葡萄AKR基因家族的鉴定和组织特异性表达分析
2021-09-14马亚男卢旭魏云春李康魏若男李胜马绍英
马亚男 卢旭 魏云春 李康 魏若男 李胜 马绍英
(1. 甘肃农业大学生命科学技术学院,兰州 730070;2. 甘肃农业大学园艺学院,兰州 730070;3. 甘肃农业大学基础实验教学中心, 兰州 730070)
在植物的生长发育过程中,不可避免地会遇到生物或非生物胁迫的影响,例如,土壤盐碱含量过高、极端温度、干旱和病虫害等[1-2]。研究表明,非生物胁迫下植物体内活性乙醛和活性氧(ROS)的水平会提高并导致植物受到损伤。为抵御不良环境植物会产生大量氧化还原酶,将ROS、活性乙醛等有毒物质分解为毒性较小的乙醇与碳水化合物,其中醛酮还原酶(AKR)家族发挥着重要作用[3]。
AKR是氧化还原酶中的组成部分,属于单体胞质蛋白,并以NADP(H)为辅酶氧化还原羧基底物,在生物体中起着核心作用[4-5]。目前关于AKR催化机制的研究的已有较多报道[6-7]。AKR家族成员均以NADPH作为辅酶,由具有相似的三维结构和还原羰基基团的蛋白酶组成[8-9]。AKR催化反应时,辅酶NADPH首先结合到AKR上,接着底物再结合至AKR上。催化反应结束后,产物首先释放,然后氧化态辅酶再释放出来。其催化底物主要有脂肪族和芳香族,而芳香族又分为单糖、酮基、醛基、类固醇等[4-5]。AKR在结构上约有320个氨基酸残基,约35 kD的长度,在进化中具有高度保守性。ARK保守结构域是经典的(α/β)8桶状折叠结构,特异性主要由(α /β)8桶状结构羧基端的3个环(LooP A、LooP B、LooP C)的不同组成和大小决定[10]。AKR催化活性位点是由 N 末端催化四联体(Asp50、Tyr55、Lys84)所构成[10]。该催化四联体被认为是AKR负责底物催化的关键氨基酸[11]。另有研究发现,C末端形成的高度保守的氨基酸残基(Ser271、Arg276)在辅基结合过程中起重要作用[10]。在正常条件下,AKR在植物生长过程中通过参与调控细胞的生长和分化发挥重要的功能[12]。当植物遭遇逆境胁迫时,AKR一方面会催化醛、酮、类固醇以及其他羰基化合物的还原活性缓解外界胁迫的压力[3],另一方面会通过调控次级代谢物合成、维持细胞渗透压来抵御逆境胁迫[13]。
