石栎属物种果实类型及其系统演化研究进展
2021-09-11覃元艺
覃元艺,陈 曦,2,3
(1.云南大学 生态与环境学院,云南 昆明 650091;2.云南大学 云南省植物繁殖适应与进化生态学实验室, 云南 昆明 650091;3.云南大学 云南省高原山地生态与退化环境修复重点实验室,云南 昆明 650091)
壳斗科(Fagaceae)包括12个属 1 000 余种,是北半球热带、亚热带及温带森林的主要建群树种,也是最重要的木本植物大科之一[1-2].石栎属(LithocarpusBlume)是壳斗科内物种多样性极为丰富的重要大属,其320多个种广布于东亚、东南亚及喜马拉雅南麓中低海拔河谷[1-4].我国和东南亚是石栎植物多样性的2个重要分布中心.石栎不仅是热带亚热带常绿阔叶林林冠层的主要树种和建群种[5-7],其种子也是这些地区众多动物类群的食物和营养来源[7-10].因此石栎对维持这些森林系统的碳汇平衡和生态多样性起着重要作用.
对比同科的栎属(QuercusL.),石栎属获得的研究关注相对较少.这首先是因为栎属植物在北半球广泛分布,而很多西方研究者并不熟悉分布于东亚的石栎.另外,栎属植物是重要的经济树种,而此前石栎属植物由于木质较差被认为经济价值不高.但最近几十年来,石栎属植物的经济开发价值受到关注,如食用[11-12]、药用[13-18]、造林绿化[19-23]等多个方面.可见,除了具有重要生态价值之外,石栎也兼顾多项潜在的经济价值.在经济利益的驱动下,人们开始从自然种群中采集和移栽石栎物种,但这种缺乏科学管理的采集造成了石栎种群的过度开发和森林群落生物多样性的丧失,并在未来可能带来生物入侵和污染问题,甚至影响全球气候变化[6, 24-26].
因此,为石栎资源的科学合理开发提供理论依据,必须首先深化石栎的基础研究.在众多方向的基础研究之中,以石栎果实形态和类型为主线拓展出的研究极具特色.由于石栎物种果实形态变异丰富(见图1),图中黑色直线代表 1 cm 长度.因部分石栎物种在果实成熟时壳斗与果实分离,因此壳斗结构缺失,未能标注.因此其果实形态学成为理解其物种多样性和进化的关键.通过将石栎的果实形态和果实类型研究与植物生理、形态发育、生物地理、种子散播和物种更新以及分子系统发育等方面贯通融合,得以对石栎的分布、物种进化和共存、物种和形态多样性、物化特性与生活史对策等一系列问题有了初步的理解.因此本文归纳了最新的石栎属植物的果实形态多样性与物种分化研究,探讨了石栎果实形态和类型与其分布、物种多样性、物种更新和分子系统发育之间的重要联系和科学意义.同时探讨石栎研究过程中存在的问题与重要的研究空白,对未来石栎基础研究方向提出建议.
