白藜芦醇调控miR-223对坏死性小肠结肠炎小鼠细胞焦亡的作用机制
2021-09-08张莹楠常新捷张英剑
王 瑛,张莹楠,常新捷,张英剑
(河南科技大学第一附属医院 1.儿外科、2.消化内科, 河南 洛阳 471000)
坏死性小肠结肠炎(necrotizing enterocolitis,NEC)是一种常见于早产儿的急性肠道炎症性疾病,是造成新生儿致残、致死的重要因素[1]。流行病学调查统计显示,在新生儿体质量低于1.5 kg的早产儿中NEC发病率高达7%,致死率高达20%-30%[2]。由于NEC病情的反复性、严重性以及复杂性,有关其确切机制尚不完全清楚。目前研究表明,早产、人工喂养、炎症风暴等与NEC的发生发展密切相关[3]。细胞焦亡是一种新型的炎症性、程序性细胞死亡方式,其主要是由caspase-1/3/4/5/11介导GSDMD剪切形成具有细胞毒性的GSDMD-N和无毒性的GSDMD-C,其中GSDMD-N募集至细胞膜与磷脂分子结合形成GSDMD-N细胞孔洞,促进炎症细胞因子IL-1β和IL-18释放,并最终导致机体强烈的炎症反应[4]。在大鼠NEC模型中,细胞焦亡相关蛋白caspase-1、IL-1β和IL-18表达显著升高[5]。另外,在小肠上皮细胞中,脂多糖(lipopolysaccharide,LPS)能够通过促进NLPR3炎症小体活化诱导细胞焦亡并加快小肠上皮细胞死亡,抑制NLRP3能够抑制LPS诱导的小肠上皮细胞焦亡,缓解NEC肠道损伤[6]。以上研究均表明,细胞焦亡参与了NEC肠道损伤的病理过程。因此,通过探寻有效药物调控细胞焦亡可能成为治疗NEC的重要方式。
白藜芦醇(resveratrol,RSV)是一类存在于葡萄、花生、虎杖等多种植物内的多酚类有机化合物。既往研究显示,RSV具有广泛的药理学活性,主要对心血管系统、神经系统、呼吸系统等疾病中的组织和细胞发挥抗凋亡、抑制炎症反应的作用[7]。新近研究显示,RSV能够能够通过抑制NLRP3表达,缓解LPS诱导的人肺上皮细胞焦亡[8]。另外,RSV能够通过调控miR-223p/NLRP3信号减少皮质神经元细胞炎症性死亡,从而发挥神经元保护作用[9]。但是,目前有关RSV对NEC细胞焦亡的影响及其作用机制尚不明确。因此,本研究拟通过构建NEC新生小鼠模型,探究RSV对NEC细胞焦亡的影响及其可能的作用机制,进而为临床治疗提供新的参考依据。
1 材料与方法
1.1 材料
1.1.1动物 选取SPF级,5日龄C57BL/6新生小鼠,体质量为(4.35±0.25)g , 湖南斯莱克景达实验动物有限公司,生产许可证:SCXK(湘)2018-0001。
1.1.2试剂与仪器 白藜芦醇(深圳汇鑫荣生物有限公司,纯度≥99%,批号:090226);配方奶粉(Abbott Laboratories,美国);LPS(Sigma公司,美国);多聚甲醛(碧云天生物技术有限公司,上海);蛋白浓度测定试剂盒(碧云天生物技术有限公司,上海);caspase-1抗体(武汉三鹰,批号:22915-1-AP);GSDMD抗体(武汉三鹰,批号20770-1-AP);IL-1β抗体(Cell Signaling Technology公司,批号:12507);IL-18抗体(Cell Signaling Technology公司,批号:54943);β-actin(碧云天生物技术有限公司,批号:AF5001);山羊抗小鼠IgG二抗(碧云天生物技术有限公司,批号:A0412);山羊抗兔IgG二抗(碧云天生物技术有限公司,批号:A0408);Western blot一抗、二抗稀释液(碧云天生物技术有限公司);ECL化学发光试剂盒(碧云天生物技术有限公司,批号:P0018FS);苏木素伊红(HE)染色试剂盒(碧云天生物技术有限公司,批号:C0105S);小鼠IL-1β ELISA检测试剂盒(碧云天生物技术有限公司,批号:PI301);小鼠IL-18 ELISA检测试剂盒(碧云天生物技术有限公司,批号:PI553);实时荧光定量PCR测定试剂盒(TAKARA,美国)。单功能酶标仪(湖南湘仪技术有限公司);低温、高速离心机(湖南湘仪实验室仪器有限公司);细胞组织碾磨仪(武汉赛维尔生物技术有限公司);Western blot检测装置(美国Bio-Rad伯乐公司);荧光倒置显微镜(日本/奥林巴斯Olympus);ChemiDoc XRS+成像系统(美国 Bio-Rad公司)。
