黑龙江省的西部松嫩平原6个杨树品种光合特征1)
2021-09-07王力刚张玉柱崔琳
王力刚 张玉柱 崔琳
(黑龙江省林业科学院齐齐哈尔分院,齐齐哈尔,161005)
黑龙江省的西部松嫩平原位于我国三北工程东部边缘区—科尔沁沙地北部边缘,属农牧交错区。杨树是该区防护林建设的主要树种,人工选育杨树新品种是其生态建设中重要的基础性工作。经过几十年的人工选育,该区杨树品种单一的状况得到改善,许多杨树新品种陆续被鉴定,并在适宜地区获得推广应用,对生态环境的改善发挥了巨大作用。已有学者对人工选育杨树的生长特性、抗逆性及其繁育技术等方面进行了较为深入细致的研究[1-7],对其光合特性也进行了相关研究,如王福森等[5]研究了DN113杨、小黑杨与其他杨树品种的净光合产物、蒸腾速率、抗旱能力;闫永庆等[8]研究了盐碱胁迫对青山杨光合特性的影响;王晶英等[9]研究了土壤对银中杨光合、蒸腾作用的影响;刘照莹[10]研究了木霉、链格孢菌对山新杨光合特性及相关基因表达的影响;耿丽英等[11]初步测定了小黑杨、青山杨、中黑防杨、银中杨的光合净生产力。这些研究对不同品种杨树的部分光合特性做了较为深入的研究,而针对松嫩平原自然生长的杨树在不同光照强度时光合作用及其对光适应特征的研究鲜见报道,其中关于这些品种生长特性与光合生理的关系尚未见报道。本研究通过盆栽试验,对不同光照强度时人工选育的6类杨树主要光合特性参数进行测定、比较,分析其主要光合特征,并结合其生长特性及生产适应性表现,从植物光合生理学的角度探讨该区人工选育杨树对松嫩平原生态环境的适应性特征,为该区防护林树种选择与推广应用提供参考。
1 研究区概况
研究区位于黑龙江省林科院齐齐哈尔分院(原黑龙江省防护林研究所)试验基地,坐落于黑龙江省的西部松嫩平原,地理坐标47°23′N、123°51′E,年平均气温3.5 ℃,≥10 ℃年平均积温2 726 ℃,无霜期136 d,年平均降水量418.1 mm,年平均蒸发量1 483.0 mm,属半干旱风沙草原区。土壤为碳酸盐栗钙土,土壤密度1.31 g·cm-3。
2 材料与方法
2.1 试验材料
试验材料为该区自20世纪70年代以来选育并通过鉴定及推广应用的6个杨树品种:小黑杨(Populussimonii×P.nigra)、银中杨(Populusalba×P.berolinensis)、山新杨(Populusdavidiana×P.bolleana)、青山杨(Populuspseudo-cathayana×P.deltoides)、中黑防杨(Populusdeltoides×P.cathayana)、DN113杨(Populus×euramericanacv.‘Dn113’)。
2.2 试验方法