在植物中,已有研究人员从拟南芥[14],番茄[15],甘蓝[16],白菜[17]及棉花[18]等物种克隆得到AKR基因,并发现AKR 基因参与植物细胞生长发育过程中许多氧化还原反应,可增强植物对于逆境胁迫的抵抗能力,也可促进植物的生长发育及代谢过程[19]。Éva等[14]发现将克隆的拟南芥AKR4C9 在缺乏AKR的野生型大麦中表达可提高植株对盐胁迫的耐受性。Kanayama等[20]报道,氧化胁迫、寒冷和盐胁迫下蔷薇科桃树(Prunus persica)中克隆到的 PpAKR1 的 mRNA丰度提高,将PpAKR1转入拟南芥提高了植株NADP-依赖型多元醇脱氢酶的活性以及对盐的耐受性。然而在葡萄中对该基因的研究还鲜有报道。因此,本研究利用生物信息学方法,分析鉴定葡萄全基因组序列中的AKR家族基因,从基因组水平分析AKR基因在葡萄染色体分布,基因结构,motif序列,顺式作用原件和进化关系等,利用相关数据库分析了AKR家族成员在葡萄不同组织的表达情况,并分析盐胁迫下葡萄AKR家族基因表达水平随时间变化规律,旨在为进一步研究AKR基因提高葡萄耐盐功能机制提供参考和理论依据。
1 材料与方法
1.1 材料
选取甘肃农业大学生命科学技术学院植物组织培养实验室保存的葡萄试管苗作为供试材料,葡萄品种为品丽珠(Cabernet Franc)。选取长势一致的单茎叶接种于含50 mL GS液体培养基的锥形瓶中培养,培养基pH为5.8-6.0,温度设置为26℃/20℃(光照/黑暗),光周期设置为16 h/8 h(光照/黑暗)。培养25 d后,挑选长势一致的葡萄试管苗进行后续实验。后续将原培养基缓慢倒出,重新倒入50 mL含50 mmol/L NaCl的GS液体培养基继续培养。分别处理3 h、6 h、9 h、12 h和24 h后采集叶片用于后续RNA提取。
1.2 方法
1.2.1 葡萄AKR家族基因成员鉴定及理化性质分析 首先从已知文献中得到22个拟南芥AKR基因ID,接着从拟南芥基因组数据库TAIR(https://www.arabidopsis.org/)下载拟南芥AKR基因编码的氨基酸序列。将获得的氨基酸序列输入到葡萄基因数 据 库 中(https://www.genoscope.cns.fr/externe/GenomeBrowser/Vitis/),查找葡萄AKR基因组成员信息。将所得基因进行DNAMAN比对,去掉冗余序列和片段小于100 bp的序列,共搜索得到13个葡萄AKR基因,分别下载基因全长、CDS序列和氨基酸序列。利用ExPASy数据库(https://web.expasy.org/protparam/)中的ProtParam在线分析工具对蛋白质的理论等电点、氨基酸大小、分子量、不稳定指数、亲水性等基本信息进行分析[21]。
