中国特有属苞藜属的系统学研究
2021-09-04李波卡冯虎元潘建斌
李波卡,冯虎元,潘建斌*
(1 兰州大学 生命科学学院,兰州 730000;2 中国科学院 植物研究所,系统与进化植物学国家重点实验室,北京 100093)
藜科(Chenopodiaceae Vent.)植物约有130余属1 500余种,主要分布于南部非洲、中亚、北美洲、南美洲及大洋洲的干草地、荒漠、盐碱地,以及地中海、黑海、红海沿岸。传统上,苋科(Amaranthaceae Juss.)被认为是藜科的近缘类群。而分子证据显示,藜科的多节草亚科(Polycnemoideae Ulbr.)与苋科成姊妹群关系[1],表明藜科是并系群。因此自APGⅡ系统开始[2],狭义藜科和苋科合并成广义苋科。为避免混乱,以下采用多识植物分类系统[3]中的广义苋科分类框架进行阐述。
苞藜属(BaoliaKung et G. L. Chu)是根据1978年孔宪武等在甘肃省迭部县发现的苞藜(BaoliabracteataKung et G. L. Chu)新种建立的单型属[4],由于采样困难等诸多因素,尚无分子水平的研究。形态学上苞藜属通过不同的特点被认为与不同的类群具有较近的亲缘关系:苞藜具有平坦的叶、两性花、5枚雄蕊、环形胚,这些特征与藜族(Chenopodieae Dumort.)一致;但其每朵花均具2枚膜质小苞片及1枚苞片,显示其具有碱蓬亚族(Suaedinae Dumort.)的特征。此外,苞藜属种皮木质且增厚,表面有独特的小凹陷,表明其可能与环翅蓬属(BienertiaBunge ex Boiss.)有较近的关系。《中国植物志》[5]将苞藜属放在环胚亚科(Cyclolobeae C. A. Mey.)藜族。之后,朱格麟[6]对苞藜属的系统位置进行修订,将其放在了环胚亚科多节草族(Polycnemeae Dum.)。Kühn的系统中苞藜属位于藜亚科(Chenopodioideae Burnett)[7],而Zhu等[8]认为,苞藜属应被放在碱蓬亚科(Suaedioideae Ulbr.)苞藜族(Baolieae Kung)。多识植物分类系统则将苞藜属置于藜亚科藜族[3](表1)。上述所有观点均缺少分子系统学证据,因此,通过分子手段确定苞藜属的系统位置非常有必要。
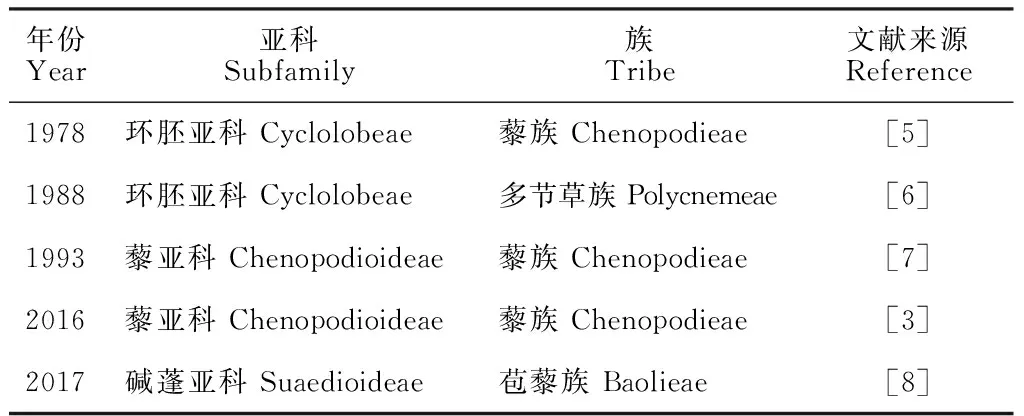
表1 苞藜属的分类历史
分子钟定年方法现已广泛用于生物类群分化年代的推断,该方法可通过整合化石证据从而得到各节点的绝对分化时间与置信区间[9]。藜粉(ChenopodipollismultiplexKrutzsch)[10]和拟盐角(SalicornitesmassalongoiPrincipi)[11]是2个年代分别为65~56 Ma和35~23 Ma的藜亚科植物化石记录,在过去的研究中已被用于推测系统发育树节点分化年代[1, 12]。传统上,苞藜属因其较原始的形态学特征而被认为是一个原始属[13]。本研究基于核基因片段(ITS)和叶绿体基因片段(atpB-rbcL,rbcL)等序列构建苋科物种系统发育树,同时采用上述化石记录对分子钟进行校正,探讨苞藜属的系统位置与分化时间。