为了便于管理与比较,本研究采用盆栽试验。选定内径55 cm,高50 cm的瓦盆48个,每个品种8个,瓦盆内装满该区野外农田土壤(碳酸盐栗钙土)备用。每个杨树品种选取8株健康2年生母根,于春季(4月中旬)栽植于备好的瓦盆内。栽植后,先将瓦盆放于温室内,便于统一浇水、松土、拔草等管理,同时保证栽植后的试验苗木拥有良好的生长环境,能够快速生长发育,达到测定条件。8月中旬,各品种均进入生长旺季,母根从萌发开始生长至1 m以上,此时开始测定工作。
采用美国拉哥公司(LI-COR)Li-6400便携式光合测定仪器进行测定工作。Li-6400光合测定系统可调节光照、CO2、H2O、温度等环境因子,为了比较不同杨树品种在一定光照强度的光合参数变化及差异,利用仪器自带可调和光源设置了6个不同光照强度,分别为0、50、200、500、1 000、1 800 μmol·m-2·s-1,各光照强度标准误控制在±2 μmol·m-2·s-1内。试验主要测定各品种叶片光合速率(Pn)、蒸腾速率(Tr)。从栽植的每个品种中选择生长良好,长势正常的3株标准株,在每个标准株上选取中上部生长健康的新生叶进行活体测定。每个标准株选3片标准功能叶,其平均值作为该株相应指标的标准值。3株标准株测定值的均值作为该品种的标准值。
2.3 数据处理
通过EXCEL完成数据处理,计算各品种叶片在测定光照强度范围的平均光合速率、蒸腾速率、叶片水分利用效率;分析各杨树品种的光合参数随光强梯度变化的规律及特性。
3 结果与分析
3.1 净光合速率随光照强度变化特征
由净光合速率测定结果(见图1)可知,净光合速率随光照强度的增强而逐渐增大,且各品种间呈现出不同程度的差异。青山杨、DN113杨、银中杨3条趋势线较接近,净光合速率一直处于较高值;山新杨、中黑防杨净光合速率曲线较接近,处于次高值;小黑杨净光合速率曲线与其他品种差距较大,一直处于较低值。净光合速率平均值由大到小依次为:DN113杨(11.35 μmol·m-2·s-1)、青山杨(10.93 μmol·m-2·s-1)、银中杨(10.52 μmol·m-2·s-1)、山新杨(8.71 μmol·m-2·s-1)、中黑防杨(8.42 μmol·m-2·s-1)、小黑杨(1.67 μmol·m-2·s-1)。其中,DN113杨、青山杨、银中杨净光合速率较高,为10.5 μmol·m-2·s-1以上;山新杨、中黑防杨为8.4~9.0 μmol·m-2·s-1;小黑杨净光合速率最低,仅为1.67 μmol·m-2·s-1。小黑杨净光合速率在光照强度200 μmol·m-2·s-1左右时仍为负值,表明此时小黑杨的光合速率小于呼吸速率。其他品种在光照为50 μmol·m-2·s-1时仍有一定程度的净光合速率。