1.2 方法
1.2.1动物分组与给药干预 将5日龄的新生C57BL/6小鼠30只,随机分为对照组(Control)、坏死性小肠结肠炎模型组(NEC)以及RSV药物干预组(RSV),每组小鼠各10只。NEC组小鼠:置于保温箱内,人工喂养,按照参考文献[10]方法进行配方奶粉喂养,每天4次,每次50 μL·g-1,并将小鼠置于100%氮气缺氧箱中,缺氧30 s,随后置于4 ℃冰箱中,冷刺激10 min,每日10 ∶00、14 ∶00、22 ∶00分别行1次缺氧冷刺激,连续3 d;在小鼠出生后6 d和7 d分别给予3 mg·kg-1的的LPS灌胃,双盲法行肠组织病理评分分值≥2分,认为成功构建NEC小鼠模型。RSV药物干预组小鼠于6-11 d,每日1次,每次给予RSV(将RSV溶解于0.5%羧甲基纤维素钠中浓度为0.1 mg·g-1)灌胃处理,于12 d处死小鼠,期间小鼠记录造模前后的体质量,解剖留取小鼠小肠末端回肠组织进行后续研究。
1.2.2各组新生小鼠一般情况观察 观察各组新生小鼠的活动以及胃肠反应状况并记录小鼠体质量。
1.2.3各组小鼠肠组织病理形态学观察 各组小鼠于处理d 12时处死,取回肠组织,肉眼观察小鼠肠组织肠腔内积气,出血情况、坏死以及形态串珠样改变等。取小鼠回肠组织1-2 cm,采用4%的多聚甲醛溶液进行固定,再用不同浓度的乙醇溶液梯度脱水,然后用二甲苯进行组织透明,最后经石蜡包埋,切片,染色。步骤根据试剂盒操作说明进行HE染色操作,光学显微镜下观察小鼠回肠组织形态和结构。
1.2.4肠组织IL-1β、IL-18含量测定 收集各组小鼠肠组织,按照组织质量 ∶裂解液体积比=1 ∶5,裂解组织,4 ℃、12 000×g离心15 min并收集各组上清液按照江苏碧云天公司提供的IL-1β和IL-18试剂盒进行各自浓度测定。
1.2.5Western blot检测肠组织caspase-1、GSDMD、IL-1β和IL-18表达 取各组大鼠肠组织,PBS润洗1-2次,离心,按照肠组织碾磨体积 ∶加入细胞组织裂解液体积=1 ∶5,冰上裂解1 h;4 ℃、12 000×g离心15 min,收集上清液,采用BCA法进行蛋白质浓度定量检测;然后取各组大鼠肠组织总蛋白30 μg,上样至聚丙烯酰胺80 V凝胶电泳,80 V电泳30 min后,转120 V、电泳60 min,至溴酚蓝指示电泳至凝胶最底层。依据蛋白质指示marker和目的蛋白分子量大小进行,切取目的胶条,采用湿转转膜法将目的蛋白转移至PVDF膜上,转膜时长2 h。用5%脱脂牛奶室温封闭2 h,孵育caspase-1抗体(1 ∶1 000),GSDMD抗体(1 ∶1 000),IL-1β抗体(1 ∶500)、IL-18抗体(1 ∶500)以及β-actin抗体(1 ∶2 000),4 ℃过夜,孵育对应的山羊抗小鼠二抗或者山羊抗兔二抗((1 ∶5 000)1-2 h,;摇床摇晃上加TPBS洗涤3次,5 min/次,然后加入显影液,显影并拍照。
1.2.6实时荧光定量PCR检测肠组织miR-223表达 依据RNA提取试剂盒说明书步骤提取小鼠肠组织总RNA,并运用紫外分光光度计检测总RNA的浓度与纯度。以RNA为模板,参照逆转录试剂盒说明书步骤将其逆转录为单链cDNA。再以cDNA为模板,根据广州锐博公司设计合成的PCR引物进行扩增。扩增条件:92 ℃,30 s;92 ℃,5 s;60 ℃,31 s,共进行30个循环。采用7300 System SDS Software 分析数据,根据2-△△Ct值相对定量样本中目的基因表达水平。其中miR-223:F:5′-GGGTGTAAAACGCAGCTCA-3′,R:5′-CGGTCGTGTCAGTTTGTCAA-3′;U6:F:5′-CCTCACTGTCCACCTTCCA-3′,R:5′-GGGTGTAAAACGCAGCTCA-3′。
