 μL乙酸乙酯及5 μL 核糖醇,涡旋30 s;(2)加入钢珠,35 Hz研磨仪研磨4 min,超声5 min(冰水浴);(3)将样本于4℃离心,10 000 r/min离心15 min;(4)移取上清液于5 mL EP管中,再加入1 000 μL提取液(甲醇∶水=3∶1,V/V)和1 000 μL乙酸乙酯,重复以上2、3步骤,合并所有上清液;(5)在真空浓缩器中干燥提取物;(6)向干燥后的代谢物中加入20 μL甲氧胺盐试剂(甲氧胺盐酸盐,溶于吡啶20 mg/mL),轻轻混匀后,放入80℃烘箱中孵育30 min;(7)向每个样品中加入30 μL BSTFA(含有1% TMCS,V/V),将混合物在70℃孵育1.5 h;(8)冷却至室温,向混合的样本中加入5 μL FAMEs(溶于氯仿);(9)上机检测。
1.2.1.2 代谢物GC-TOF-MS检测 Agilent 7890气相色谱-飞行时间质谱联用仪配有Agilent DB-5MS毛细 管 柱(30 m×250 μm×0.25 μm,J&W Scientific,Folsom,CA,USA),GC-TOF-MS具体分析条件如表1。
1.2.1.3 代谢物测定数据处理与分析 使用ChromaTOF软件(V4.3x,LECO)对质谱数据进行处理[19],使用LECO-Fiehn Rtx5数据库,将质控(Quality Control)样本中检出率50%以下或RSD>30%的峰去除[20]后的有效数据用于后续实验分析。
采用R(3.3.2)包ropls进行正交偏最小二乘法-判别分析(orthogonal projections to latent structures- discriminant analysis,OPLS-DA)模型计算,获取更可靠的代谢物的组间差异信息;采取将差异倍数(fold change)、t检验的P值(P value<0.05)和OPLS-DA模型的VIP值(variable importance in the projection,VIP>1)——变量投影重要度相结合的方法筛选差异代谢物;采用单因素方差(ANOVA)分析绘制的火山图(volcano plot)统计差异代谢物在T0-T5和T0-T15比较组中的表达水平;采用KEGG(Kyoto Encyclopedia of Genes and Genomes)对差异代谢物进行通路富集分析。
1.2.2 微生物16S rDNA测序分析 微生物16S rDNA的扩增及测序委托北京百迈客生物科技有限公司完成。PCR扩增引物为:(16SF)AGRGTTTGATYNTGGCTCAG,(16S-R)TASGGHTACCTTGTTASGACTT;PCR扩 增 程 序:95℃预变性5 min;95℃变性30 s,55℃退火30 s,72℃延伸90 s,30个循环;72℃延伸7 min,4℃保存。
采 用PacBio测 序 平 台,对CCS(Circular Consensus Sequencing)序列进行Barcode识别和长度过滤,得到Optimization-CCS,使用Usearch软件[21]对Tags在97%的相似度水平下进行聚类并划分OTU,根据OTU的序列组成得到其物种分类,基于OTU分析结果,对样品在各个分类水平上进行分类学分析,获得各样品在门、属分类学水平上的物种分布图与在目水平上的物种聚类热图。稀释性曲线(dilution curve)用于检验测序数据量[22];ACE、Chao1、Shannon及Simpson指数用于分析细菌菌群的多样性及丰度;非度量多维标定法(Non-MetricMulti-Dimensional Scaling,NMDS)[23]分析细菌菌群结构差异;斯皮尔曼(Spearman)相关性分析物种在环境样本中的共存关系。通过典范对应分析(canonical correspondence analysis,CCA)[24]在属水平细菌菌群和pH等理化指标之间的相关性,样品射线与pH射线成锐角即为正相关,成钝角为负相关。