当光照强度为200 μmol·m-2·s-1时,DN113杨、银中杨净光合速率达8.0 μmol·m-2·s-1以上,此时小黑杨净光合速率仅为-0.117 μmol·m-2·s-1;当光照强度达到1 800 μmol·m-2·s-1时,各品种净光合速率均达到较大值,按最大值到最小值排列依次为:青山杨24.50 μmol·m-2·s-1、DN113杨23.00 μmol·m-2·s-1、银中杨21.40 μmol·m-2·s-1、中黑防杨18.10 μmol·m-2·s-1、山新杨17.70 μmol·m-2·s-1、小黑杨8.97 μmol·m-2·s-1。随着光照强度的增大,各品种的净光合速率均持续增大;当光照强度超过1 000 μmol·m-2·s-1后,除青山杨外,其他各品种的净光合速率增幅变缓。
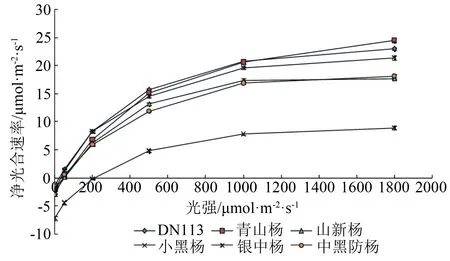
图1 6个杨树品种净光合速率随光强变化特征
3.2 叶片蒸腾速率随光强变化特征
由蒸腾速率测定结果(图2)可知,蒸腾速率随着光照强度递增呈逐渐增大趋势,各品种间呈现不同程度差异。其中,DN113杨、山新杨在光照强度1 000 μmol·m-2·s-1时,蒸腾速率达到1个峰值,分别为4.74、4.57 mmol·m-2·s-1,之后随着光强继续增大而缓慢降低;其他品种的蒸腾速率均随着光照强度递增,持续增大。图2可看出,光照强度为0~50 μmol·m-2·s-1时,各品种蒸腾速率增幅较大(0.55~1.00 mmol·m-2·s-1);光照强度为50~1 800 μmol·m-2·s-1时,各品种蒸腾速率增幅较光照强度增幅相比,平缓很多(0.52~1.02 mmol·m-2·s-1)。由此可知,青山杨一直处于较高的蒸腾速率状态,小黑杨一直处于较低的蒸腾速率状态。6个品种的杨树平均蒸腾速率由大到小依次为:青山杨(4.87 mmol·m-2·s-1)、中黑防杨(4.47 mmol·m-2·s-1)、DN113杨(4.22 mmol·m-2·s-1)、山新杨(4.10 mmol·m-2·s-1)、银中杨(4.01 mmol·m-2·s-1)、小黑杨(2.87 mmol·m-2·s-1)。蒸腾速率作为树木水分生理的重要指标,是树木对水分生理需求的量度,较高的蒸腾速率反映较高生理耗水需求[12-13]。可见,从植物生理来看,青山杨、中黑防杨、DN113杨属于相对耗水品种,小黑杨则属于相对节水品种。

图2 6个杨树品种蒸腾速率随光强变化特征
3.3 叶片水分利用效率随光照强度变化特征