1.2.2 葡萄AKR基因家族进化关系、基因结构、motif序列分析 利用 ClustalX 对拟南芥、苜蓿和葡萄的氨基酸序列进行多重比对[22];使用 MEGA7.0 软件构建AKR基因家族系统发育树,对构建的系统进化树进行自检,bootstrap设定为1 000,处理缺失数据选用Complete deletion,选用P-distance模型对葡萄 AKR基因进行分类[23]。运用GSDS(https://gsds. cbi.pku.edu.cn/)进行基因结构分析[24];利用MEME网站(https://meme-suite.org/tools/meme)进行motif分析[25]。
1.2.3 葡萄AKR家族顺式作用元件及基因芯片表达谱分析 使用PlantCARE软件(https://bioinformatics.psb.ugent.be/webtools/plantcare/html/)对启动子顺式作用元件进行预测分析[26]。使用BAR数据库(https://bar.utoronto.ca/#)检索葡萄不同生长阶段的卷须、叶片、芽、轴、皮肤、花、种子、花粉、果肉、果皮、根等组织器官中AKR基因的表达水平,再对数据进行log2转换,最后使用Omic-share网站(https://www.omicshare.com/tools/Home/Soft/getsoft)制作热图。
1.2.4 葡萄AKR基因家族二级结构及亚细胞定位分析 使用WoLF PSORT网站(http//www.genscript.com/wolf-psort.html)在线进行亚细胞定位。蛋白质二级结构则通过SOPMA在线分析网站(https://www.predictprotein.org/signin#)进行预测。
1.2.5 盐胁迫下葡萄AKR基因家族的实时定量分析 RNA提取采用植物提取试剂盒AG21019(湖南艾科瑞生物工程有限公司,长沙)并按照说明书进行操作。cDNA合成采用TIANGEN公司FastKing gDNA Dispelling RT SuperMix试剂盒,过程参照说明书。引物设计及后期引物合成均由上海生工生物工程技术服务有限公司完成(表1)。荧光定量总体系为20 μL:上游引物1 μL,下游引物1 μL,cDNA 2 μL,ddH2O 6 μL及SYBR Green 10 μL。反应程序设定为:95℃预变性30 s;95℃变性5 s,60℃退火30 s,循环次数为40。

