准噶尔盆地南缘无叶假木贼种子雨特征
2017-04-26王婷婷牛攀新江萍李明艳王梅
王婷婷+牛攀新+江萍+李明艳+王梅


摘 要:该文通过布设种子雨收集器并结合统计分析,对准噶尔盆地南缘无叶假木贼种子雨特征和散播规律进行了研究。结果表明,在洪积扇和风沙区,无叶假木贼种子雨的平均累积密度分别达到853粒/m2和751粒/m2,种子散布的高峰期均集中在10月末到11月初的时间段,其落种量分别占整个种子雨的82.42%和76.03%,其后種子雨密度随时间逐渐减小。变异函数分析表明,洪积扇和风沙区的无叶假木贼种子雨分别在8.97m和0.42m的有效变程内,种子雨具有明显的空间格局,洪积扇空间格局由空间自相关和随机因素引起的空间异质性分别占45%和55%,而风沙区空间变异性主要由随机因素引起。准噶尔盆地南缘无叶假木贼种子雨密度大并且在时间上比较集中,同时其时空分布异质性较高,这些特征均将影响无叶假木贼种群的分布格局和种群更新。
关键词:种子雨;散播;无叶假木贼;准噶尔盆地;洪积扇
中图分类号 S718 文献标识码 A 文章编号 1007-7731(2017)07-0016-05
Abstract:The seed rain of Anabasis aphylla on the south edge of the Junggar Basin was studied by laying seed rain traps and using laboratory statistics. The aim was to survey the temporal dynamics and spatial distribution of seed rain to determine the laws and factors impacting seed dispersal. This study focused on the quality and quantity of seeds during different phases of seed rain. The cumulative annual seed rain intensity respectively reached 853 seeds/m2 and 751 seeds/m2 in the diluvial fan and sand area,although in some small local environments it reached as high as 2118 seeds/m2. The peak period of seed rain was from evening October to early November,respectively producing 82.42% and 76.03% of the total seed rain in the diluvial fan and sand area. Subsequently seed rain density decreased gradually with time. The seed rain density stabilized in mid-November and the process of seed rain essentially ended by evening November. Semivariogram analysis of exponential model revealed significant spatial seed rain patterns respectively within an effective range of 8.97m and 0.42m in the diluvial fan and sand area. 45.0% of the spatial heterogeneity was caused by spatial autocorrelation and 55.0% was caused by random factors in the diluvial