树木叶片水分利用效率(WU,E)指单位水量叶片蒸腾散失时光合作用所同化CO2的量,为光合速率与蒸腾速率的比值(Pn∶Tr)[14]。通过各光照强度对应的净光合速率与蒸腾速率,可计算出各光照强度对应的各品种叶片水分利用效率(图3)。随光照强度从低到高递增,各品种的水分利用效率呈现与净光合速率类似的变化趋势—逐渐增大。光照强度为0~500 μmol·m-2·s-1时增幅较大(3.423~5.117 μmol·mmol-1);光照强度为500~1 800 μmol·m-2·s-1时增幅比较平缓(0.917~1.464 μmol·mmol-1),DN113杨仅为中黑防杨的1/3~1/4。光照强度为1 800 μmol·m-2·s-1时,各品种水分利用效率达到极值,其中DN113杨为4.968 μmol·mmol-1、银中杨为4.693 μmol·mmol-1、青山杨为4.422 μmol·mmol-1、山新杨为3.960 μmol·mmol-1、中黑防杨为3.515 μmol·mmol-1、小黑杨为2.710 μmol·mmol-1。6个品种的杨树平均水分利用效率由大到小排列依次为:DN113杨(2.46 μmol·mmol-1)、银中杨(2.37 μmol·mmol-1)、青山杨(2.04 μmol·mmol-1)、山新杨(1.94 μmol·mmol-1)、中黑防杨(1.70 μmol·mmol-1)、小黑杨(0.27 μmol·mmol-1)。可见,DN113杨、银中杨、青山杨是水分利用效率较高的品种,小黑杨、中黑防杨是水分利用效率相对较低的品种。
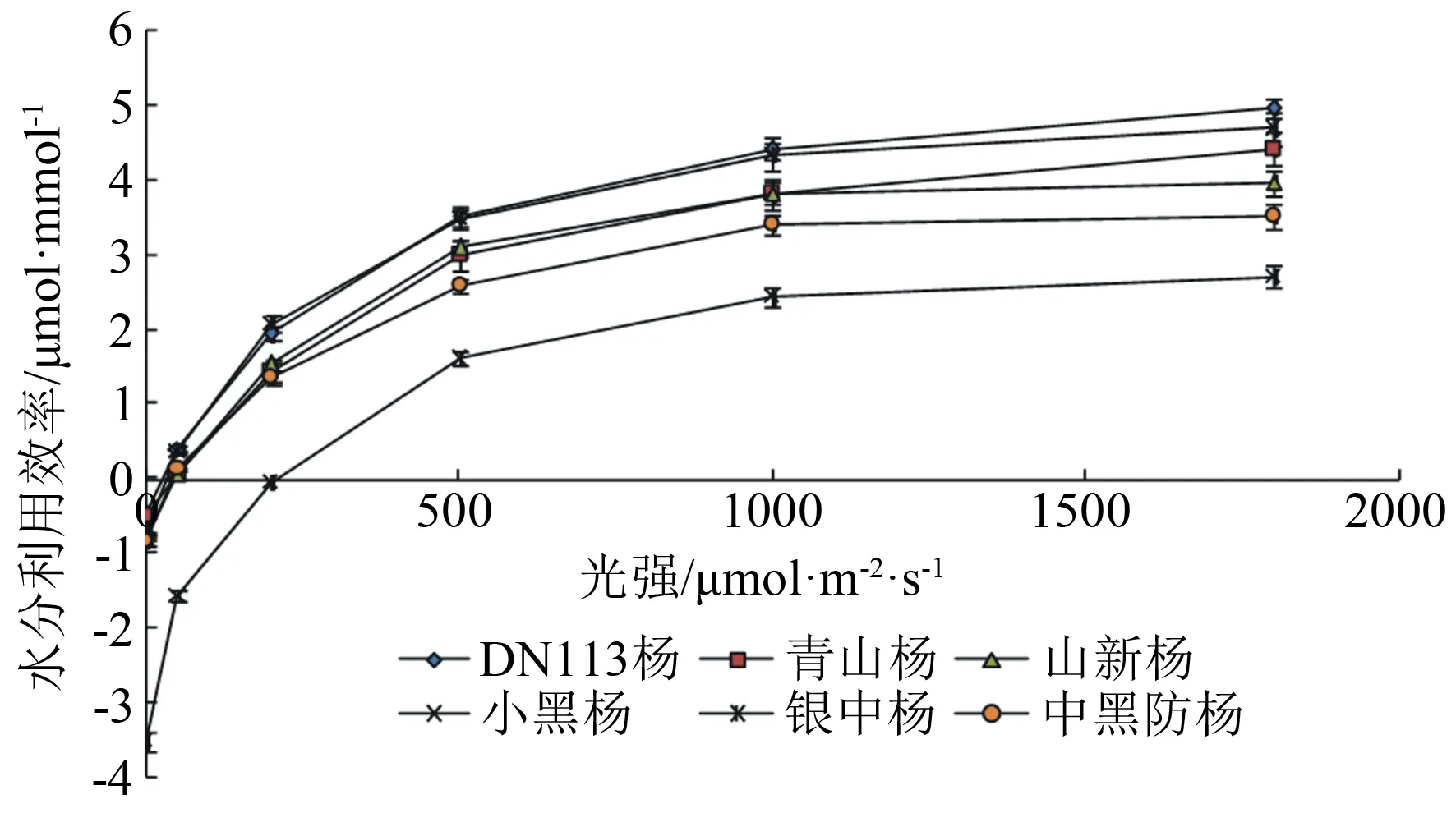
图3 6个杨树品种水分利用效率随光强变化特征
3.4 不同品种光补偿点与光饱和点比较
不同品种杨树对光适应性的多样性表现为光合作用特征不同,即不同品种的光补偿点、光饱和点不同[15]。通过SPSS软件拟合净光合速率与光照强度关系曲线,构建相关模型,计算各品种的光补偿点、光饱和点等光合生理特征值,结果见表1。DN113杨、青山杨、银中杨光饱和点较高,其值分别为1 835.0、1 800.0、1 790.0 μmol·m-2·s-1,比较接近,仅相差45 μmol·m-2·s-1,处于前3位;山新杨、中黑防杨、小黑杨光饱和点较低,其值分别为1 640.0、1 520.0、1 516.7 μmol·m-2·s-1,处于后3位,其中,小黑杨光饱和点值最低,中黑防杨次之,二者较为接近,相差不到5 μmol·m-2·s-1。对于光补偿点,DN113杨最低,其值为18.9 μmol·m-2·s-1;小黑杨最高,其值为210.8 μmol·m-2·s-1;其余几个品种比较接近,银中杨为31.0 μmol·m-2·s-1、青山杨为38.3 μmol·m-2·s-1、中黑防杨为41.8 μmol·m-2·s-1、山新杨为42.0 μmol·m-2·s-1。