1 材料和方法
1.1 实验材料
苋科植物材料取自野外(表2),使用硅胶对采集的新鲜叶片进行快速干燥,之后在实验室完成DNA的提取。
1.2 DNA提取和样品测序
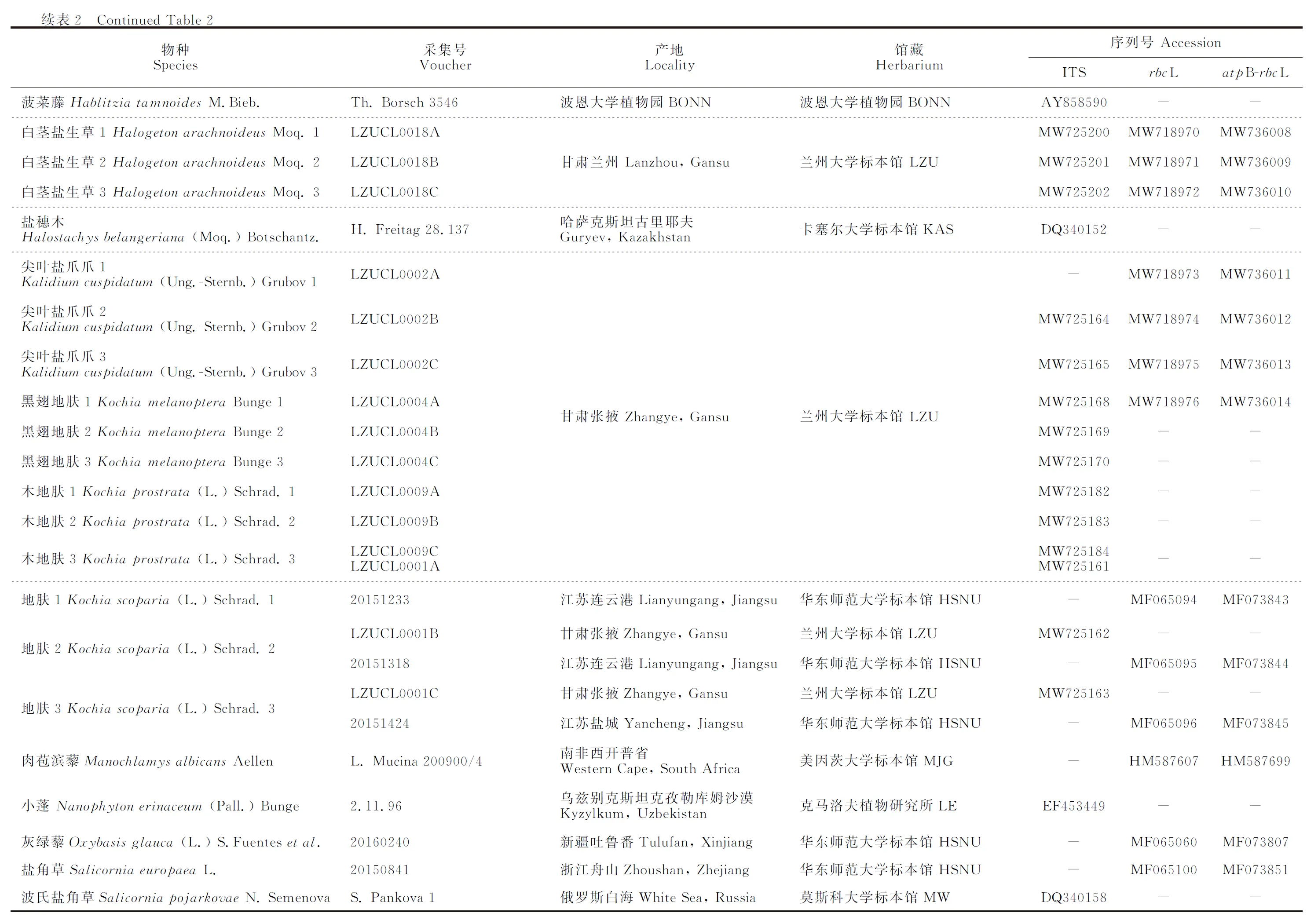
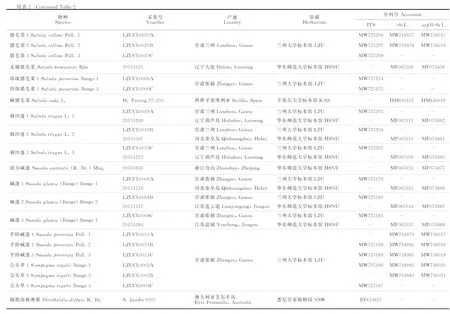
选取核糖体RNA基因转录间隔区ITS、叶绿体rbcL和叶绿体非编码区atpB-rbcL片段,进行序列扩增、测序与系统发育分析。除对所采集物种进行测序外,其他物种的基因序列下载自GenBank数据库(表2)。
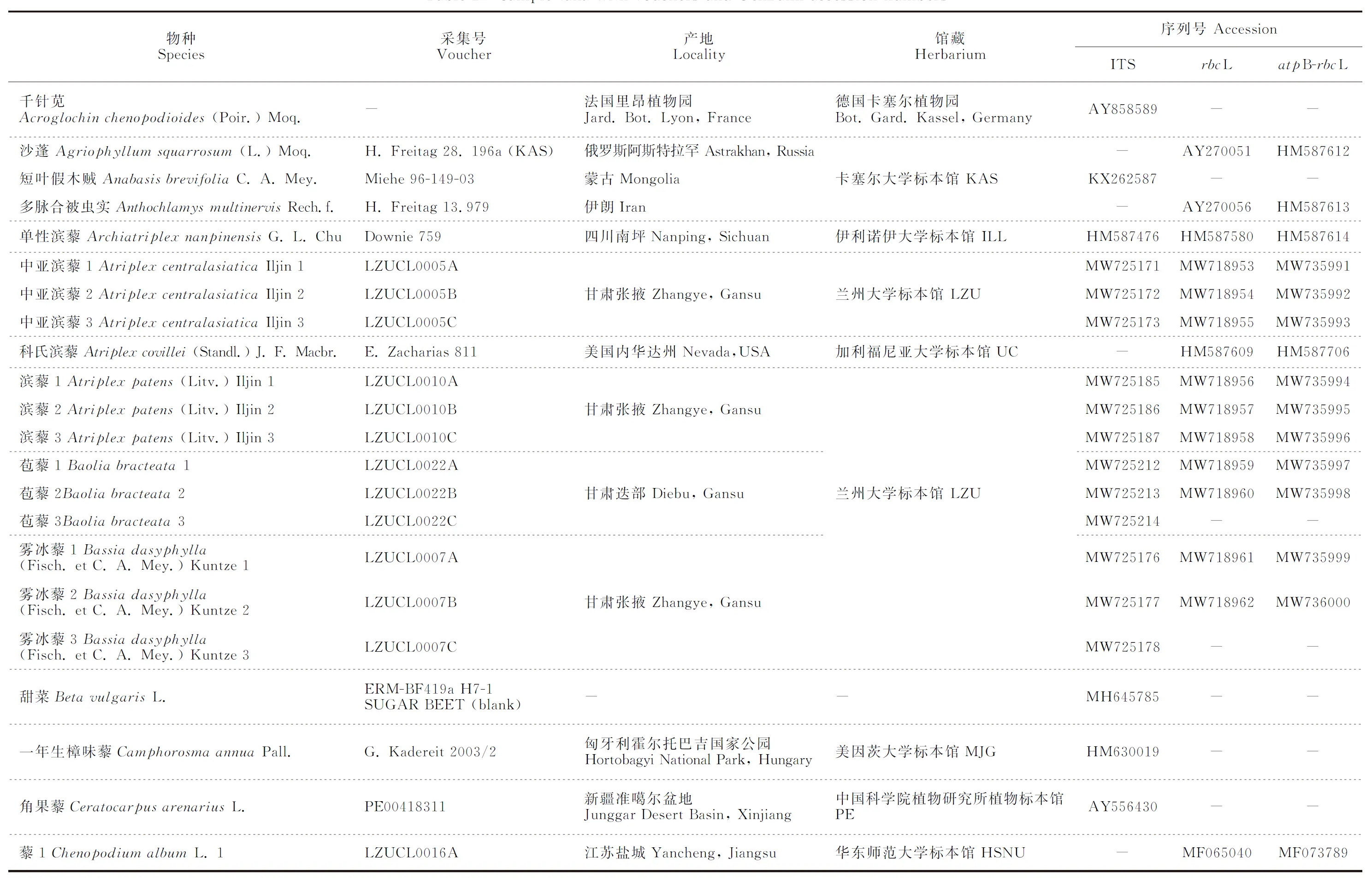
表2 实验材料来源与GenBank序列号

1)总DNA提取。使用天根生化科技有限公司生产的植物基因组DNA提取试剂盒完成总DNA的提取。
2)目的片段扩增。采用文献[14]中的引物与PCR反应条件,完成ITS、rbcL、atpB-rbcL三个目的片段的扩增。PCR过程使用 2720 热循环仪,反应体系 40 μL:DNA 2 μL,dNTPs 10 mmol/L,引物 2 μL(每种引物5 μmol/L), DNA聚合酶4 U,缓冲液4 μL,ddH2O 26.4 μL。取200~1 000 μg基因组DNA完成对ITS、rbcL、atpB-rbcL片段的扩增。
3)纯化与测序。使用天根生化科技有限公司生产的通用型DNA纯化回收试剂盒完成PCR产物纯化,由北京擎科新业生物技术有限公司完成测序。
1.3 系统发育分析
MEGA 7[15]软件内置的Muscle程序被用于多序列比对,比对结果经手动调整后保存为fasta格式。rbcL、atpB-rbcL两个数据集采用多基因联合分析,经Sequence Matrix V1.7.8[16]完成拼接后组装为一个序列矩阵。ITS 序列与rbcL、atpB-rbcL拼接后的序列数据使用PGDspider V2.1.1.5[17]转换成nexus格式,分别用于Beast V1.10.4[18]进行贝叶斯树构建,用IQ-TREE[19]进行最大似然树的构建。
在贝叶斯树的构建中,使用PAUP V4.0[20]对ITS、atpB-rbcL+rbcL两个序列矩阵的nexus文件进行打分测试,将得到的打分(score)文件使用Mrmodeltest V2.3[21]选出最适核酸替代模型(在AIC标准下,ITS数据为 GTR+I+G,atpB-rbcL数据为GTR+G,rbcL数据为GTR+I+G)。Beast V1.10.4[18]用于对上述核基因ITS数据与叶绿体基因atpB-rbcL+rbcL数据进行贝叶斯分析与分化时间评估。