表1 葡萄AKR基因家族实时荧光定量引物Table 1 Real-time fluorescence quantitative primers of AKR gene family in grape
1.2.6 试验数据统计与分析 选择葡萄UBI基因对数据进行归一化处理,基因相对表达量使用2-ΔΔCt法计算。使用SPSS 20计算组间差异的显著水平,Excel软件计算组内误差。使用Omic-share(https://www.omicshare.com/tools/Home/Soft/getsoft)网站绘制热图。
2 结果
2.1 葡萄AKR家族基因理化性质分析
由表2分析可知,经blast比对得到13条葡萄AKR家族基因,分别命名为VvAKR01- VvAKR13。其分布于葡萄19条染色体中的3条:Chrom01分布的基因最多,依次是VvAKR02- VvAKR08;其次是Chrom08,VvAKR09-VvAKR13均分布于此染色体;Chrom18分布1个基因VvAKR01。成员编码氨基酸数在275-430之间,VvAKR06氨基酸序列最长,含430个氨基酸残基,VvAKR11最短,只含有275个氨基酸残基;不稳定指数介于27.01-51.55,VvAKR07基因编码的氨基酸不稳定性最低,为27.01,VvAKR05编码的氨基酸不稳定性最高,为51.55;脂肪指数介于83.57-96.88之间,且该基因家族均具有疏水性;成员分子量在32 083.49-48 655.51之间;理论等电点(pI)分布在5.83-9.01之间。
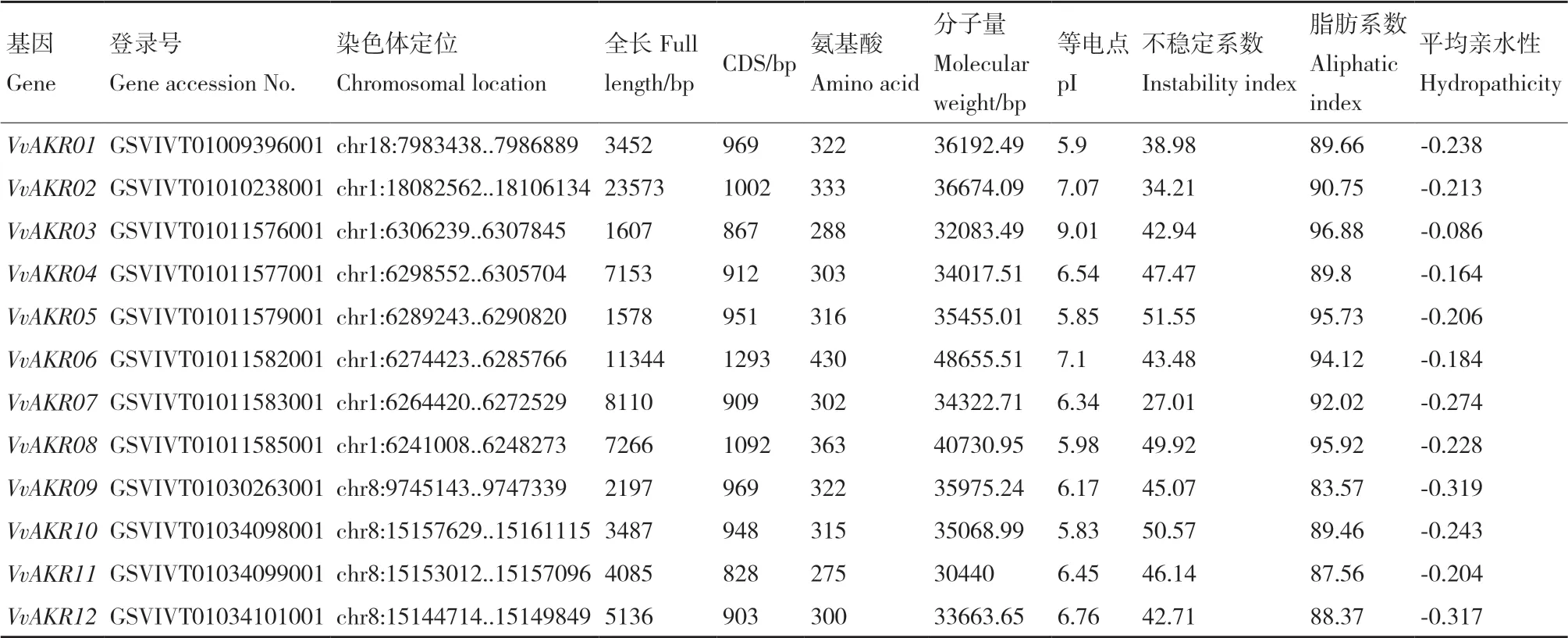
表2 葡萄AKR基因理化性质Table 2 Physical and chemical property of AKR genes in grape
2.2 葡萄AKR家族基因同源性比对及其进化分析
对拟南芥、苜蓿和葡萄AKR家族基因(AtAKR、MtAKR和VvAKR)采用邻接法(Neighbor-Joining,NJ)构建系统发育树,通过构建聚类树将其分为3个亚族(I、II、III),由图1可知,其中I亚族包含的葡萄AKR家族成员最多,有7个,分别为VvAKR01、03、04、05、06、07和08;II亚族包含6个家族成员,分别为VvAKR02、09、10、11、12和13;III亚族没有发现VvAKR基因。
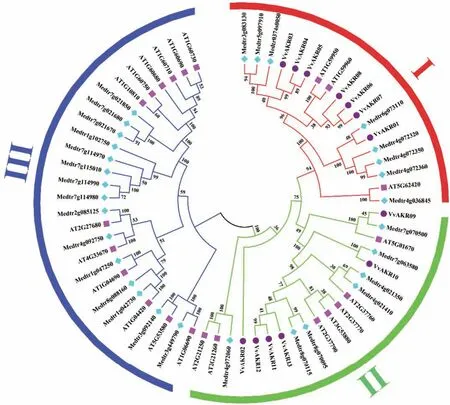