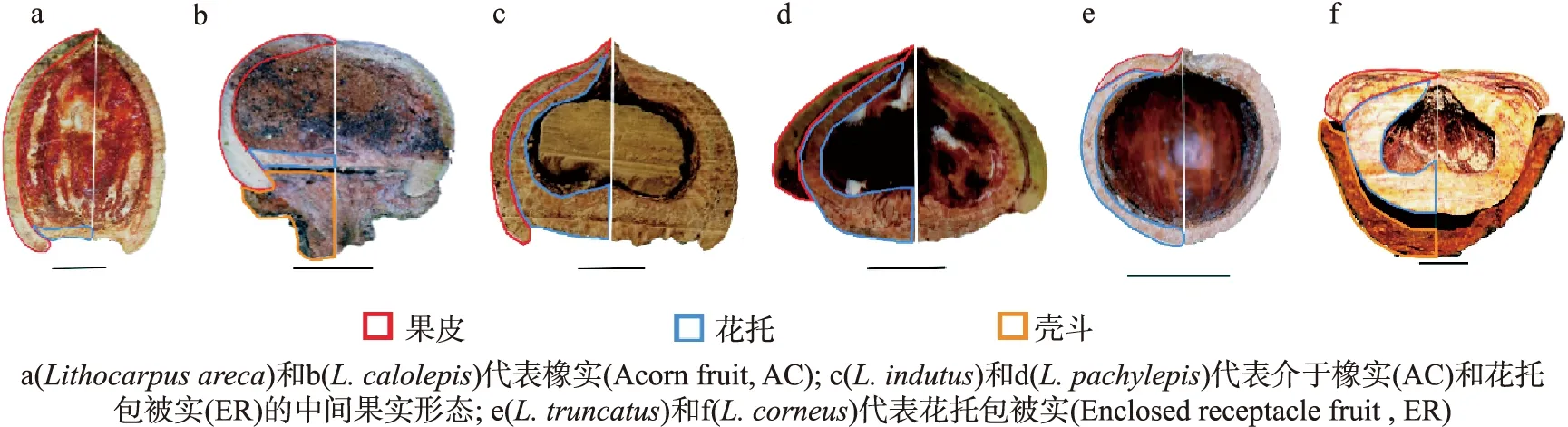
图1 6个石栎物种果实的纵截面形态结构
1 石栎果实形态多样性与物种分化研究
1.1 石栎属2种主要果实类型的形态和物化权衡
由于石栎物种的无性结构(如枝、叶等)分化不明显[28],因此形态变异丰富的果实和壳斗被作为物种鉴定的重要证据广泛应用于传统分类学中[29-31].石栎物种单个的坚果果实被杯状壳斗结构所包被[32](见图1b、1f).壳斗是壳斗科植物的重要特征,是无性枝融合后形成的扁平状或碗状结构[33].壳斗内是单个的石栎坚果,其果壳由果皮和花托组成[27](见图1).
早在20世纪中期,法国科学家Camus就注意到石栎物种的果实形态变异显著(图1).2001年,Cannon和Manos首次尝试分析石栎果实的形态多样性,用elliptic fourier analysis (EFA)量化21种热带石栎物种果实的花托和果皮形态差异,发现这21个物种的果实形态分化可以被归纳为2种果实类型:橡实(acorn fruit,后简称AC)和花托包被实(enclosed receptacle fruit,后简称ER)[28](图2).图中黑色直线代表 1 cm 长度.橡实(AC)与栎属植物果实(俗称麻栗果)形态十分类似,单薄的果皮是包被其种子的主要结构,花托存在于种子基部,通常为扁平凹形(图2a、2c);而花托包被实(ER)的果皮急剧缩短仅覆盖于种子顶部,花托却急剧延伸加厚成为包裹种子的主要结构(图2d、2f).这项研究为石栎的果实形态和果实类型研究奠定了重要基础,但其局限性体现在(1)仅包括热带的21个石栎物种,缺乏对亚热带石栎和丰富石栎物种多样性(320多个种)的代表;(2)该研究通过肉眼判断物种果实类型的方法缺乏数据支撑.

图2 形态学研究对橡实(AC)(Lithocarpus kawakamii,a~c)和花托包被实(ER)(L. corneus,d~f)的果实结构理解的变化发展
为更好地体现石栎属层面的果实形态分化,陈曦等针对广泛分布于中国和东南亚的98个石栎物种(代表了石栎属1/3的物种多样性),从物种果实纵截面图像利用Pappus-Guldinus定理重建果实3D形态,量化多项果实形态特性(包括果实和种子体积、外果皮和花托对种子的包被程度等),并进行了种内和种间的果实形态变异分析.研究结果表明,石栎物种内的形态分化较小,石栎属果实多态性主要由种间形态分化造成.且该研究通过对比外果皮和花托对种子的包被程度进行果实类型划分,相较于Cannon和Manos用肉眼进行果实类型划分的方法更加客观准确.该研究结果支持Cannon 和Manos提出的2种主要果实类型:橡实(AC)和花托包被实(ER)代表了石栎属层面的果实形态多样性,但橡实(AC)是石栎属更常见和普遍的果实类型(只有不到1/3的物种生产花托包被实(ER)).该研究还证明了2种果实类型存在形态和物理防御分化:花托包被实(ER)和橡实(AC)的主要物理防御结构分别是花托和果皮(图2),花托包被实(ER)的果实和种子较大,且果实物理防御水平显著高于橡实(AC)[7].