fan. However,the spatial heterogeneity almost was caused by random factors in the sand area. In summary,A. aphylla produces a high quality and high intensity seed rain at the south edge of Junggar Basin with a relatively high temporal and spatial heterogeneity. These characteristics would be expected to influence the distributional patterns and regeneration of A. aphylla population.
Key words:Seed rain;Dispersal;Anabasis aphylla;Junggar basin;Diluvial fan
种子散播是植物种群通过实生苗更新过程中的重要阶段,扩散的格局直接影响到幼苗的定植和分布,是森林群落更新中繁殖体的主要来源[1-3]。种子散播的初始格局(即种子雨)决定了一个物种潜在的可利用的生境,这为研究种群动态建立了一个样板[4]。种子散播已经被证明在控制种群发展方面起到了显著作用[5]。种子雨直接影响到种子库的大小、质量和格局,也可以用来解释微生境作用下的空间变化,进而影响植物幼苗的定植和更新[6-8]。
种子雨的研究始于20世纪30年代,目前国外已做了大量工作,国外种子雨研究多集中在对一些种群和群落的种子雨[4,9]、种子库和幼苗补充等方面的研究[8,10]。近年来,国内关于荒漠植物种子雨的相关研究多集中在准噶尔盆地东南缘的梭梭[11]、浑善达克沙地的沙地榆[3]、额济纳的胡杨[12]、古尔班通古特沙漠的羽毛针禾[13]等几个物种,而对于无叶假木贼种子雨的研究还没见报道。
无叶假木贼(Anabasis aphylla)种群分布区是温带典型荒漠地带,是适应干旱、半干旱气候的地带性土质荒漠的顶级植物群落[14]。准噶尔盆地南缘的无叶假木贼群落,是防风固沙、保护当地生态环境和周边土地资源的重要的植物群落类型。无叶假木贼是当地洪积扇的主要超旱生小灌木植物种[15]。由于环境的不断恶化及人為的不断干扰,使准噶尔盆地南缘无叶假木贼种群的分布范围逐渐缩小。由于种群数量减少,导致更新上的断层,这对维持洪积扇植被的稳定和持续发展将产生不利的影响。因此,研究无叶假木贼种群种子雨时空格局特征,对于深入分析种群的更新对策和植被的生态恢复与重建具有重要意义[15-16]。
1 研究区概况
研究区位于准噶尔盆地南缘小拐自然保护区北部,该地区有洪积扇和风沙土两种主要的生境类型。洪积扇无叶假木贼群落以无叶假木贼为主达到90%以上,其他的植物种主要有白刺(Nitraria roborowskii)和红砂(Reaumuria soongorica)等,风沙区以无叶假木贼和梭梭(Haloxylon ammodendron)为主(表1)。
2 研究方法
2.1 种子雨样品收集 在准噶尔盆地南缘选择洪积扇和风沙区2个无叶假木贼种子撒播研究区,垂直主风向(西北风)设置样地。样地中无叶假木贼植株密度为该区域的中等程度,样方长宽均为25m。种子收集器的布置采用典型网格法,以5m为间隔,在样方中均匀布置,每个研究区设置36个,共布设72个收集器。种子雨收集器由周长1m的铁丝围成的圆形收集框和网目1mm的白色尼龙网组成。圆形收集框由3根长约35cm的粗铁丝支撑。收集器框口离地面距离为10cm,尼龙网袋深20cm,顺着风向摆放。种子雨收集器的布设于种子扩散前的10月8日完成。种子收集工作于种子自然下落开始,每7d收集一次,至种子雨基本结束止,将收集器中所收集到的种子、同化枝和其他凋落物一同带回,并对收集物进行人工分拣,统计种子的数量。