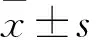
2 结果
2.1 各组新生小鼠一般情况比较Control组新生小鼠生长发育正常,未出现胃肠道症状,体质量逐渐增加。NEC组新生小鼠于造模后3 d出现腹胀、腹泻、稀便等症状,发育迟缓,体质量降低(P<0.01)。RSV干预组新生小鼠体质量增加(P<0.05)(Tab 1),腹胀、腹泻、稀便等症状减轻。
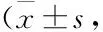
Tab 1 Changes in body mass of each group of newborn mice before and after modeling n=6)
2.2 各组新生小鼠肠道组织大体及病理变化大体观察显示,Control组小鼠肠组织呈现淡黄色、部分出血样改变、肠道无胀气。NEC组小鼠可见肠道明显损伤,肠道胀气明显,肠壁变薄,易破损,呈现串珠样改变。RSV干预组小鼠肠组织大部分呈现淡黄色,弹性尚可,肠道有轻度胀气,有轻度串珠样变化。病理组织切片显示,Control组小鼠肠组织结构清晰正常,肠道绒毛高耸以及上皮完整,黏膜层、黏膜下层和固有层无断裂,充血水肿现象;NEC组小鼠肠组织形态结构紊乱,坏死严重,绒毛变性明显部分脱落并伴有水肿,肌层变薄,断裂等;RSV组小鼠肠组织损伤明显减轻,形态结构排列相对规则,绒毛层次不齐,水肿现象减轻,黏膜层、黏膜下层和固有层未见明显分离(Fig 1)。

Fig 1 Observation of gross and pathological morphology of intestinal tissues of newborn mice in each group (×200)
2.3 RSV对NEC小鼠细胞焦亡的影响与Control组相比,NEC组小鼠肠组织cleaved-caspase-1、GSDMD-N、IL-1β和IL-18蛋白表达上调(P<0.01),caspase-1和GSDMD-N表达差异无统计学意义(P>0.05);与NEC组相比,RSV干预组小鼠肠组织cleaved-caspase-1、GSDMD-N、IL-1β和IL-18蛋白表达下调(P<0.05或P<0.01),caspase-1和GSDMD-N表达差异无统计学意义(P>0.05)(Fig 2)。ELISA检测与Control组相比,NEC组小鼠肠组织IL-1β和IL-18含量增加(P<0.01);与NEC组小鼠相比,RSV组小鼠肠组织IL-1β和IL-18含量降低(P<0.05)(Fig 3)。
2.4 RSV对NEC小鼠miR-223表达的影响与Control组相比,NEC组小鼠肠组织miR-223表达下调(P<0.01);与NEC组相比,RSV组小鼠肠组织miR-223表达上调(P<0.01)(Fig 4)。

Fig 2 Effect of RSV on pyroptosis of intestinal tissue cells in newborn mice n=6)

Fig 3 Effect of RSV on content of IL-1β and IL-18 in intestinal tissues of newborn mice n=6)

Fig 4 Effect of RSV on expression of miR-223 in intestinal tissues of newborn mice n=6)
3 讨论