对于光补偿点,除小黑杨外,其他品种均较低;对于光饱和点,除小黑杨、中黑防杨外,其他品种均较高,对光的适应范围较广。DN113杨、青山杨、银中杨3个速生品种明显表现为光补偿点较低,光饱和点较高,其中DN113杨光饱和点最高,光补偿点最低,在相对较弱的光照时也可实现营养物质积累。小黑杨光补偿点最高,光饱和点最低,对光的适应范围相对最窄,表现为其生长量明显低于其他品种,与实际生产表现基本吻合[1,4,7]。可见,速生杨树品种应具备较强的光适应能力。
光达到饱和点时的光合速率体现了树木最大光合作用能力。由表1可知,光达到饱和点时,各品种的光合速率有明显差异。DN113杨、青山杨、银中杨净光合速率均超过30 μmol·m-2·s-1,其中,DN113杨净光合速率最大(32.98 μmol·m-2·s-1);其次为青山杨(31.04 μmol·m-2·s-1)、银中杨(30.94 μmol·m-2·s-1);山新杨、中黑防杨净光合速率均超过20 μmol·m-2·s-1,分别为25.54、21.85 μmol·m-2·s-1;小黑杨最低(10.23 μmol·m-2·s-1),与其他品种差距较大,仅为DN113杨、青山杨、银中杨的1/3,山新杨、中黑防杨的1/2。可见,光合作用能力最强的为DN113杨、青山杨、银中杨;其次为山新杨、中黑防杨;小黑杨最弱。
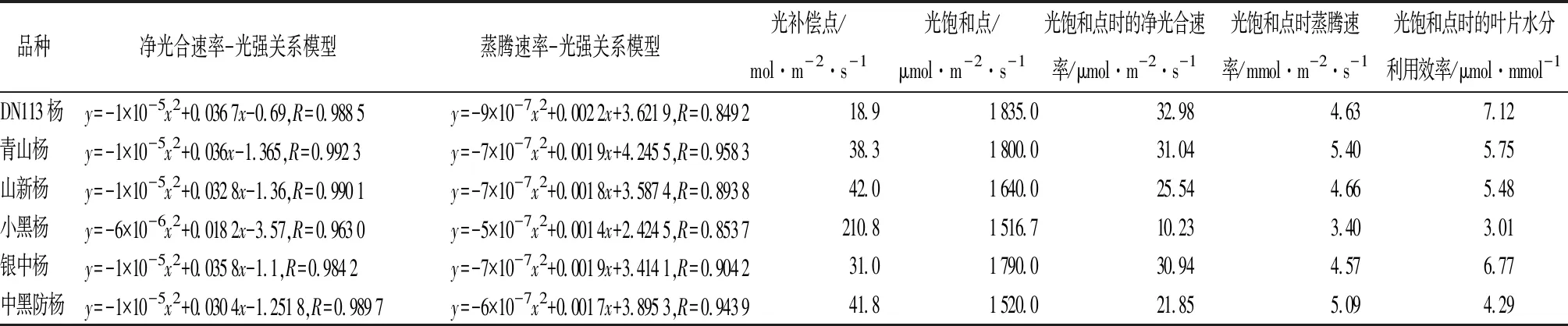
表1 6个杨树品种主要光合特性比较
光达到饱和时叶片水分利用效率是树木重要的光合特征之一。光达到饱和时,各品种的水分利用效率由大到小排列依次为DN113杨(7.12 μmol·mmol-1)、银中杨(6.77 μmol·mmol-1)、青山杨(5.75 μmol·mmol-1)、山新杨(5.48 μmol·mmol-1)、中黑防杨(4.29 μmol·mmol-1)、小黑杨(3.01 μmol·mmol-1),与平均水分利用效率次序一致。
4 讨论
杨树因速生、易繁殖成为我国北方主要人工造林树种,但不同品种间差距较大,其生长特性在其光合生理特征上有所体现。本研究针对黑龙江省的西部6个人工选育的杨树品种,通过主要光合生理参数测定,比较分析可知,净光合速率以DN113杨、青山杨、银中杨较高,均值达10.5 μmol·m-2·s-1以上;山新杨、中黑防杨均值为8.4~9.0 μmol·m-2·s-1;小黑杨均值最低,不到2.0 μmol·m-2·s-1。小黑杨的净光合速率与其他品种有较大差别,与其生长性状表现较为吻合[1,4,7]。蒸腾速率以青山杨、中黑防杨、DN113杨较高,均值达4.2 mmol·m-2·s-1以上;山新杨、银中杨次之,为4.0~4.1 mmol·m-2·s-1;小黑杨最低,不到3.0 mmol·m-2·s-1。