2 结果
2.1 不同连作年限菠萝园土壤样本OPLS-DA法判别分析
图1显示,不同连作年限菠萝园土壤微生物代谢物模型参数中R2X=0.776,R2Y=0.979,Q2Y=0.888,说明不同连作年限菠萝园土壤代谢模型对自变量X的解释程度为77.6%;对分类变量Y的解释程度为97.9%,对样本变量的预测程度为88.8%,该模型R2Y非常接近1,表明OPLS-DA模型为有效模型,样本数据真实可靠度较高、稳定性和预测能力较强。

图1 不同连作年限菠萝土壤所有样本OPLS-DA得分图Fig.1 OPLS-DA scores of all soil samples from the pineapple orchard in different continuous-cropping years
OPLS-DA模型能直观的展示出组间的分离情况,18个样本均处于95%置信区间内,T5、T15组间接近且有部分重叠但与对照明显分开,表明T5和T15组间代谢模式接近,但均与对照存在显著差异。组内各样本聚集,分散程度较小。
2.2 不同连作年限菠萝园土壤差异代谢物火山图分析
图2分别显示了T5组、T15组与对照组(T0)间的差异代谢产物分布情况。由火山图可知,T5组与T0组土壤间有120个差异代谢物,其中显著增加(P<0.05)的有119个,显著降低(P<0.05)的有1个;T15组与T0组土壤微生物中有80个差异代谢物,其中显著增加(P<0.05)的有60个,显著降低(P<0.05)的有20个。

图2 差异代谢物火山图Fig.2 Volcano plot of differential metabolites
2.3 显著差异代谢物筛选
运用OPLS-DA模型的分析方法,将差异倍数(FC>2)、t检验P值(P value <0.05)和VIP值(VIP >1)相结合的标椎对T0-T5比较组和T0-T15比较组差异代谢物进行筛选,共筛选出11个具有显著性差异(P <0.05)的代谢物(表2)。其中,有机酸4种,氨基酸及其衍生物2种,糖类3种,醇类2种。
据表2可知,T0-T5比较组和T0-T15比较组差异代谢物的组分含量存在差异。与T0相比,T5中的有机酸类代谢物(2R,3S)-2-羟基-3-异丙基丁二酸、4-羟基苯甲酸、4-羟基-3-甲氧基苯甲酸、3,4-二羟基苯甲酸的表达分别升高了1.924倍、1.538倍、1.569倍和1.740倍;而在T15中,与T0相比这4种代谢物的表达分别提高了2.099倍、2.191倍、1.879倍和1.746倍。类似地,T5中N-乙酰基-β-丙氨酸、N-氨基甲酰谷氨酸的表达分别较T0升高了2.211倍和2.327倍;而在T15中N-乙酰基-β-丙氨酸、N-氨基甲酰谷氨酸的表达分别较T0升高了3.186倍和3.440倍。

表2 不同比较组间差异代谢物Table 2 Differential metabolites among the comparison groups
与T0比较,T5中的糖类、醇类代谢物表达提高的倍数高于T15中糖类、醇类代谢物含量提高的倍数。T5中的葡萄糖、槐糖、松三糖的含量水平分别比T0升高了4.878倍、4.201 倍和1.292倍;而在T15中葡萄糖的含量水平比T0提高了1.580倍,槐糖、松三糖的含量则分别降低了3.892倍和1.386倍。类似地,与T0相比,T5中的醇类代谢物(3β,5α,6β)-胆固醇3,5,6-三醇、山梨糖醇含量提高了3.071倍和2.244倍;而T15中这2种代谢物的含量水平分别提高了1.568倍和1.901倍。