NEC是由多种因素导致的肠道屏障弱化,炎症反应加剧,上皮损伤并形成恶性循环的一种严重危害早产儿生命健康的获得性胃肠疾病,但是其发病原因尚不清楚。目前,广泛性研究认为缺血缺氧、感染、肠道菌群的失衡等是诱发NEC的重要因素。新近研究表明,炎症反应的失衡亦是导致NEC肠道损伤的重要原因,过度的炎症因子释放导致组织和细胞的炎症级联反应进而诱导细胞死亡,加重NEC的进程[11]。IL-1β能够通过募集、激活免疫细胞,诱发炎症介质释放并介导炎症级联放大反应,最终诱导NEC肠道损伤[12]。另外,TNF-α亦可以通过诱导小鼠小肠上皮细胞凋亡和炎症性死亡诱导小鼠肠黏膜损伤[13]。以上研究表明,炎症在NEC疾病进展中发挥重要作用。因此,明确NEC炎症的发生机制对改善和治疗NEC尤为关键。细胞焦亡作为一种新的炎症性细胞死亡方式,在心血管、肿瘤、呼吸以及神经系统疾病均得到深入研究。组织和细胞产生适度焦亡反应可以维持机体的稳态,一旦细胞焦亡失衡会诱导炎症级联放大反应,加重组织和细胞损伤。对此,本研究首先通过构建NEC新生小鼠模型,探究细胞焦亡与NEC的关系。结果显示,与Control组相比,NEC组小鼠大体观察可见肠道明显损伤,肠道胀气明显,肠壁变薄,易破损,呈现串珠样改变;病理组织切片显示,肠组织形态结构紊乱,坏死严重,绒毛变性明显部分脱落并伴有水肿,肌层变薄,断裂等,表明NEC小鼠模型建立成功。进一步通过Western blot和ELISA实验发现,与Control组相比,NEC组小鼠肠组织cleaved-caspase-1、GSDMD-N、IL-1β和IL-18蛋白表达上调,IL-1β和IL-18含量增加。以上实验结果初步表明细胞焦亡参与了NEC小鼠肠道损伤。RSV具有典型抗炎活性作用,其能够通过抑制炎症因子释放,缓解多种细胞焦亡。与NEC组相比,RSV组小鼠肠组织cleaved-caspase-1、GSDMD-N、IL-1β和IL-18蛋白表达下调,IL-1β和IL-18含量降低,说明RSV能够通过抑制细胞焦亡,缓解NEC小鼠肠道损伤。那么有关RSV对NEC小鼠细胞焦亡的调控机制又是如何呢?
微小RNA(microRNA,miRNA)是一类长约20-24 nt的内源性非编码小RNA,它可通过与靶基因3′非翻译区(3′-UTR)结合,降解靶基因mRNA或抑制其翻译,从而降低靶基因的表达水平,参与调控多种基因表达。既往研究证实miRNA与NEC密切相关。miR-223位于X染色体q12位点,表达受到多个转录因子调控[14]。在一项队列研究中证实43例NEC患者粪便中miR-223表达显著高于非NEC患者[15]。另外,在对NEC患儿外周血白细胞进行高通量测序显示,与对照组相比,NEC组共有482个差异表达miRNA,其中已知58个上调,68个下调,has-miR-223-5p表达与测序结果一致上调[16]。上述研究提示:miR-223可能是NEC的一种重要调控miRNA。研究发现miR-223-3p参与包括免疫系统-炎症反应、血管系统疾病、血液系统疾病等。但是有关miR-223在NEC中的作用机制尚不清楚。本研究通过RT-PCR检测各组新生小鼠肠组织miR-223表达水平,结果显示与Control组相比,NEC组小鼠肠组织miR-223表达下调;与NEC组小鼠相比,RSV组小鼠肠组织miR-223表达上调。本研究结果miR-223表达水平与以往研究不同可能是因为检测标本的差异。既往文献资料显示,在LPS诱导的人肾小管上皮细胞焦亡中,miR-223表达下调,过表达miR-223能够靶向NLRP3抑制LPS诱导的人肾小管上皮细胞焦亡[17]。另外,在大鼠细菌性脑膜炎模型中,RSV能够上调miR-223,抑制NLRP3,进而降低皮至神经元细胞炎性死亡[9]。本研究结果表明,RSV能够抑制NEC小鼠细胞焦亡,缓解小鼠肠道损伤,其可能与上调miR-223表达相关。
综上所述,RSV抑制NEC小鼠肠组织细胞焦亡,缓解NEC肠道损伤,其机制可能与上调miR-223表达相关。以上研究结果初步阐明了RSV在NEC中的作用,进一步丰富了NEC小鼠细胞焦亡的发生机制,以其为NEC治疗提供新的思路。但是有关miR-223是如何调控细胞焦亡尚不清楚。因此,在后续的研究中计划将通过体外培养小鼠小肠上皮细胞,探究过表达miR-223对LPS诱导小肠上皮细胞焦亡的影响,进而为RSV治疗NEC提供可靠的实验依据。