蒸腾速率作为树木水分生理的指标,其大小反映了树木对水分的生理需求量度[12-13]。小黑杨较低的蒸腾速率表明其对水分需求相对较小,属节水型品种;银中杨次之;青山杨、中黑防杨、DN113杨则为相对耗水型品种,需要水分充足的条件才能保证其正常生长及速生。叶片水分利用效率以DN113杨、银中杨、青山杨较高,其均值分别为2.46、2.37、2.04 μmol·mmol-1;山新杨、中黑防杨次之,分别为1.94、1.70 μmol·mmol-1;小黑杨为最低,为0.27 μmol·mmol-1,仅为前几个品种的1/6~1/9。6个杨树品种中,小黑杨的净光合速率、蒸腾速率及叶片水分利用效率最低,这主要是由树种本身遗传特性所决定,也是导致其生长相对较慢的直接原因。同样时间段内,小黑杨干物质积累量最少,但其材质相对其他品种较好,其抗逆性及适应性也最强。
从各杨树品种的父母本来源看,小黑杨为小叶杨和欧洲黑杨杂交种,小叶杨为母本,属青杨派,欧洲黑杨为父本,属黑杨派;银中杨为银白杨与中东杨杂交种,银白杨为母本,属白杨派,中东杨为父本,属黑杨派,银中杨则属白杨派品种,是白杨派与黑杨派远缘杂交的一项重大突破;山新杨为山杨和新疆杨的杂交种,山杨为母本,为白杨派中的山杨亚派,新疆杨为父本,为白杨派中的白杨亚派;青山杨为拟青杨与山海关杨的杂交种,拟青杨为母本,属青杨派,山海关杨为父本,属黑杨派;中黑防杨为美洲黑杨与青杨的杂交种,美洲黑杨为母本,属黑杨派,青杨为父本,属青杨派;DN113杨是欧美杨在我国北方寒冷地区的优良无性系,是黑龙江省首次成功引种驯化的纯种欧美杨品种,属黑杨派。净光合速率较大的青山杨、银中杨,父本分别是山海关杨、中东杨;DN113杨属欧美杨,均具有速生的特征,此3个品种由于更多地遗传了其亲本中速生一方的特性而表现出较高的净光合速率。山新杨、中黑防杨的净光合速率相对较低;小黑杨最低,原因是小黑杨的2个亲本小叶杨、欧洲黑杨与前几个品种亲本相比,速生性相对较差。关于不同杨树品种光合特性与其亲本来源的关联性研究尚待深入研究,以便对其生长与适应性特征作出更为科学的揭示。
各品种所表现出的光合特征差别与其光补偿点、光饱和点特征密切相关。小黑杨光补偿点为210.8 μmol·m-2·s-1,为几个品种中最高,是其他品种的5~11倍;光饱和点为1 516.7 μmol·m-2·s-1,为几个品种中最低。树木叶片的光饱和点反映了树木对光照的要求,一般光补偿点、光饱和点均较低的植物是典型的耐阴性植物,反之为典型的喜阳性植物。光补偿点较低、光饱和点较高的品种对光环境的适应性较强;光补偿点较高、光饱和点较低的植物对光照的适应性较差[16]。