2.4 差异代谢物KEGG富集分析
2.4.1 基于HMDB数据库的差异代谢物统计 如图3所示,T0-T5比较组在HMDB数据库检索到的差异代谢物种类有8种,T0-T15比较组检索到的差异代谢物种类有6种。在T0-T5比较组和T0-T15比较组中同时检索到的差异代谢物有5种,分别为脂肪酰基、苯及其取代衍生物、有机氧化合物、类固醇和类固醇衍生物、肉桂酸及其衍生物。其中,苯及其取代衍生物、肉桂酸及其衍生物、类固醇和类固醇衍生物在T0-T5比较组和T0-T15比较组所占比例相同。在T0-T5组检索到的有机氧化合物类代谢物有3个,在T0-T15组有2个;T0-T5组检索到的脂肪酰基类代谢物有8个,在T0-T15组有3个。此外,仅在T0-T5组中检索到的差异代谢物有3种,分别为吡啶及其衍生物、非金属氧阴离子化合物和羧酸及其衍生物;仅在T0-T15组中检索到的差异代谢物有1种,为二嗪类代谢物。

图3 T0-T5比较组和T0-T15比较组差异代谢物HMDB分类图Fig.3 HMDB classification map of differential metabolites from the comparison groups of T0-T5 and T0- T15
2.4.2 差异代谢物代谢通路分析 KEGG通路富集分析表明,显著差异代谢物主要富集在11条代谢通路上(表3),分别为果糖和甘露糖代谢、类固醇生物合成、苯丙烷生物合成、泛醌和其他萜类醌的生物合成、酪氨酸代谢、苯丙氨酸代谢、苯丙氨酸、酪氨酸和色氨酸的生物合成、亚油酸代谢、叶酸生物合成、异喹啉生物碱的生物合成和不饱和脂肪酸的生物合成。
表3结果显示,这11条通路所对应的代谢物有8个。3,4-二羟基苯甲酸主要参与苯丙氨酸、酪氨酸和色氨酸的生物合成。4-羟基苯甲酸和4-羟基肉桂酸共同参与苯丙氨酸代谢和泛醌及其他萜类醌的生物合成途径;并且,4-羟基苯甲酸还参与叶酸生物合成途径;而4-羟基肉桂酸参与苯丙烷和异喹啉生物碱的生物合成途径,此外,4-羟基肉桂酸还可单独参与酪氨酸的代谢过程。山梨糖醇主要参与果糖和甘露糖代谢过程,胆固醇主要参与类固醇生物合成途径;亚油酸既可单独参与亚油酸代谢途径,又可与山嵛酸和木质酸一起共同参与不饱和脂肪酸的生物合成途径。

表3 差异代谢物代谢通路分析Table 3 Metabolic pathway analysis of the differential metabolites
2.4.3 KEGG富集网络图 KEGG富集网络图(图4)更直观显示了T5、T15差异代谢物所参与的关键代谢通路,且每条关键通路多为一种以上的代谢物所共同参与的,同一条通路的代谢物组成有明显差异。由图4可知,不饱和脂肪酸的生物合成途径在T5、T15代谢中的影响均最大,但参与不饱和脂肪酸的生物合成的代谢物有差异,在T5中受到油酸、山嵛酸、花生酸、亚油酸和木质酸共同的影响,而在T15中主要受到亚油酸、木质酸和山嵛酸的影响;其次,苯丙氨酸代谢途径在T5中的影响较大,泛醌及其他萜类醌的生物合成途径在T15中的影响较大,而这两种代谢途径均是由4-羟基苯甲酸和4-羟基肉桂酸所共同参与的。此外,图4-A显示T5对氧化磷酸化途径的代谢影响较大;图4-B显示T15对β-丙氨酸代谢、苯丙氨酸、酪氨酸、色氨酸的生物合成和亚油酸代谢的影响较大。以上结果表明,T5、T15代谢水平存在一定的差异,显著差异代谢物在T5、T15同一条代谢通路中的影响也有所不同。

图4 差异代谢物KEGG富集网络图Fig. 4 KEGG pathway enrichment map of differential metabolites