为理解2种果实类型其物理防御权衡的形成过程和发育差异,陈曦等分析了代表2种果实类型的20个石栎物种在发育过程中的的多项果实形态动态特征:种子大小、外中果皮联合体体积(由于2个组织结合紧密,故而作为一个整体进行分析)、内果皮体积和花托体积的增长变化,再现了2种果实类型从果实发育初期到成熟的动态发展过程[27].这项研究的重要性首先在于对石栎果实结构的修正认定:此前石栎果实形态和果实类型研究[7, 28, 35]中的被认为包被橡实(AC)的主要结构——“外果皮”(图2b、2e),实为外、中、内果皮的总和,也就是果皮结构(图2c、2f) (为避免概念混乱,前研究中出现“外果皮”结构在本文中均纠正为“果皮”)[27].更重要的是,该研究发现2种果实类型在果实发育早期的形态结构和组织分化顺序:外中果皮联合体首先分化出现,内果皮和花托后分化出现,最后才是种子发育;但2种果实类型在果实发育后期的组织分化差异造就了它们在成熟时的果实形态分化和物理防御权衡:果实发育后期,橡实(AC)的内果皮发育显著,而花托包被实(ER)的花托发育显著,于是形成了单薄的果皮代表了橡实(AC)的弱物理防御,而木质化加厚的花托是花托包被实(ER)的强物理防御[27].此外,该研究也支持Brett(1964)提出的壳斗功能假说:壳斗为处于发育初期阶段的果实,尤其是橡实(AC)提供机械保护.这项修订明确了石栎果实结构和2种果实类型的形态差异的发育学原因,不仅为理解2种果实类型功能进化机制奠定了重要基础[36],也为研究其他果实发育研究提供了模型范本.
除了果实形态和物理防御权衡,这2种果实类型也存在种子化学成分权衡:花托包被实(ER)种子的营养成分(粗蛋白质、脂肪、单糖和总非结构型碳水化合物)较高,而橡实(AC)种子中代表化学防御的拒食素(中性洗涤纤维、酸性洗涤纤维、纤维素、半纤维素、木质素、总单宁,缩合单宁和水解单宁)水平更高[35].出乎意料的是,橡实(AC)中主要的拒食素成分并非为人熟知的单宁[37],而是纤维素[35].部分哺乳动物的唾液中存在有单宁绑定唾液酶(tannin binding silavary proteins, TBSPs),这种酶可以抑制单宁的毒性,从而抑制单宁作为化学防御的角色.因此,研究者们猜想可能石栎种子的捕食者唾液中存在单宁绑定唾液酶,因此石栎种子转而投资生产无法被动物分解的纤维素,来抑制动物对能量和营养的获取,成为更有效的化学防御[35].
综合以上石栎果实类型形态分化和理化特性研究,发现石栎2种果实类型存在一系列的形态和物化权衡:橡实(AC)的果实和种子较小,单薄的果皮是其种子的主要防御结构,因此物理防御弱,但种子内的化学防御水平高;而花托包被实(ER)的种子和果实更大,种子富含营养,而木质化加厚的花托是包裹种子的主要结构,代表了强物理防御[7, 27, 28, 35](表1).但是,需要注意的是,观察到有一些石栎物种代表了不便于划分、介于2种果实类型间的中间形态(图1c、1d):它们的花托急剧延长加厚,几乎覆盖了整个种子,但同时果皮也没有缩短,而是覆盖在花托外部,和花托共同保护种子.陈曦等也指出虽然可以根据花托和果皮对种子的绝对包被程度将这些物种划分成橡实(AC)或者花托包被实(ER)[7],但是它们在形态上展现出来的特殊性对石栎果实形态和进化研究有重要意义.通过对具有中间果实形态的物种进行详细的果实理化特性分析,并与2种果实类型进行对比,是回答中间果实类型与2种果实类型关系和差异的重要环节,也是理解石栎果实类型进化的关键.

