额济纳湿地多枝柽柳种群的遗传多样性与遗传结构
2021-09-04张雅楠贾媛媛李佳彬党振华
张雅楠,黄 蕾,张 雷,贾媛媛,李佳彬,党振华*
(1 内蒙古大学 生态与环境学院, 蒙古高原生态与资源利用教育部重点实验室/内蒙古草地生态学重点实验室, 呼和浩特 010021; 2 内蒙古自治区林业科学研究院, 呼和浩特 010010; 3 内蒙古大青山森林生态系统定位观测研究站, 呼和浩特 010010)
遗传多样性和遗传结构是评价特定物种及其种群生存现状的重要指标。前者是任何物种或群体应对外界环境压力并得以延续的内在基础,而遗传结构则是生物适应、物种形成及进化的表现[1]。评价物种遗传多样性,剖析群体遗传结构,是适应进化研究的两个重要内容,对种质资源的保护与开发具有重要的理论意义和现实价值[2]。DNA分子标记因数量多、含有的变异信息丰富、受环境因素影响小,成为群体遗传学研究的有效工具[3]。SSR(simple sequence repeat)又称微卫星DNA(microsatellite DNA),通常是由1~6个核苷酸串联重复成几十个核苷酸的DNA序列,具有在真核生物基因组中分布广泛、多态性高、共显性等特点[4-5],是最常用的分子标记技术之一,被广泛应用于玉米(Zeamays)[6-7]、烟草(Nicotianatabacum)[8-9]、狗尾草(Setariaviridis)[10]、甜菜(Betavulgaris)[11-12]、丹参(Salviamiltiorrhiza)[13-14]等植物的种群遗传、起源与进化、种质资源保护等研究。EST-SSR(express sequence tag-SSR)是位于基因转录序列中的微卫星DNA,因其具有易开发、多态性丰富、种间通用性好、对特定基因的功能有潜在影响等特性,在遗传图谱构建、种质特性鉴定、目标性状分子标记筛选、功能基因组等领域中均有应用。得益于高通量测序技术的飞速发展,大量研究利用转录组测序技术在非模式物种中进行了EST-SSR的识别、特性分析及标记开发,极大推动了该标记自身特性的研究及其在相关研究中的应用。吕冰冰等[15-16]利用EST-SSR标记,并结合SSR标记和SRAP标记构建了小麦(Triticumaestivum)遗传连锁图谱,丰富了小麦功能分子标记的数量;Ranathunge等[17]报道了向日葵(Helianthusannuus)EST-SSR的变异与不同种群特定基因的差异表达相关;在种群遗传学研究中发现,该标记的引物扩增成功率、等位基因数目、多态性及种间通用性均与基因组SSR相似,并能客观评价物种的遗传多样性,表现出EST-SSR标记在该方面研究中的巨大潜力[18-19]。
柽柳(Tamarix)是起源于第三纪的古老植物,全球约有90个种,主要分布于欧洲、亚洲和非洲的干旱、半干旱草原及盐碱地带[20]。中国的柽柳资源集中分布在华北、东北和华东等地的沙荒地、干旱盐碱地及盐碱湿地,常见于草地、滩涂、河谷及海滨沙地。该属植物具有抗旱、抗风沙、抗病虫害、抗盐碱和耐水湿的能力,常形成大面积柽柳纯林,具有极强的生态适应性,在盐碱地造林绿化方面具有广泛的应用前景,是营造水土保持林、沿海滩涂及河岸防护林的优良树种[21]。