运行Beast V1.10.4软件内置的 BEAUti V1.10.4程序生成xml文件。具体步骤如下:(1)在替代模型的选择中,atpB-rbcL设定为GTR+G模型,ITS、rbcL均设定为GTR+I+G;(2)所有数据集采用对数正态分布的非相关宽松分子钟模型(uncorrelated relaxed log normal molecular clock model)[22];(3)树先验(tree prior)选择生灭过程(Birth-Death Process)[23],并使用随机起始树(random starting tree);(4)采用年代(百万年,Ma)分别为 65~56 Ma 和35~23 Ma的藜粉(Chenopodipollismultiplex)和拟盐角(Salicornitesmassalongoi)2个化石记录,分别标定系统发育树根节点以及盐角草亚族(Salicorniinae Ulbr.)冠群分化年代;(5)系统发育树根部及盐角草亚族年代均设置为正态分布,均值/标准差分别设置为:60.5/2和29/2.5,其他先验采用默认设置;(6)执行参数(Operators)采用默认设置;(7)马尔可夫链-蒙特卡洛算法运行代数为8×107,抽样频率为每1 000代。生成的xml文件经Beast V1.10.4运行后,使用Tracer V1.7.1[24]对各参数的有效样本容量(effective sample size,ESS)进行收敛性(convergence)评估,老化样本(burn-in)设置为10%,使用TreeAnnotator V1.10.4程序总结最大分支可信度树(maximum clade credibility tree)以及各节点的后验概率(posterior probability,PP)、95%最高后验密度区间(highest posterior density heights interval, HPD)以及Median heights。
使用IQ-TREE提供的WEB服务器[25]完成最大似然树的构建。核酸替代模型选择采用IQ-TREE整合的ModelFinder模块[26],在BIC标准下,ITS数据的最适模型为SYM+I+G4,atpB-rbcL数据的最适模型为 K3Pu+F+G4,rbcL数据的最适模型为TPM3u+F+I+G4。使用IQ-TREE支持的超快自展(ultrafast bootstrap)[27],自展值(bootstrap,BS)设置为1000。系统发育树分支中,贝叶斯树后验概率≥0.95或最大似然树自展值≥95视为高支持率。
2 结果与分析
2.1 基于ITS序列的系统发育分析