图1 拟南芥、苜蓿和葡萄AKR基因的进化树Fig.1 Phylogenetic tree of AKR genes in Arabidopsis,alfalfa and grape
2.3 葡萄AKR家族基因结构分析
对13个葡萄AKR基因进行结构分析。由图2可知,外显子数在4-13之间,VvAKR01外显子数最少,仅有4个,VvAKR09和VvAKR13外显子数最多,分别有13个;VvAKR02基因最长,可达23 kb左右,VvAKR05基因最短,仅1.5 kb左右;VvAKR01,10,11和12基因均含有外显子,内含子及上下游结构,而VvAKR03和VvAKR04仅具有外显子和内含子,无上游结构和下游结构。
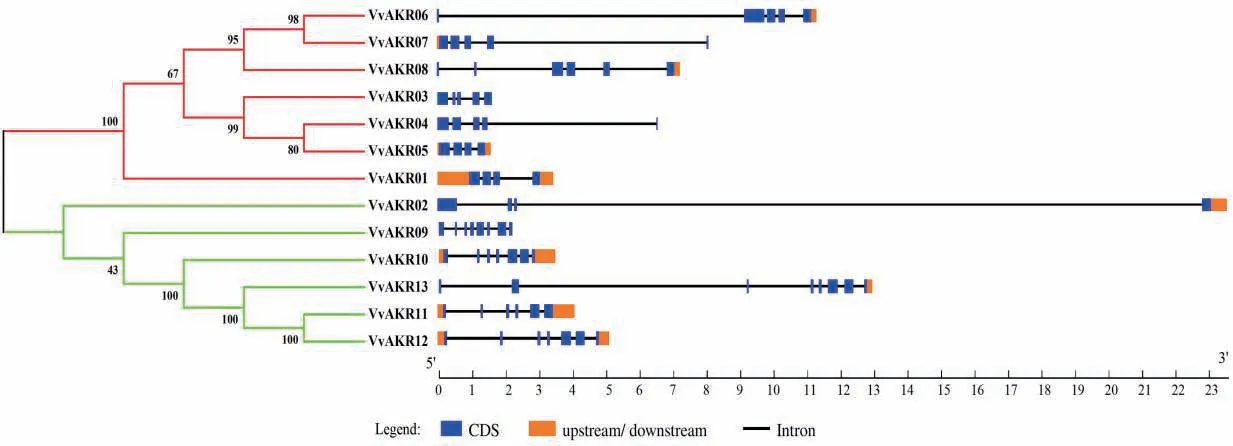
图2 葡萄AKR家族基因结构Fig.2 Structures of AKR gene family in grape
2.4 葡萄AKR家族基因motif序列分析
在葡萄AKR家族基因中共找到了6个motif。由图3可知,13个VvAKR家族成员中,最长氨基酸保守序列有50个氨基酸残基,最短序列有12个,所有成员均含有氨基酸的保守域结构。保守motif分析表明,除VvAKR08外,其他基因均含有motif 1-6;分析发现,VvAKR基因家族成员的C端由motif 3组成;N端由motif 4组成。
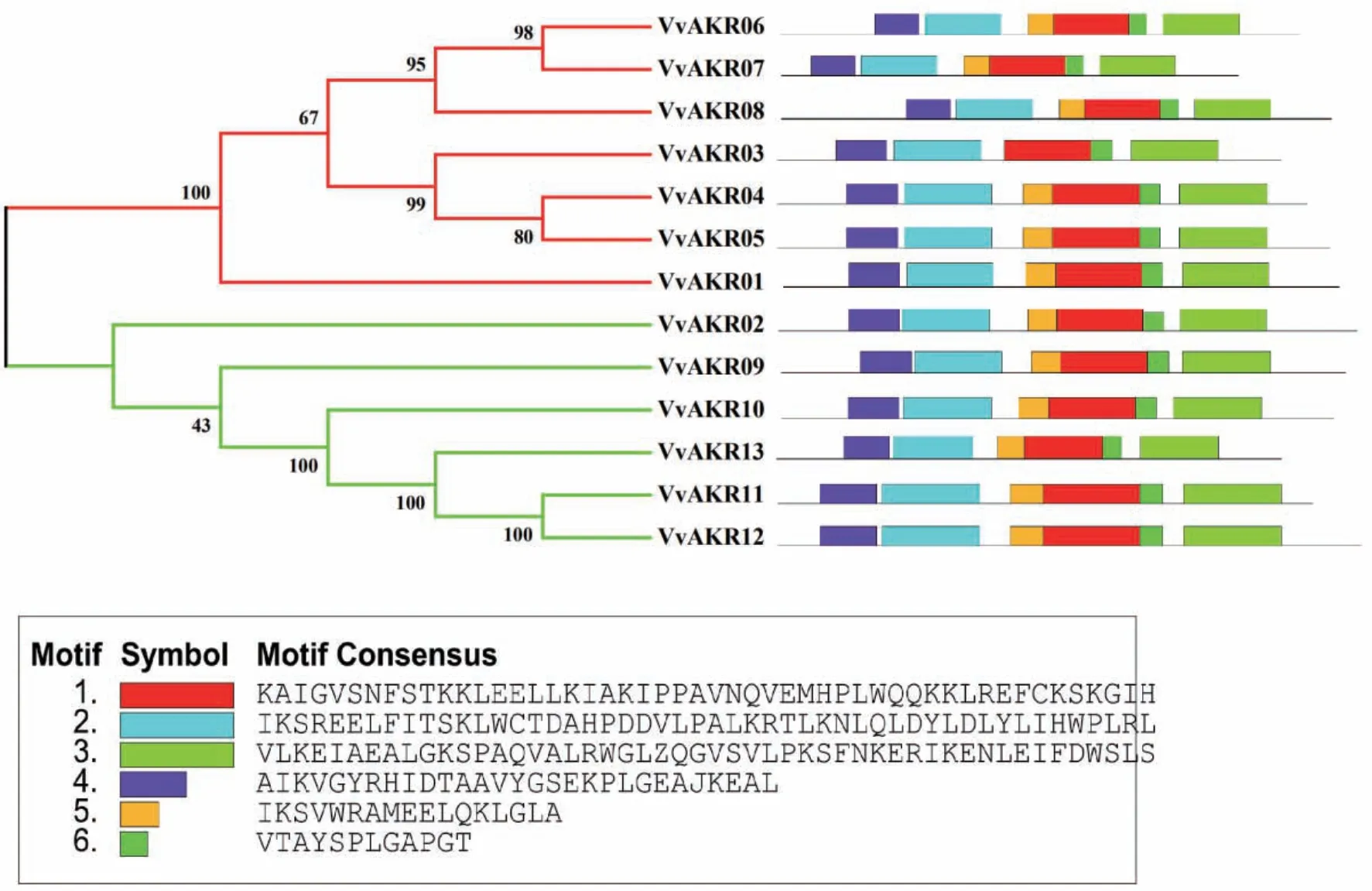
图3 葡萄AKR家族motifs(1-6)分析Fig.3 Motif analysis of AKR gene family in grape
2.5 葡萄AKR基因家族的顺式作用元件分析
启动子顺式作用原件预测结果表明,葡萄AKR家族基因启动子区域鉴定到20个特异响应顺式作用元件,结果如图4所示。该转录因子都存在核心元件TATA-box和增强元件CAAT-box,具备真核生物基因启动子的基本结构特征。除基本的顺式作用元件外,AKR基因还存在其它元件:激素响应元件(TATC-box,CGTCA-motif,TGACG-motif,TGAelement,AuxRR-core,TCA-element和ABRE),胁迫响应元件(MBS,LTR,TC-rich),器官特异性元件(CAT-box)和光调控元件(ACE,G-box,TCTmotif,MRE,I-box,GATA-motif)。激素响应元件具体有茉莉酸甲酯(CGTCA-motif,TGACG-motif),赤霉素(TATC-box),水杨酸(TCA-element),脱落酸(ABRE)和生长素应答顺式元件(AuxRR-core,TGA-element)。