表1 石栎属两种果实类型形态和理化特性对比
1.2 2种果实类型的地理分布特性
为了探究石栎果实形态多样性与其地理分布的内在联系,陈曦等人调查了93个石栎物种的分布情况,发现代表2种果实类型的石栎物种的经纬度分布有很大的重叠性,共享中国和东南亚2个主要分布中心[7].从群落层面上看,2种果实类型的石栎物种也常常共生[35, 38-40],甚至具有相似的优势度[7].广义线性分析显示石栎种子随海拔升高和纬度降低而变大,且石栎果实物理防御均随海拔和纬度升高而增大[7].作者们推测种子变大可能是为了补偿由于海拔升高而产生的环境胁迫(如低温、短光和周期等)的适应性结果,且大种子需要更多的物理防御投资抑制种子捕食;而热带地区石栎种子更大则可能是热带环境因子较亚热带地区更适于生产大种子、或者热带是更多的种子散播媒介等原因综合影响下的结果[41].
目前,石栎属的分布概况和2种主要果实类型的分布格局基本明晰,不难看出2种果实类型的果实形态分化和物化权衡并非由于其地理分布分化造成,而更可能是和与其果实相互作用的动物的长期共同进化的过程中,产生的生态位分化的结果.将在第2部分对石栎果实和动物的相互关系做详细的论述.除此之外,气候因子决定了物种的适宜生境,对物种的分布格局产生直接影响[42-45].并且,有证据显示地理因子并不能完全解释物种特征和种间分化[46].因此,在未来分析温度、降水等重要气候因子对石栎物种和2种果实类型分布的影响和规律,是进一步理解石栎物种多样性和果实类型分化的重要步骤.
2 石栎的种子散播和物种更新研究
2.1 石栎种子的捕食散播研究
石栎种子是东亚热带亚热带常绿阔叶林内众多动物类群的重要食物和营养来源[7-10].其中,昆虫和脊椎动物是2类最主要的种子捕食者[32],分别代表着与石栎种子的2种截然不同的相互关系:抑制种子散播的拮抗关系和有助于种子散播的互利共生关系[32, 47-48].昆虫与石栎种子的拮抗作用体现在昆虫消费种子,获得了能量和繁衍后代的机会,但种子产量被严重削弱,种子散播受限,从而对石栎种群重建产生不利影响[48].而具有分散囤积行为的脊椎动物则和石栎植物形成了互利共生的关系,它们同时扮演着捕食者和散播者的双重角色:它们在消费种子获取食物和能量的同时,也搬运种子使其远离母树,将种子埋藏在有利于种子萌发和幼苗建立的土壤浅表,扩大了石栎的分布范围和生存空间,以及繁衍后代的机率[8-9,49-50].