ITS序列矩阵包含物种33个,67条序列。序列长度为686 bp,其中变异位点390个(56.9%),保守位点286个(41.7%)。通过Beast分析得到的系统发育树见图1,最大似然树与贝叶斯树拓扑结构相似,不同之处是平卧碱蓬(Suaedaprostrata)以低支持率(BS=76)与猪毛菜族(Salsoleae Dumort.)聚成一支。结果显示,千针苋属(AcroglochinSchrad.)、虫实属(Corispermum)、苞藜属(Baolia)组成的分支以低支持率(PP=0.88/BS=64)嵌入藜族(Chenopodieae),且以低支持率(PP=0.58/BS=53)与藜属(ChenopodiumL.)、单性滨藜属(ArchiatriplexG. L. Chu)、刺壤藜属(GrayiaHook. et Arn.)、滨藜属(AtriplexL.)形成的分支(PP=1/BS=100)互为姊妹群。在盐角草族(Salicornieae Dumort.)中,平卧碱蓬(Suaedaprostrata)与碱蓬(Suaedaglauca)、盐角草亚族(Salicorniinae)组成的分支(PP=0.75/BS=88)形成姊妹群关系。樟味藜族(Camphorosmeae Moq.)、猪毛菜族各属以高支持率(PP=1/BS=99)形成一单系群。

后验概率展示在分支上,自展值展示在分支下;对节点年代的测定展示在节点后,同时展示95%置信区间;下同图1 基于ITS序列数据集的Beast分析分子钟系统发育树Posterior probabilities are shown above branches, bootstrap values of the maximum likelihood analysis are shown below branches; Estimated ages of nodes are given behind the respective node, and corresponding bars demonstrate 95% confidence intervals; The same as belowFig.1 Molecular phylogenetic tree from the Beast analysis based on the ITS sequence dataset
2.2 基于atpB-rbcL+rbcL联合序列的系统发育分析
atpB-rbcL+rbcL联合序列矩阵包含物种35个,61条序列。拼接后序列总长度为1 992 bp,其中变异位点有422个(21.2%),保守位点1 570个(78.8%)。Beast分析得到的系统关系树见图2,最大似然树与贝叶斯树拓扑结构大致相同。系统发育树显示,苞藜属(Baolia)、沙蓬属(AgriophyllumM. Bieb.)、合被虫实属(AnthochlamysFenzl)与虫实属(Corispermum)共同构成一较高支持率的单系群(PP=1/BS=93),且苞藜属以高支持率聚为一支(Baolia:PP=1/BS=100)。