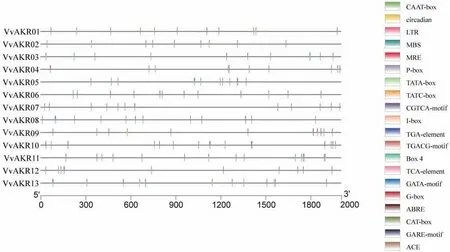
图4 葡萄AKR基因家族的顺式作用元件分析Fig.4 Cis-acting element analysis of AKR gene family in grape
2.6 葡萄AKR家族基因芯片表达谱分析
使用mRNA水平的表达数据制作热图。葡萄AKR家族基因芯片表达模式分析表明,13 个VvAKR基因的表达量在不同植物组织和发育时期有较大的差异(图5)。在几乎所有葡萄组织中,例如卷须,茎、芽、根、花、叶、种子、幼苗等器官,VvAKR02,VvAKR11和VvAKR12都表现出相对较高的表达水平,VvAKR09仅在种子中高表达,VvAKR10、VvAKR13则仅在花粉中高表达。此外,VvAKR03、VvAKR04、VvAKR05、VvAKR06在葡萄组织中表达量均相对较低。
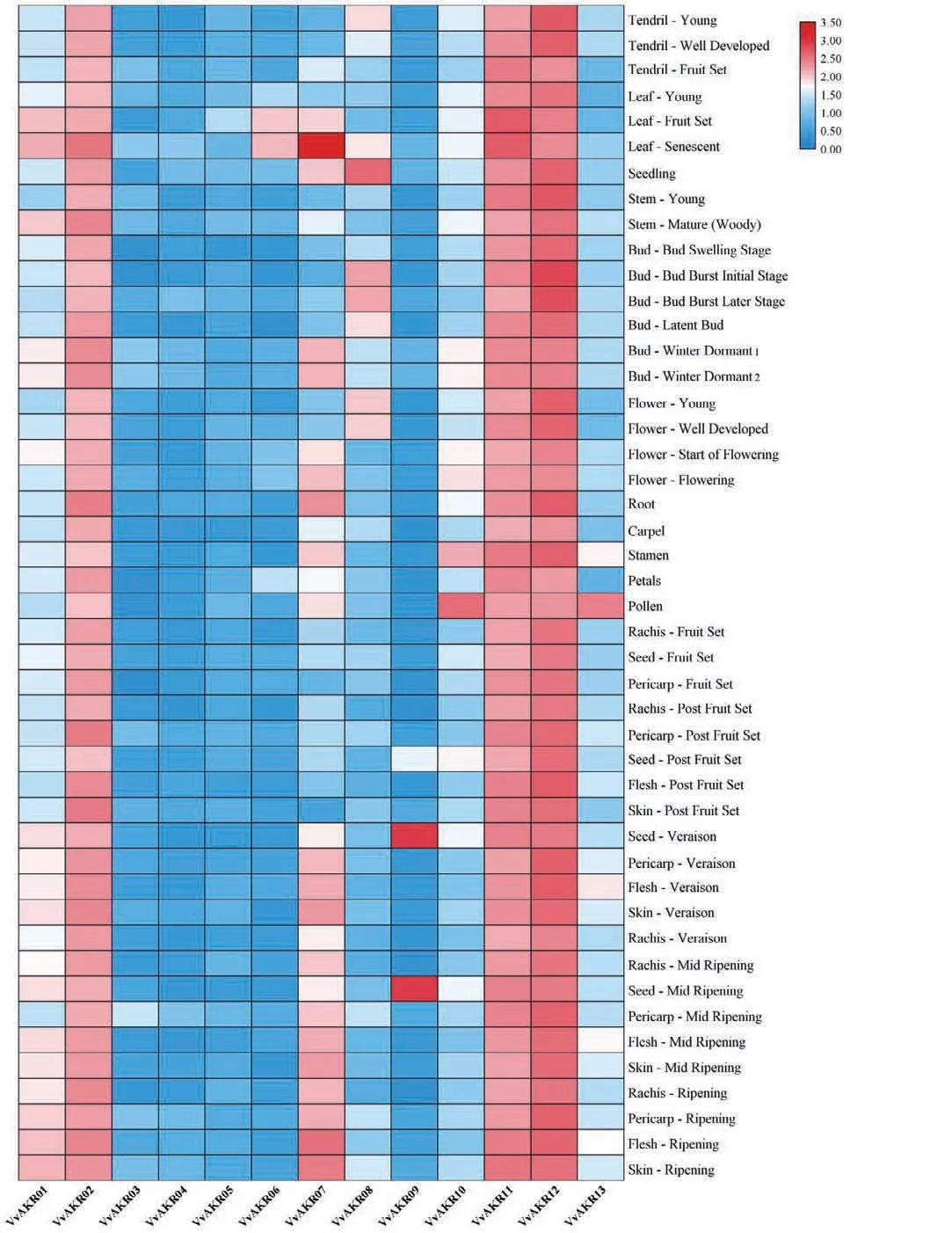
图5 葡萄AKR家族基因表达谱Fig.5 Expression profile of AKR genes in grape
2.7 葡萄AKR家族蛋白质二级结构和亚细胞定位预测
二级结构分析结果如表3所示,葡萄AKR家族基因编码的13个蛋白的二级结构均含有α-螺旋(34.42%-48.91%),β-转角(3.80%-8.39%),扩展链结构(7.38%-18.06%)和不规则卷曲(33.42%-44.19%)。亚细胞定位分析表明,葡萄AKR家族基因主要在细胞核中表达。除VvAKR05外葡萄AKR家族基因在细胞核中均有表达;在叶绿体中表达的 有VvAKR01、02、03、04、05、06、09、10和13;在细胞骨架中表达的有VvAKR01、02、04、06、07、08、09、11和12;在细胞膜中表达的有VvAKR03、06、08、09和12;在线粒体中表达的只有VvAKR03、05和06;在内质网中表达的仅有VvAKR05和VvAKR11;在液泡中表达的只有VvAKR11和VvAKR12。
2.8 盐胁迫下葡萄AKR家族基因表达特征分析