2.2 种子撒播距离 在准噶尔盆地南缘洪积扇和风沙区各设一个无叶假木贼种子散播距离观测点。在种子扩散前选择结实情况较好的地段,设置1个面积1m×10m的植株样方,用红色染料(自喷漆)对种子进行染色,在入冬第一场雪前,沿东西南北4个方向,每5米设置1m×1m样方,调查样方内染色种子数,调查种子聚居地的生境条件。
2.3 数据处理
2.3.1 种子雨密度的计算 种子雨密度=种子数量/收集面积
2.3.2 种子雨的空间分析 种子雨的空间分布采用变异函数的主要参数块金值、基台值、尺度、空间结构比等来反映,并以此为基础用插值法,分析空间格局[11]。变异函数的主要参数计算使用GS+7.0软件。
3 结果与分析
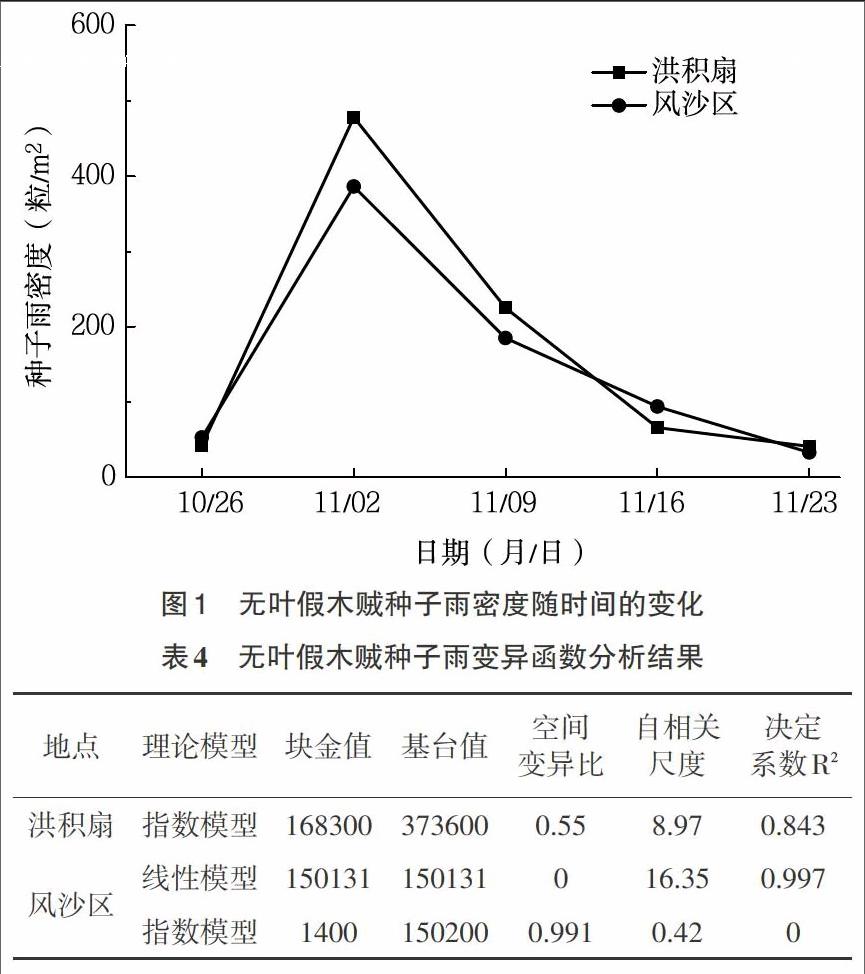
3.1 无叶假木贼种子雨的大小 无叶假木贼种子雨收集工作自2013年10月18日种子自然下落起,至2013年11月末种子雨基本结束止,持续大约40d的时间。洪积扇和风沙区在不同时间不同的种子雨收集框所收集到的无叶假木贼种子雨密度均从最少的0粒/m2到最多的分别为1 225粒/m2和1 014粒/m2,变异较大。从整个种子雨散播过程来看,两个研究区的无叶假木贼种子雨的累积密度达到平均853粒/m2和751粒/m2,累计密度范围分别为122~2 118粒/m2和181~1 887粒/m2,局部小环境种子雨累积密度差异接近18倍(表2、3)。
3.2 无叶假木贼种子雨密度的时间动态 准噶尔盆地南缘无叶假木贼种子开始散布的时间是在10月中旬末,风沙区的种子散播要早于洪积扇。无叶假木贼种子初期成熟量较少,导致种子撒播量在初始阶段较低;而随着成熟量的增加,种子散播量就迅速达到了其整个种子散布过程的最高峰。2个研究区的无叶假木贼种子散布的高峰期集中在10月27日至11月9日时间段,其落种量分别占整个种子雨的82.42%和76.03%(表2、3)。其后,无叶假木贼种子散布量随时间迅速减小,种子散布量保持在较低的水平,到11月末种子散布基本结束,但是植株上仍有2%~5%的未成熟或败育的种子宿存(图1)。
3.3 梭梭种子雨的空间特征 对无叶假木贼种子雨特征进行各向同性的变异函数分析显示,滞后距离大于16.72m时,计算的变异函数值摆动性较大,但在16.72m的滞后距离内,变化具有规律性,即在小的分隔距离内,有较低的变异函数值,随着分隔距离的加大,变异函数值也增大,并逐渐趋于平稳。以16.72m为最大滞后距离进行理论模型拟合,结果显示,2个研究区种子雨的指数模型的自相关尺度分别为8.97m和0.42m。在该自相关尺度内,种子雨具有明显的空间格局,其指数模型空间结构方差与总变异方差的比值分别为为0.55和0.991(表4),这也表明了种子雨由空间自相关和随机因素引起的空间异质性的比例。另外,在这个尺度之外,种子雨在空间上的变化,表现出随机性。总体来讲,洪积扇无叶假木贼种子雨密度的空间异质性由空间自相关和随机因素共同起作用,而风沙区则基本上由随机因素决定。同时,洪积扇无叶假木贼种子雨高密度斑块面积较大,种子在空间上呈聚集分布(图2)。由于洪积扇无叶假木贼种子的指数模型拟合度高为最适模型,而风沙区的线性模型拟合度较高,因此采用对反距离加权平均(IDW)插值法,以两个模型来计算出的基本特征值来制作差值图(图2、3)。
4 讨论与结论
4.1 无叶假木贼种子雨的大小 种群种子雨密度的大小反映了植物结实能力的大小以及更新的潜在能力[11]。在整个种子雨期间,无叶假木贼在洪积扇和风沙区2个地点平均累积种子雨密度分别为为853粒/m2和751粒/m2。由于植物种之间的种子产量的千差万别,不同种群之间的种子雨密度也会相差也较大[17]。例如:准噶尔盆地东南缘梭梭种群年种子雨强度187粒/m2[11];浑善达克沙地的56、40和23年生孤立沙地榆年种子雨强度分别为(7 744±16.26、5 236±18.87、2 456±12.67)粒/m2,表现出了不同龄级沙地榆的种子雨强度有较大差异[3];额济纳胡杨的年种子雨强度在20 115~60 071粒/m2,且不同生境条件下种子雨强度有很大差异[12];阿根廷西部荒漠优势种Bulnesia retama的年种子雨强度在(532.2±81.1)粒/m2[18]。可见,相对于其他荒漠植物种,无叶假木贼种子雨密度处于中等水平,但是考虑到植株体大小,小灌木无叶假木贼种子雨的总量相对较大。此外,无叶假木贼种子雨密度也像额济纳胡杨一样表现出了不同生境之间的差异。