目前,针对石栎果实与动物相互关系对研究十分有限,只了解到一些昆虫和脊椎动物捕食者的片段信息:如象甲、雪片象、瘿蜂可能是某些石栎物种种子的昆虫捕食者[51];而目前已知的石栎的脊椎动物捕食散播者多为啮齿类动物,如白腹鼠属(Niviventercoxingi)、鼠属(Rattus)、巨鼠属(Berylmys)、姬鼠属(Apodermus)、小泡鼠属(Leopoldamys)、巢鼠属(Micromys)、社鼠(Niviventerconfucianus)和针毛鼠(Niviventerfulvescens)等[9, 49-50, 52],而虽然观察到更大型的哺乳动物如野猪也捕食石栎果实[7],但它们是否对种子有散播还有待考证.果实的物理防御和化学成分特性和其动物捕食者密切相关[32],因此,共生且物化特性相异的2种果实类型的石栎是种子捕食散播研究的理想模型.而且由于代表这2种果实类型的石栎物种往往共生[7],因此我们推测具有形态和物化权衡的2种果实类型可能是针对昆虫和脊椎动物不同捕食压力下,进化形成的2种不同的种子保护、散播和种群更新的适应性对策:花托包被实(ER)的物理防御强,限制了昆虫对其捕食,同时促进了脊椎动物对果实的散播,其种子大且营养丰富,有利于种子萌发和幼苗建立,但由于单个果实生殖能量投入高,限制了果实总产量和建植的种苗数量;而机械防御弱的橡实(AC)被更多类型的昆虫捕食,也被脊椎动物更多地原地消费,但其单个果实的生殖能量投入小,果实总产量高,因此足够数量的果实可以躲避捕食,萌发定植.因此,全面调查2种石栎果实类型的昆虫捕食者和脊椎动物捕食散播者,同时结合石栎果实形态和理化特性,分析2种果实类型与昆虫和脊椎动物的相互作用差异,是验证2种果实类型生活史策略的假说和解析石栎2种果实类型进化动力和其共生机制的关键步骤.
2.2 石栎种子萌发特性及幼苗生长研究
在种子散播后,种子的萌发和定植是影响石栎种群重建和分布的另一重要步骤.虽然石栎属植物可以大量结实,但在成年石栎的森林中却很难发现石栎幼苗,这可能与石栎种子的低萌发率相关[26, 53-56].种子的萌发率同时受到非生物因子和生物因子两方面的影响.影响石栎种子萌发的重要的非生物因子包括温度、湿度和植物激素等,如研究发现石栎种子在 25 ℃ 条件下萌发率最高[53-54];且4%的硝酸钾、浓硫酸,赤霉素等都能促进石栎种子的萌发[26, 54-55].生物因子如昆虫,他们捕食石栎种子,除了降低种子产量并限制种子散播外,昆虫携带的真菌、细菌和病毒也会造成种子感染甚至死亡[32,48,56-57].种子通过各种化学和物理防御来降低被捕食和感染的风险[58-59],但这些防御也可能成为种子萌发的阻碍.例如,不同的研究都发现剥去果壳可以促进石栎种子的萌发[26,53-54].以上研究让我们对影响石栎种子萌发的因素有了一些基础的了解,但是,我们对石栎属2种果实类型的种子在萌发特性上的差异却不甚了解:较大且富含营养的花托包被实(ER)种子是否比较小的橡实(AC)种子萌发率更高;或者花托包被实(ER)的木质化加厚的果壳是其萌发的阻碍,而橡实(AC)果壳脆弱因此萌发率更高?2种果实类型的种子萌发是否需要不同的适宜环境?解析2种果实类型萌发特性差异与其果实形态和物化权衡之间的联系是我们理解石栎物种和果实类型分化的关键,也是理解2种类型石栎物种的共存和生活史策略的重要步骤.
而目前针对石栎幼苗生长的研究,主要集中于提高苗木成活率的育苗方法:包括(1)适于石栎幼苗生长的基质配方:如研究发现最适于东南石栎(L.harlandii)幼苗生长的基质配方为泥炭土∶珍珠岩∶锯屑为 3∶1∶1[60];(2)肥料对石栎幼苗的生长和生物量表现的影响:如耳叶石栎(L.elegans)的幼苗在一公顷 100 kg N尿素肥料剂量下生长和生物量表现最好[26];(3)通过控制蒸腾作用来提高石栎苗木成活率:如摘除榄叶石栎(L.oleaefolia)苗木上部分较大叶片,减少叶面蒸腾蒸发的水分可提高其苗木成活率等[61].这些研究为石栎的造林和引种栽培奠定了重要理论指导,对石栎的科学合理开采提供了重要的科学理论基础.但是,我们对自然生态系统中石栎的幼苗生长和生存却鲜有了解.石栎幼苗的形态和成树形态差异明显,这造成了野外石栎幼苗辨认难以及不同石栎物种幼苗鉴定困难的情况.因此,我们建议在本地寻求专业支持,如林业局或者熟悉植物的本地山民协助进行石栎幼苗识别和物种鉴定,同时对幼苗生长发育情况进行年间追踪观测,深入了解石栎群落结构和种群重建过程,分析探讨石栎多样性和分布形成原因.