2.5 不同连作年限菠萝园土壤细菌16S测序数据
对T0及T5、T15土壤进行16S全长测序(6次重复),分析了细菌群落的多样性。通过Barcode识别后共获得122 534条CCS序列,每个样品至少产生3 889条CCS序列,平均产生6 807条CCS序列,过滤后共获得121 795条(99.47%)有效CCS序列用于后续实验分析,样品平均序列长度为1 447 bp,在97%的相似水平下进行聚类分析,共获得OUT 9582个。由图5可知,当测序量接近5 000读长时,稀释性曲线(dilution curve)逐渐趋于平缓,表明本试验样品序列充足,取样能够真实地反映菠萝地土壤中的细菌群落结构多样性。
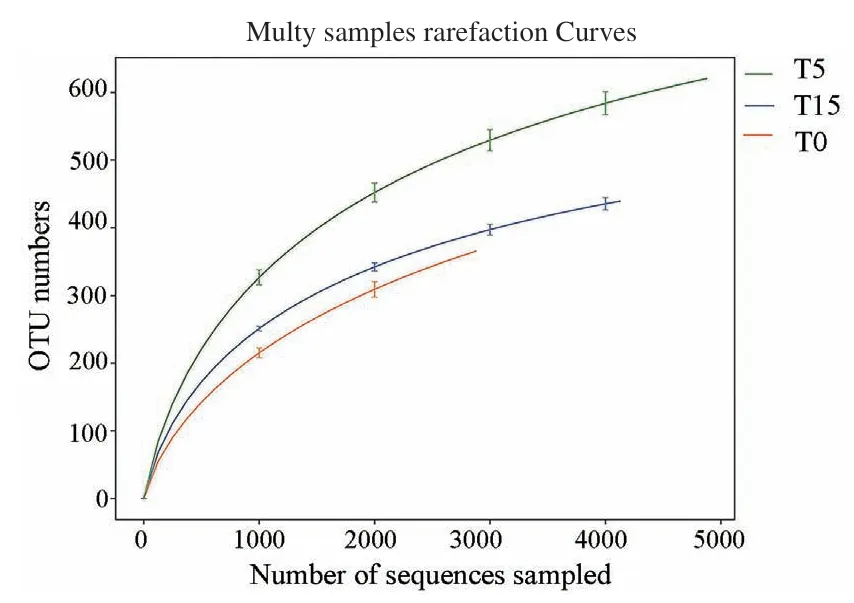
图5 不同连作年限菠萝园土壤样品的稀释性曲线Fig. 5 Dilution curve of soil samples in the different continuous-cropping years of the pineapple orchard
如图6,细菌在OTU水平上的NMDS分析结果显示,T5、T15土壤细菌群落分布比T0更集中,且T5、T15之间细菌群落明显分开,说明不同种植年限菠萝地土壤细菌群落结构存在显著差异。

图6 不同连作年限菠萝园土壤细菌群落NMDS分析Fig. 6 NMDS analysis of different soil bacterial community in the different continuous-cropping years of the pineapple orchard
2.6 不同连作年限菠萝园土壤细菌群落多样性和相对丰度变化
在门水平上对细菌群落相对丰度进行分析,如图7-A所示,从土壤中鉴定到的细菌主要来自6个门,约占细菌总数的67.72%,分属放线菌门(Actinobacteriota)、变形细菌门(Proteobacteria)、绿弯菌门(Chloroflexi)、酸杆菌门(Acidobacteriota)、浮霉菌门(Planctomycetota)、厚壁菌门(Firmicutes)。其中放线菌门(Actinobacteriota)、变形细菌门(Proteobacteria)、绿弯菌门(Chloroflexi)、酸杆菌门(Acidobacteriota)、浮霉菌门(Planctomycetota)为菠萝地土壤的优势菌群,这些菌在T5、T15土壤中分别占细菌总数的70.51%和89.82%。图7-A显示,T5、T15土壤中厚壁菌门(Firmicutes)所占比例较T0明显减少,放线菌门(Actinobacteriota)、绿弯菌门(Chloroflexi)和浮霉菌门(Planctomycetota)所占比例显著增加。变形细菌门(Proteobacteria)和酸杆菌门(Acidobacteriota)在T5土壤中所占比例最高,但在T15土壤中比例降低,低于T0。
图7-B表明,从属水平上分析T5、T15土壤细菌群落的相对丰度明显高于T0。与T0相比,T5土壤在属水平上相对丰度显著增加的细菌群落主要有伍氏束缚菌(Conexibacter)、Thermosporothrix、Aciditerrimonas、Acidibacter、Fimbriiglobus、Actinoallomurus;T15土壤显著增加的属有分枝杆菌属(Mycobacterium)、中华单胞菌属(Sinomonas)、Conexibacter、Thermosporothrix、Aciditerrimonas、Fimbriiglobus、Actinoallomurus。但T5、T15土壤伯克氏菌属(Burkholderia)的相对丰度较T0明显降低。