为探究盐胁迫下葡萄AKR家族基因表达水平随时间变化的规律,以未用盐处理的葡萄叶片为对照,采集盐处理3 h,6 h,9 h,12 h和24 h五个不同时长的葡萄叶片进行qRT-PCR试验。结果表明(图 6),随着盐胁迫时间延长,VvAKR02,03,05,06,07,10,11,12和13表达水平在不同时间点存在不同程度的提升,但变化趋势不尽相同。其中相对表达量变化趋势表现为先升高后降低最后回升的基因为VvAKR02,05,07,11,12和13,在3 h或6 h表达量上调,随后降低,再于12 h和24 h逐步回升,且这6个基因在盐胁迫后各个时间点的表达水平都较对照表达上调。而VvAKR03,06和10虽然在某一个时间点较对照上调表达,但盐胁迫后总体表达水平下调。VvAKR01,04,08和09盐胁迫处理后不同时间点表达水平显著降低。
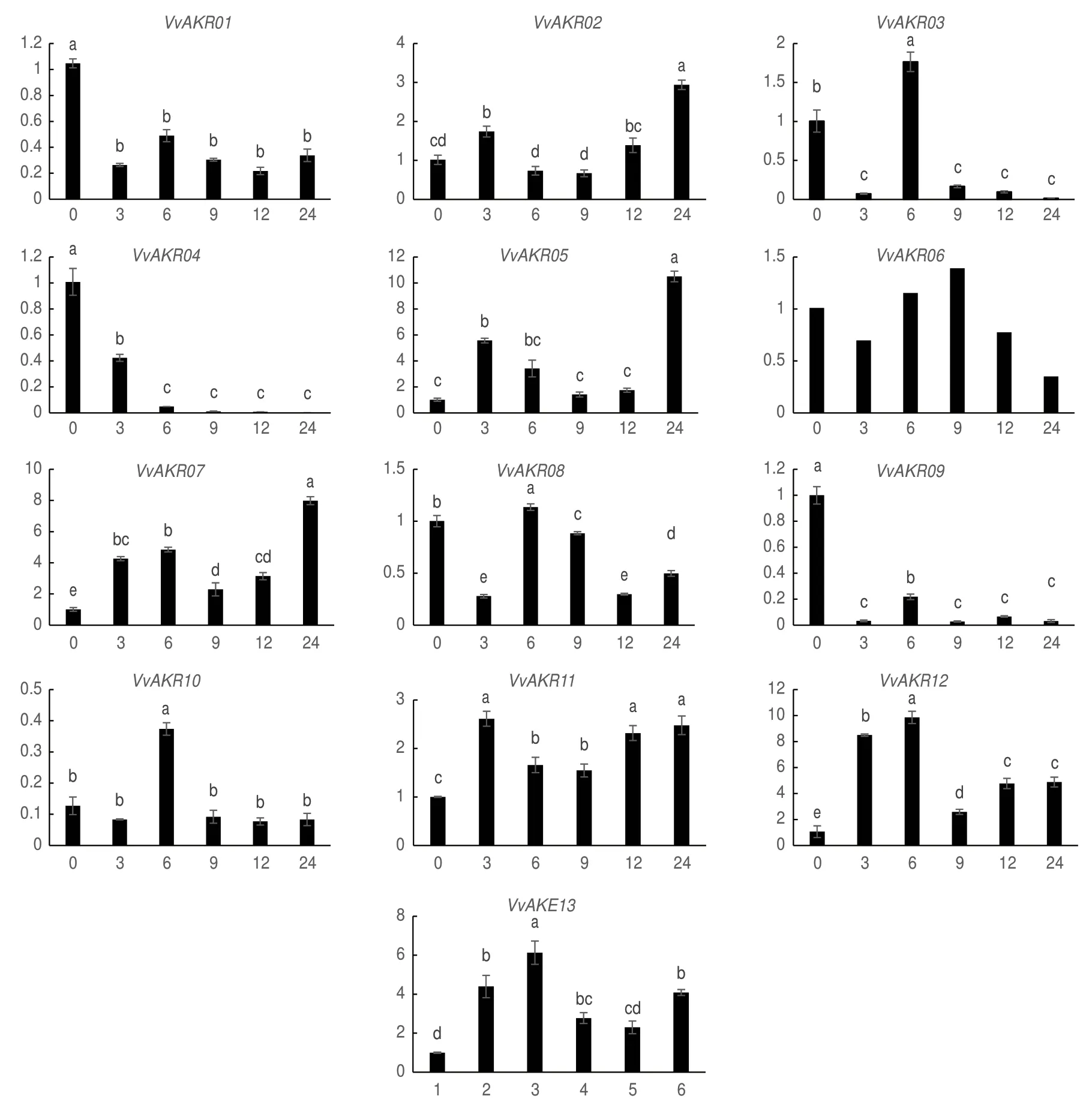
图6 葡萄AKR家族基因响应盐胁迫表达模式分析Fig.6 Expression pattern analysis of grape AKR family genes in response to salt stress
3 讨论
本文以拟南芥已知AKR基因序列为基础,同源比对出13个葡萄AKR家族基因成员。将葡萄与拟南芥、苜蓿的AKR家族基因共同构建进化树后,可将所有基因根据进化关系分为3个亚族(I、II、III)。I亚族包含的葡萄AKR家族成员最多,III亚族中没有葡萄AKR基因分布,II亚族中包含VvAKR02,VvAKR09,VvAKR10,VvAKR11,VvAKR12和VvAKR13六个成员,其中VvAKR02,VvAKR11,VvAKR12和VvAKR13亲缘关系相近且在响应盐胁迫时的荧光定量结果中都呈现上调表达的情况且分支中成员与植物响应盐胁迫相关,推测该进化树对VvAKR功能分类有一定参考作用。同一亚族氨基酸序列上外显子的分布位置差异性较大,且序列长度无明显规律。葡萄AKR家族基因中共找到了6个motif,除VvAKR08外,其他基因均含有motif 1-6,表明葡萄AKR家族基因间保守程度高。在蛋白质编码水平上外显子数量、长度及分布位置无规律,在蛋白水平上成员间保守程度高,AKR家族不同成员采取不同的转录翻译过程最终形成具有相近功能的成熟蛋白质,说明此家族基因上的保守程度与蛋白上的保守程度并不一致。亚细胞定位和二级结构分析表明,葡萄AKR家族基因成员多在细胞核中表达,因此推测该家族成员对葡萄细胞核内的信号转导具有重要功能,此研究结果与甘蓝及苜蓿AKR家族的聚类分析结果相似[16,27]。