3 石栎属的分子系统发育及两种果实类型的进化关系
随着分子生物学的迅猛发展,利用基因数据进行分子系统发育研究成为目前研究的热点领域之一.Cannon和Manos在2003年利用叶绿体基因(cpDNA)和内转录间隔区(nrITS)构建了少量婆罗洲热带及亚洲大陆石栎物种的系统发育树,并提出石栎属起源于东南亚热带,随后向亚洲方向迁徙的理论[62].他们认为向亚洲迁徙的过程中,一次向西扩散到欧洲(只有化石记录,没有现存物种),另一次向东扩散到北美西部,有一个现存的物种[62].而后来的研究发现,北美西部的现存物种并非石栎物种,而应认定为新属Notholithocarpus[63-64].由于Cannon和Manos的石栎理论还有待进一步的验证,Yang等于2018年通过内转录间隔区(nrITS)和叶绿体基因的atpB-rbcL序列,构建了84个石栎物种的3个系统发育树(包括联合内转录间隔区(nrITS)和叶绿体基因的atpB-rbcL序列构建的一个系统发育树以及单独由内转录间隔区(nrITS)或叶绿体基因的atpB-rbcL序列构建的2个系统发育树).相对于Cannon和Manos在2003年构建的热带及少量亚洲大陆石栎物种的系统发育树,该研究囊括了1/3的石栎物种多样性并代表了石栎的主要分布地区(东亚、东南亚),成为目前已发表的最完善的石栎属系统发育树.另外,他们否定了Cannon和Manos的石栎热带起源的假说[62],指出石栎属起源于中国亚热带,于中新世早期向东南亚迁移,随后在大巽他群岛进行原位多样化,造成了目前东南亚石栎物种丰富的现状[65].
另外,Cannon和Manos在2001年通过内转录间隔区(nrITS)构建了21个热带石栎物种系统发育树,并和果实形态研究相结合,初步揭示了2种果实类型的系统进化关系:花托包被实(ER)至少在不同进化枝中独立进化了2次,可能是进化中后出现的果实类型[28].他们还发现果实类型的形态变化和基因变异没有必然的对应关系:橡实(AC)物种的种内和种间形态变异均小于其基因的变异[66-67],而花托包被实(ER)形态的变异却大于其基因的变异[28].这说明花托包被实形态学方面临更大的选择压力,同样暗示2种果实类型可能是对应不同捕食压力下分化的产物.但这项研究使用的基因数量(一个内转录间隔区(nrITS))和物种数量少(21个物种),且其果实类型划分缺乏数据支持.