由上述4属形成的分支在一般支持率下与藜族(Chenopodieae)形成姊妹群关系(PP=0.83/BS=100)。由于没有得到序列信息,千针苋属(Acroglochin)并未包括在此次分析中。与ITS分析不同,盐角草亚族(Salicorniinae)和碱蓬亚族(Suaedinae)形成高支持率(PP=0.99/BS=100)的姊妹群关系。猪毛菜族(Salsoleae)、樟味藜族(Camphorosmeae)以高支持率聚为一单系群(PP=0.99/BS=98)。

图2 Beast分析得到的基于atpB-rbcL+rbcL联合序列数据集的分子钟系统发育树Fig.2 Molecular phylogenetic tree from the Beast analysis based on the concatenated sequence of atpB-rbcL and rbcL
2.3 分化时间估计
表3展示由Beast V1.10.4分子钟定年得到的结果,总结了ITS序列、atpB-rbcL+rbcL联合序列中主要节点的分化年代(均值及95%置信区间)。苞藜属的分化时间在2棵树上分别为17.0和28.7 Ma前。
3 讨 论
3.1 苞藜属与虫实族的单系性
分别基于核ITS序列以及基于叶绿体atpB-rbcL+rbcL联合序列的系统发育树分析显示,苞藜属(Baolia)在atpB-rbcL+rbcL系统发育树中与藜亚科(Chenopodioideae)虫实族(Corispermeae)的沙蓬属(Agriophyllum)、合被虫实属(Anthoch-lamys)、虫实属(Corispermum)互为姊妹群(PP=1/BS=93)。而在ITS系统发育树中苞藜属与虫实属也以较高的支持率聚成一支(PP=0.92/BS=89),表明苞藜属与虫实族具有较近的亲缘关系,这与传统上基于形态学对苞藜属分类地位的划分有很大不同[4, 6-8]。从形态上看,苞藜属与虫实族其他类群均为具有平坦叶的一年生草本,具两性花且花柱二枚。然而苞藜属的独特之处在于:1)虫实族植物常被毛,而苞藜属无毛;2)虫实族植物花序穗状,苞藜属的花则簇生叶腋;3)虫实族植物不具小苞片而苞藜属存在小苞片。从分布来看,虫实族在亚洲、欧洲及北美洲(虫实属)均有分布,在中国分布于东北、华北、西北和青藏高原,而苞藜属为中国特有,仅分布于甘肃西南部的迭部县。

表3 使用Beast程序得到的分歧时间(百万年)评估结果(括号内为95%置信区间)
通过以上分析认为,苞藜属应放在藜亚科(Chenopodioideae),又因其系统地位、形态学与地理分布的特殊性,将其单独放在苞藜族(Baolieae)。
3.2 虫实族的系统位置