4.2 无叶假木贼种子雨的时间格局 一个种群尽管个体成熟时间参差不齐,种子雨仍有一定的期限[19]。从开始到结束无叶假木贼的整个种子雨散播过程表现出明显的时间异质性。无叶假木贼种子雨散播过程从10月中旬末期开始,种子经历7d的充分成熟和气候干燥后迅速达到种子雨量的最大值,其后散落量逐渐减少,到11月底基本结束,历时约40d的时间。不同的树种种子雨降落的动态也不相同,梭梭种子雨散播过程从11月初开始,并迅速达到种子雨量的最大值,到翌年的1月下旬基本结束,历时近3个月时间[11]。浑善达克沙地沙地榆种子从5月18日开始至7月2日散播结束,持续了大约45d[3]。额济纳胡杨种子雨在3个生境里开始散播的时间分别为8月5日、8日和7月15日,持续的时间为9~15d,时间较短且生境之间的持续时间和散播初期的差异均较大[12]。相对而言,在荒漠植物里无叶假木贼种子雨持续的时间为偏短类型。
梭梭从种子雨散播过程一开始,种子雨量就达到了整个种子雨散播过程种子雨量的最大值,其后种子雨量随时间逐渐减少,到种子雨后期,种子雨量维持在相对较低的水平[11]。浑善达克沙地沙地榆在整个种子雨过程中,种子雨散播过程初期时间较长且种子雨量较小,其后逐渐增大,直到达到最大值,而种子雨过程后期种子雨量又逐渐减少至较低水平[3]。额济纳胡杨的种子雨散播过程中有一个时间较短的、种子雨量较小的散播初期,而后进入一个持续时间较长的高峰期,最后逐渐消退[12]。无叶假木贼和胡杨种子雨散播过程较为相似,有一个短暂的散播初期,这一点与梭梭种子雨散播过程有所区别。无叶假木贼在7月末至8月初开花,种子成熟较晚,可能在散播初期种子成熟较少,导致初期种子雨量也少。造成这种现象的原因也可能是和无叶假木贼种子雨过程及气象因子有关,据观测在野外偶然的一场大风或降雨(雪)都将会造成无叶假木贼种子的集中大量散落。
4.3 无叶假木贼种子雨的空间格局 在种子雨的格局研究中,采用地统计学的方法研究种群种子雨散布格局是近年来生态学中常用的方法[20]。梭梭种子在散落过程中主要依靠重力和风力的共同作用进行扩散,在群落内部由于不同植株树冠的相互阻挡减小了风的作用力,从而使种子主要降落在母树树冠范围内,而母树的聚集分布直接导致了种子雨的聚集分布[11]。无叶假木贼种子雨陆续下落的种子形成了异质性的空间分布格局,在母株附近,有聚集分布的特征;遠离母株后,倾向于随机分布。
通过种子雨散落的种子或是其他繁殖体,必须经受环境筛的考验,只有那些落入“安全岛”的种子才有生根出苗的机会[21]。从格局尺度看,无叶假木贼种子雨的指数模型的自相关尺度较小(8.97m和0.42m),小尺度的种子雨异质性格局,对种群更新具有深刻的意义。因为这样的格局,使扩散的种子分布于多种小生境或微立地,可躲避密度制约的摄食和死亡,从而有更多的更新机会[22]。洪积扇无叶假木贼植物组成简单、土壤硬实,有利于种子的二次散播最大散播距离可达135m;而风沙区无叶假木贼植株高大、沙土疏松,因而造成洪积扇种子雨聚集分布明显,最大散播距离只有60m。风沙区无叶假木贼种子散播格局的随机性因素较大,而实际情况中影响种子雨散布格局的因素非常复杂,可分为生物因素和非生物因素,前者主要有母树的分布情况[20]、植株高度[23]、种子重量[24]以及动物捕食[25]等。后者主要包括地形[26-28]和风向[29,30]等。要对无叶假木贼种群种子散布格局影响因素进行深入分析,需要将各种影响因素考虑在内,进行更进一步的长期定位研究。
参考文献
[1]李小双,彭明春,党承林.植物自然更新研究进展[J].生态学杂志,2007,26(12):2081-2088.
[2]肖治术,张知彬,王玉山.以种子为繁殖体的植物更新模型研究[J].生态学杂志,2003,22(4):70-75.
[3]谷伟,岳永杰,李钢铁,等.浑善达克沙地沙地榆种子雨的扩散规律[J].生态学报,2012,32(11):3440-3448.