图7-C所示为在目水平上T0与T5、T15土壤细菌群落相对丰度的差异。与T0相比,T5土壤中显著增加的细菌目有Azospirillales、黄色单胞菌目(Xanthomonadales)、Polyangiales、Pedosphaerales、Isosphaerales、Solibacterales、酸杆菌目(Acidobacteriales)、芽单胞菌目(Gemmatimonadales)、Gammaproteobacteria-Incertae-Sedis、细链孢菌目(Catenulisporales);显著减少的细菌目有链霉菌目(Streptomycetales)、假诺卡式菌目(Pseudonocardiales)。而T15土壤相对丰度显著增加的细菌目有乳杆菌目(Lactobacillales)、弗 兰 克 氏 菌 目(Frankiales)、土壤红杆菌目(Solirubrobacterales)、黏 球 菌 目(Micrococcales)、酸 微 菌 目(Acidimicrobiales)、Streptosporangiales、芽孢杆菌目(Bacillales)、棒杆菌亚目(Corynebacteriales)、Gaiellales;显著减少的细菌目有根瘤菌目(Rhizobiales)、链霉菌目(Streptomycetales)、假诺卡式菌目(Pseudonocardiales)。T5、T15相对丰度均显著减少的细菌目有链霉菌目(Streptomycetales)和假诺卡式菌目(Pseudonocardiales)。

图7 不同连作年限菠萝园土壤细菌群落在门、属、目水平上的相对丰度Fig. 7 Relative abundance of different soil bacterial community at phyla,genus and order level in the different continuouscropping years of the pineapple orchard
2.7 不同连作年限菠萝园土壤细菌群落多样性 分析
表4所示,T5土壤的ACE和Chao1指数均显著高于T15和T0;而T15土壤的ACE和Chao1指数均低于T5和T0。结果表明T5土壤细菌相对丰度升高,T15细菌群落相对丰度下降。如表4所示,T5、T15的Simpson指数分别为0.01和0.04,显著低于T0(0.12);而T5、T15的Shannon指数分别为5.39、4.66,显著高于T0(3.90)。结果表明T5、T15土壤中细菌群落多样性增加,且以T5土壤中细菌群落多样性相对更高。

表4 不同连作年限菠萝园土壤细菌丰度与多样性Table 4 Diversity of relative abundance of different bacterial of soil under different continuous-cropping years of the pineapple orachard
2.8 不同连作年限菠萝地土壤细菌群落与土壤pH的相关性
由图8可知,连作年限与土壤pH成负相关;细菌属(Aciditerrimonas)、Acidibacter、Actinoallomu- rus、Paraburkholderia与pH成正相关,分枝杆菌属 (Mycobacterium)、Gaiella、中 华 单 胞 菌 属(Sino- mo-nas)、Conexibacter、Thermosporothrix与pH成 负 相关。