生物信息学分析表明,AKR家族成员均具有疏水性,不稳定指数均较低。不同基因编码的氨基酸数各不相同,平均含有325个氨基酸,平均蛋白质量为36 kD,这与先前发表的大多数AKRs含有约320个氨基酸,蛋白大小为33-37 kD的观点一致[28],说明鉴定结果可靠。基因芯片分析表明,13个VvAKR基因的表达量在不同植物组织和发育时期各不相同,这种在转录表达上的差异可以归因于启动子中顺式元件的种类和数量。VvAKR02,VvAKR11和VvAKR12几乎在所有葡萄组织中均表现出相对较高的表达水平,包括卷须、茎、芽、根、花、叶、种子、幼苗等,推测这几个基因对葡萄的生长发育起着主要调控作用[18]。VvAKR09在种子中高表达,而VvAKR10,VvAKR13仅在花粉中高表达,说明不同的AKR家族成员具有组织特异性,此研究结果与苜蓿AKR基因家族的研究结果相似[27]。
顺势作用元件结果表明,VvAKR顺式作用元件中大量存在生长素,脱落酸,低温和干旱等响应元件,表明其在植物生长和对环境胁迫的响应中具有潜在的功能。由前人对AKR家族进行全基因组分析和功能分析可知,当植物受到干旱[29-30]、高温和低温[2,29]、高盐[29]、紫外线[30]、过氧化氢[31]、百草枯[31]。病原菌毒素[32-33]等生物和非生物胁迫时,植物体内的AKR被诱导表达,进而参与一系列的代谢过程。本文在分析盐胁迫处理后葡萄AKR基因表达模式后可知,各基因在胁迫后不同时间点的表达存在明显差异。VvAKR02、VvAKR05、VvAKR07、VvAKR11、VvAKR12和VvAKR13表 达 显 著 上 调,VvAKR03、VvAKR06和VvAKR10存在不同程度的上调,VvAKR01、VvAKR04、VvAKR08和VvAKR09的表达量表现为下调。这说明葡萄AKR家族基因参与葡萄盐胁迫响应,且不同基因对盐胁迫的响应模式不同,其中VvAKR02、VvAKR05、VvAKR07、VvAKR11、VvAKR12和VvAKR13可能与葡萄耐盐相关。此研究结果与洋地黄中克隆得到的AKR基因表达情况 相似[29]。
4 结论
本文从葡萄中鉴定得到13个AKR家族基因,该家族基因在葡萄不同发育时期和不同组织器官表达存在差异;上游存在丰富的顺式作用元件,除核心元件和增强元件之外,同时还存在各种激素及胁迫响应元件,器官特异性元件和光调控元件;在盐胁迫下,该家族中VvAKR02,VvAKR05,VvAKR07,VvAKR11,VvAKR12和VvAKR13可受盐胁迫诱导表达上调。通过对葡萄AKR基因结构,进化关系,功能的研究分析,有助于了解葡萄AKR基因家族成员的结构与功能的关系,并且为揭示AKR基因响应盐胁迫的调控机制和信号转导网络的研究和挖掘葡萄耐盐基因奠定一定的理论基础。