[4]Schupp E W. Seed-seedling conflicts,habitat choice,and patterns of plant recruitment[J].American Journal of Botany,1995,82(3):339-409.
[5]Scherff EJ,Galen C,Stanton M L. Seed dispersal,seedling survival and habitat a nity in a snowbed plant:limits to the distribution of the snow buttercup Ranunculus adoneus[J].Oikos,1994,69(2):405-413.
[6]Howe H F,Miriti M N. No question:seed dispersal matters[J].Trends in Ecology and Evolution,2000,15(11):434-436.
[7]Clark J S,Silman M,Kern R,Macklin E,Lambers J H R. Seed dispersal near and far:patterns across temperate and tropical forests[J].Ecology,1999,80(5):1475-1494.
[8]Seidler T G,Plotkin J B. Seed dispersal and spatial pattern in tropical trees. PLoS Biology,2006,4(11):2132-2137.
[9]Urbanska K M,Erdt S,Fattorini M. Seed rain in natural grassland and adjacent ski run in the Swiss alps:a preliminary report[J].Restoration Ecology,1998,6(2):159-165.
[10]Armesto J J,Díaz I,Papic C,Willson M F. Seed rain of fleshy and dry propagules in different habitats in the temperate rainforests of Chiloé Island,Chile[J].Austral Ecology,2001,26(4):311-320.
[11]吕朝燕,张希明,刘国军,等.准噶尔盆地东南缘梭梭种子雨特征[J].生态学报,2012,32(19):6270-6278.
[12]张玉波,李景文,张昊,等.胡杨种子散布的时空分布格局[J].生态学报,2005,25(8):1994-2000.
[13]龙丽红,王慧,马晓丽,等.古尔班通古特沙漠羽毛针禾种群种子雨特征[J].干旱区研究,2014,31(3):516-522.
[14]黄培佑.从龟裂地植物群落建群现象剖析生态演替中有关问题[J].新疆环境保护,1995,17(3):4-6.
[15]楚光明,李卫红,庄丽,等.假木贼属植物研究进展[J].湖北农业科学,2009,48(4):996-999.
[16]楚光明,王梅,张硕新.准噶尔盆地南缘洪积扇无叶假木贼种群空间点格局[J].林业科学,2014,50(4):8-14.
[17]Urbanska K M,Fattorini M. Seed rain in high-altitude restoration plots in Switzerland[J].Restoration Ecology,2000,8(1):74-79.
[18]Farrell C,Hobbs R J,Colmer T D. Microsite and litter cover effects on seed banks vary with seed size and dispersal mechanisms:implications for revegetation of degraded saline land[J].Plant Ecol,2012,213(7):1145-1155.
[19]邹春静,徐文铎,刘广田.沙地云杉种群种子雨的时空分布规律[J].生态学杂志,1998,17(3):16-19.
[20]刘双,金光泽.小兴安岭阔叶红松林种子雨的时空动态[J].生态学报,2008,28(11):5731-5740.
[21]Harper J L.Population Biology of Plants. London:Academic Press,1977.
[22]Janzen D H. Herbivores and the number of tree species in tropical forests[J].The American Naturalist,1970,104(1):501-528.
[23]高润宏,王国玲,李俊清,等.绵刺繁殖对策与种群更新的生境选择行为研究[J].干旱区资源与环境,2007,21(9):150-155.
[24]李军,郑师章,钱吉,等.野生大豆种子雨的研究[J].應用生态学报,1997,8(4):372-376.
[25]Moles A T,Falster D S,Leishman M R,Westoby M..Small-seeded species produce more seeds per square meter of canopy per year,but not per individual per lifetime[J].Journal of Ecology,2004,92(3):384-396.
[26]杨辉,华鹏,黄培祐.苦豆子种子特性与种群扩展关系的研究[J].干旱区资源与环境,2006,20(1):198-201.
[27]王巍,马克平,高贤明.东灵山地区脊椎动物对辽东栎坚果捕食的时空格局[J].植物学报,2000,42(3):289-293.
[28]刘茂秀,丛者福,魏岩.莫索湾地区梭梭群落种间关联性的研究[J].石河子大学学报(自然科学版),2005,23(4):449-453.
[29]孙秀玲,许志春,才让丹周.云杉矮槲寄生种子雨的时空分布格局[J].西北林学院学报,2014,29(4):65-68.
[30]沈泽昊,吕楠,赵俊.山地常绿落叶阔叶混交林种子雨的地形格局[J].生态学报,2004,24(9):1981-1987.
(责编:张宏民)