图8 不同连作年限菠萝园土壤中细菌群落与pH值的相关性CCA分析Fig. 8 CCA analysis correlating bacterial community to pH value of soil under different continuous-cropping years of the pineapple orchard
3 讨论
糖类作为微生物的主要碳源,为微生物正常代谢提供所需要的能量。已有研究表明,大豆连作导致葡萄糖的组分含量下降,同时,随连作时间的延长,土壤多糖含量逐渐降低,影响土壤有机质的组成及植株的生长发育[25-26]。这与本研究中,连续种植了15年菠萝的土壤中槐糖、松三糖的表达水平降低的结果相似。
酚酸类代谢物是多数作物的主要化感物质,其自毒潜力随种植年限的增加而有上升的趋势[3,27]。本研究表明,菠萝连作后土壤中的4种有机酸类代谢物的表达水平均升高,尤其是在菠萝连作15年的土壤中酸类代谢物的表达升高幅度更大。由于长期连作,施肥量大,有机酸类物质增加,造成土壤逐渐酸化,不仅容易滋生出大量好酸性微生物,也加剧了土壤的自毒作用[27-32]。菠萝连作5年、15年后土壤中4-羟基苯甲酸等酚酸类物质的含量水平均提高,且连作15年提高的幅度更大,表明菠萝长期连作对土壤环境产生了不利影响。在蚕豆、花生等的连作障碍研究中发现,肉桂酸、苯甲酸和对羟基苯甲酸等自毒物质是导致连作障碍的重要影响因 子[3,30]。茄子连作障碍的研究也表明,连作后土壤中的肉桂酸浓度增加,加剧了茄子的自毒作用[33]。连作后土壤中的对羟基苯甲酸含量增加影响了土壤微生物活性与多样性,导致土壤中的细菌、放线菌数量减少,真菌数量增加[34],土壤中有益生物减少、有害生物增加,最终导致连作障碍的发生[35]。
KEGG代谢通路分析表明,不同连作年限菠萝地土壤差异代谢物主要富集在不饱和脂肪酸的生物合成途径等11条代谢通路中,并且每条关键通路多为一种以上的代谢物所共同参与的。不饱和脂肪酸的生物合成途径对连作5年、15年菠萝土壤代谢的影响均最大,该途径是由亚油酸、山嵛酸和木质酸等有机酸共同参与的代谢途径,这可能与连作导致土壤酸化,有机酸参与的代谢活动增强有关[36-37]。
土壤细菌代谢旺盛、繁殖快,种类多、数量大,是土壤微生态环境质量的重要组成部分,对作物的生长有着重要的影响。研究发现,细菌群落的变化与连作障碍关系密切[12,38]。本研究通过微生物16S测序对细菌群落结构分析结果表明,与对照相比,连续种植菠萝5年和15年的土壤细菌群落多样性提高,但连续种植菠萝15年的土壤细菌群落的相对丰度降低,这与其他作物连作相关研究结果相 似[12,39]。土壤pH值在细菌群落结构形成中起着重要作用,是决定细菌群落结构演替与群落结构发生分异的主要驱动因子,土壤pH与土壤细菌群落的相关性最高[38,40-41]。基于CCA分析结果表明,土壤pH与菠萝连作年限呈负相关,这不仅为菠萝长期连作后导致土壤酸化做出了解释,也为菠萝连作后导致土壤细菌群落减少,土壤pH变化在调节细菌群落发生分异中的作用提供了有利的证明。长期种植单一作物导致土壤微生物多样性出现不同程度的下降,根际微生态环境受到破坏[14],需要采取减施化肥、轮作、休耕等措施控制土壤的酸化与盐分积累,提高微生物群落尤其是细菌群落的多样性,提高土壤生态系统的稳定性[42-43]。
4 结论
菠萝连作后土壤中的酸类代谢物表达水平升高。连续种植5年菠萝的土壤中葡萄糖、槐糖、松三糖等糖类代谢物的表达水平升高。连续种植了15年菠萝的土壤中槐糖、松三糖的表达水平降低;葡萄糖的表达水平虽高于菠萝园边角地未耕作过的土壤,但低于连续种植5年菠萝的土壤中葡萄糖的表达水平。细菌16S测序表明,连续种植菠萝5 年和15 年的土壤细菌群落结构存在明显差异;连作5年、15年土壤细菌群落多样性提高,但连作15年的土壤细菌群落相对丰度降低。