ITS和atpB-rbcL+rbcL数据集得到的系统发育树显示,虫实族与藜族(Chenopodieae)关系密切。ITS系统树分析显示,藜族为一并系群,虫实族聚成较高支持率的单系(PP=0.92/BS=89)并嵌入藜族内部。而在atpB-rbcL+rbcL系统树中,虫实族与藜族成员互为姊妹群(PP=0.83/BS=100)。在过去的一些基于叶绿体基因的研究中,也得到了虫实族与藜族成姊妹群关系的证据[28-30]。Berner 等[31]在一项基于核基因ITS的研究中发现,太平洋虫实(CorispermumpacificumMosyakin)与异被滨藜亚族(Suckleyinae G. L. Chu et Stutz)的土荆芥[ChenopodiumambrosioidesL.=Dysphaniaambrosioides(L.) Mosyakin et Clemants]和 异被滨藜[Suckleyasuckleyana(Torr.) Rydb.]形成的分支互为姊妹群,和藜亚族(Chenopodiinae Scott)、轴藜亚族(Axyridinae Heklau)、甜菜族(Beteae Volkens)形成一多枝结构。而Kadereit 等[1]发现虫实族与猪毛菜族(Salsoleae)、盐角草族(Salicornieae)、樟味藜族(Camphorosmeae)形成的分枝聚在一起。从形态上看,虫实族的3个属:虫实属(Corispermum)、合被虫实属(Anthochlamys)和沙蓬属(Agriophyllum),具有平坦的叶、分枝状毛被和穗状花序等。因此,相比于猪毛菜族、樟味藜族、甜菜族和盐角草族,虫实族的形态学特征也与藜族更为接近。
本研究结果支持虫实族作为一个稳定单系群的地位,并且它与藜族有着更为紧密的关系。
3.3 苞藜属的分化时间
基于狭义藜科(Chenopodiaceae)植物属种在现代的分布格局及亲缘关系,朱格麟[13]推测狭义藜科在早白垩纪起源于中国西南部,并且苞藜属(Baolia)是一个原始属。与大多数狭义藜科植物干旱缺水的生境不同,苞藜(Baoliabracteata)的生境相对湿润。研究表明,苞藜属与虫实属(Corispermum)在28.7~17.0 Ma间分化,即晚渐新世至中中新世。从地史上看,该时期喜马拉雅造山运动使造山带东部的夏季降雨量增加,也加重了中国西北部的干旱程度。因此我们推测,苞藜属与虫实属的共同祖先是起源于中国西南部的狭义藜科原始类群的后代,并在晚渐新世至中中新世之间与虫实族其他类群分化,苞藜属适应了湿润的气候,并在第三纪末的气候转冷至第四纪冰期得以幸存,在约3.1~0.6 Ma出现了种内分化,可能是由于冰期时其分布范围收缩引起的地理隔离导致。虫实属、合被虫实属(Anthochlamys)和沙蓬属(Agriophyllum)则适应了干旱环境,在欧亚大陆广泛扩散。其中,虫实属最终可能沿白令陆桥在北美洲登陆[32-33]。由于缺少化石材料,对苞藜的研究还需进一步深入。此外,因苞藜分布域狭小且生境受到人为干扰,故在此呼吁尽快对其开展(异地)保护工作,加强物种生物学研究。
致谢:感谢任广朋、苏敏在数据分析、实验操作中提供的帮助。