本研究中,DN113杨、青山杨、银中杨是光补偿点较低(18.9~38.3 μmol·m-2·s-1)、光饱和点较高(1 835.0~1 790.0 μmol·m-2·s-1)的品种,表明这些品种对光照的适应性较强,在较宽光照幅度内均可积累干物质。在光饱和点时,这3个品种的净光合速率较高,达30 μmol·m-2·s-1以上,为其速生奠定了光合生理基础。山新杨、中黑防杨对光照的适应幅度次之,而小黑杨对光照的适应幅度相对最窄,在光饱和点时其净光合速率仅为10.23 μmol·m-2·s-1,为DN113杨、青山杨、银中杨的1/3左右,这正是小黑杨生长相对其他几个品种生长较慢的主要原因。
黑龙江省的西部松嫩平原属半干旱地区,节水抗旱一直是该区林木良种选育的首要目标。杨树作为该区主要造林树种,选育节水抗旱的优良杨树品种成为该区杨树育种工作的重点。小黑杨作为20世纪70年代该区选育出的优良杨树品种[17],尽管在生长性状方面不如其他品种,但经过几十年的时间验证以及区域上大面积的推广应用,经受住了全省大部分平原区的抗逆性考验,适应性强,抗旱、抗寒性表现良好[5],是该区杨树造林的主要品种。其他品种的杨树除生长指标外,在适应性、抗逆性方面与小黑杨相比有一定差距,推广应用时受环境条件的限制,尤其是水分条件。DN113杨、青山杨、中黑防杨等速生树种在水肥条件较好区域表现出良好的生长性状。银中杨生长速度较快且抗逆性亦较强,目前被广泛应用于城乡绿化及生产造林,推广应用区域逐年扩大,已成为三北地区主要造林绿化树种之一。山新杨生长与抗逆性方面均处于中等,因其树形优美,更适于园林绿化。
5 结论
本研究通过比较研究黑龙江省的西部松嫩平原6个杨树品种在0~1 800 μmol·m-2·s-1光照强度的主要光合生理特征,探究了其生长性状的光合生理机制。结果表明,净光合速率均值由大到小依次为:DN113杨、青山杨、银中杨、山新杨、中黑防杨、小黑杨;蒸腾速率均值由大到小依次为青山杨、中黑防杨、DN113杨、山新杨、银中杨、小黑杨;水分利用效率均值由大到小依次为DN113杨、银中杨、青山杨、山新杨、中黑防杨、小黑杨。比较而言,DN113杨、青山杨属净光合速率、蒸腾速率、水分利用效率均较高品种;银中杨属净光合速率、水分利用效率较高,蒸腾速率中等品种;山新杨属净光合速率、蒸腾速率、水分利用效率均为中等品种;中黑防杨属净光合速率中等、蒸腾速率较高、水分利用效率中等品种;小黑杨为净光合速率、蒸腾速率、水分利用效率均较低品种。6个杨树品种中,DN113杨、青山杨、银中杨光补偿点较低、光饱和点较高,对光照适应幅度较宽;山新杨、中黑防杨次之;小黑杨光补偿点相对最高,而光饱和点相对最低,对光照的适应幅度相对最窄。本研究探索了6个杨树品种生长性状差别的光合生理机制,为其在适宜地区推广应用提供了参考。