因此,对Yang等[65]发表的系统发育树上的49个石栎物种的果实类型进行精确划分,并将其标注到系统发育树上(图3).结果显示,49个物种中,32个物种属于橡实(AC)类型,17个物种属于花托包被实(ER)类型.且花托包被实(ER)的在不同的进化枝之中反复多次出现,说明其不是进化选择的偶然性,在进化上存在一定的优势.但这些分析还不足以完全理解2种果实类型之间的进化关系.首先,Yang等建立的系统发育树所使用的基因数量少(只有内转录间隔区(nrITS)和叶绿体基因的atpB-rbcL序列),因此系统树的稳定性较弱,种间关系支持力度不足.因此,增加基因数量来建构完善且稳定的石栎系统发育关系,并完善石栎物种果实形态多样性研究十分必要.更重要的另一个方面是,目前单纯的分子系统发育研究存在很多缺陷,如基因组数据集的分析技术存在偏差[68],或者单纯依赖分子数据可能会误判物种的系统发育关系等[69-70].形态学证据是完善分子系统发育和物种间关系的重要环节.因此,将石栎物种果实形态学特征(如果实类型、果实大小、叶片大小等)与其分子发育系统结合,对其进行祖先形状重建和系统信号发育研究可以解决分子数据难以处理的物种系统位置、提高分子进化树的准确性[71].另一方面,通过系统发育树追踪果实特征的进化可以初步探究果实类型变化的分子基础,了解石栎果实形态多样性与其物种进化的相互关联,为探究石栎属关键种系统位置和理解2种果实类型进化关系提供佐证.这不仅可以帮助我们进一步验证石栎的起源(热带还是亚热带),也是揭示2种果实类型进化关系的关键.

图3 结合分子系统发育树的石栎物种果实类型演化关系图
4 总结与展望
石栎物种果实形态多态性与其物种进化和多样性形成密切相关.2种主要的果实类型:橡实(AC)和花托包被实(ER)代表了石栎属物种间丰富的果实形态变异.这2种果实类型存在果实形态和物化特性的权衡:橡实(AC)的果实和种子较小,种子内的高拒食素成分是强化学防御,包被种子的单薄果皮代表了弱物理防御;而花托包被实(ER)的种子和果实更大,种子营养成分高,木质化加厚的花托保护种子代表了强物理防御[7, 28, 35].果实发育学证据显示这2种果实类型的形态权衡主要由花托和果皮在果实发育后期相异的分化速率造成[27].地理分布研究显示,这2种果实类型的石栎物种同时共享中国南部和东南亚2个分布中心,且往往在不同地区的不同森林类型中共生[7].
基于以上研究基础,课题组推测2种果实类型在形态和物化特性上的分化如果不是地理分布差异造成,那它们很可能存在生态位的分化,代表了不同捕食压力下长期进化出现的2种适应性对策:花托包被实(ER)物理防御强,既限制了昆虫对其种子的捕食,也促进了脊椎动物对种子的散播,同时高营养的种子利于种子萌发和定植;而机械防御弱的橡实(AC),不仅被更多类型的昆虫所捕食,也被脊椎动物进行大量原地消费,但其果实小,果实总产量大,最终足够数量的果实能够躲避捕食,完成种子萌发定植.因此,未来急需开展石栎果实形态和理化特性与昆虫和脊椎动物的相互作用研究,来解析石栎2种果实类型的进化动力和其共生机制.同样重要的是,分析对比2种果实类型的石栎种子的萌发特性和幼苗生长情况,是回答2种果实类型的种群更新差异、生态位分化和生活史策略的另一个关键.
通过将果实形态研究与石栎分子系统发育结合,初步揭示了花托包被实(ER)可能是进化中后出现的果实类型[28].而最新的石栎系统发育树虽然物种数量丰富[65],但是使用基因数量较少,所以对种间关系支持力度不足,且缺乏形态数据支撑.因此,完善石栎的分子系统发育关系,并把果实形态和果实类型研究与石栎分子系统发育相结合进行祖先状态重建和系统发育信号分析,将为石栎属内关键种的系统位置提供果实形态佐证,明晰2种果实类型的进化关系,同时为分析果实多样性与物种进化的相互关联提供重要证据.
总之,石栎果实形态和果实类型研究不仅是石栎分类鉴定的重要证据,也对理解石栎的种子散播和物种更新、种间联系、进化和物种多样性和形态多样性关联方面有重要作用.在此重提植物形态学的重要性,通过将石栎果实形态研究与生物地理学、系统发育、生态学相结合,为理解石栎物种分化与多样性、物种进化和共存研究提供了一个全新的视角,也为其他物种的进化和多样性研究提供了范本.