此外,柽柳易于培育薪炭林,可用作园林景观植物;其细嫩枝叶可入药,荒漠区柽柳根部常有野生管花肉苁蓉寄生,具有一定的药用和经济价值[22]。目前,国内外学者主要从生物学特征[23-24]、种群生态学[25-26]和抗逆机理[27-28]等方面对柽柳开展了研究。随着DNA标记技术的普及,不同柽柳植物的群体遗传多样性和遗传结构研究也被陆续报道。杨晴等[29]采用ISSR标记调查了河北省秦皇岛市柽柳种群的遗传多样性和亲缘关系,认为该地区柽柳具有较高遗传多样性;温月仙等[30]利用叶绿体和核基因序列对黄河流域甘蒙柽柳(Tamarixaustromongolica)开展了种群遗传学研究,检测到其叶绿体基因的遗传多样性低于核基因,遗传变异主要发生在种群内,无明显谱系地理结构;苏志豪等[31]利用7个SSR标记鉴定出塔里木盆地的沙生柽柳(Tamarixtaklamakanensis)具有一定的遗传多样性,但种群的平均等位基因多样性较低。可见,不同柽柳植物的群体遗传特征因所处生境的气候、水分、土壤等条件不同而表现出较大差异。
多枝柽柳(Tamarixramosissima)属多年生灌木或小乔木,在柽柳属植物的所有生境类型中均有分布,如荒漠、半荒漠、草原和湿地等,是西北干旱、半干旱区分布最广泛的柽柳属植物[26,32]。在潮湿河岸地带,该植物的水分竞争能力和泌盐能力极强,对草本植物的生长具有限制作用,易形成单优势种植物群落;在荒漠区,多枝柽柳通过根系的“提水作用”将水分从深层土壤和地下水输送到浅层土壤,水分和养分富集形成“沃岛效应”,间接为荒漠区动植物提供了适宜栖息地,在生态系统和植物种质资源多样性维持上均具有重要价值[33]。由于气候变化、过度开垦和开发、过度放牧等自然和人类活动干扰,中国西北地区环境不断恶化、荒漠化加剧。黑河下游的额济纳绿洲是西北干旱区的主要天然绿洲,代表性植物群落有胡杨(Populuseuphratica)和柽柳(Tamarixspp.)等。其中,以柽柳及其为主的群落所占面积最大(70%以上),尤以多枝柽柳(T.ramosissima)最多。20世纪60年代以来,进入黑河下游的水量逐渐减少,导致了河流和湿地面积大量缩减,使额济纳绿洲荒漠化的进程显著加快,威胁着大量植物的生存和繁衍[34]。李森等[35]发现,人为活动影响是额济纳绿洲荒漠化的主导因素,如对黑河的过度开发和绿洲区的“三滥”活动。不幸的是,额济纳柽柳群落赖以生存的水资源均源于黑河及其补给的地下水,当供水量不足时,其生境便开始衰退,演变为裸地和流沙,最终荒漠化[36]。可见,柽柳群落的生存状况关系到额济纳绿洲生态系统的稳定,是评价其健康与否的重要指标。本研究以额济纳低海拔湿地的12个天然多枝柽柳种群为研究对象,利用EST-SSR标记技术分析其遗传多样性和遗传结构,旨在探明该地区多枝柽柳种质资源的基本特征,为该区域的生态与环境保护提供科学依据。
1 材料和方法
1.1 样品采集
样本于2019年8月采自内蒙古自治区阿拉善盟额济纳旗,选取12个野生多枝柽柳种群,每个种群采集24个长势良好、无病虫害个体的幼嫩叶片。采样时,每个个体间隔5~10 m,采样后置于变色硅胶中保存,带回实验室备用。采样位置的详细地理信息见表1和图1。
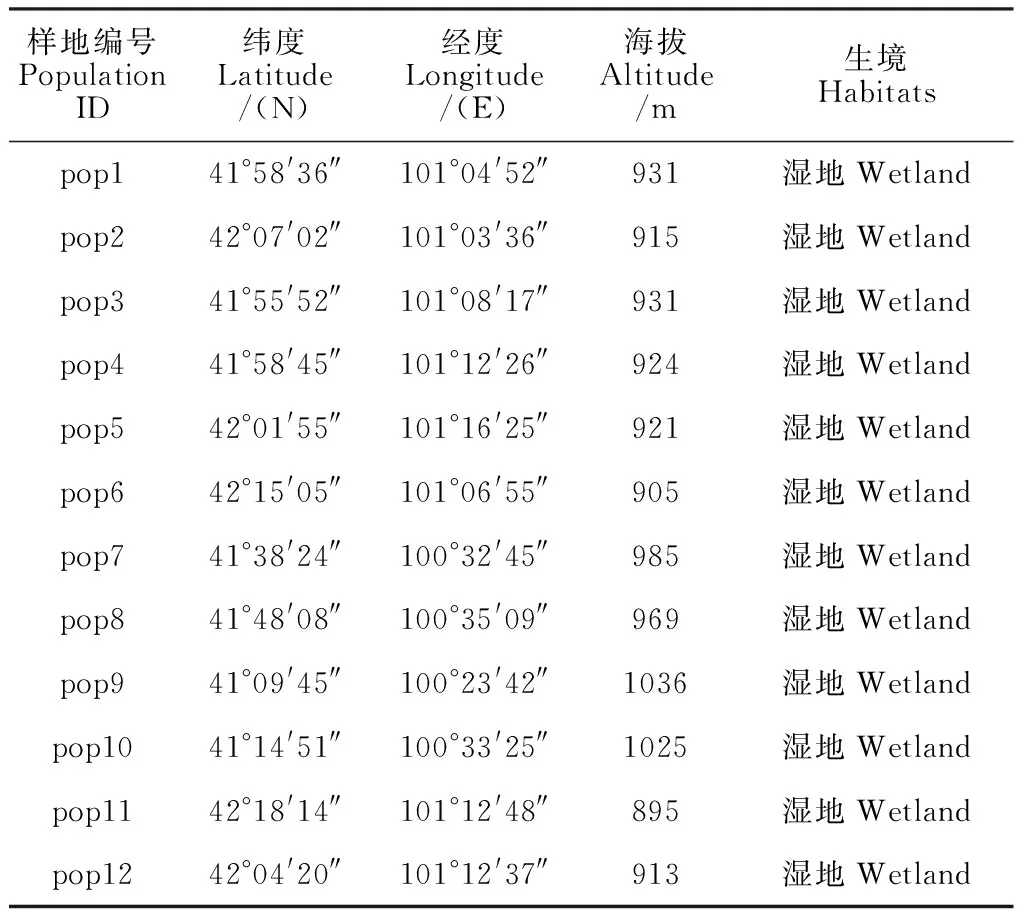
表1 多枝柽柳采样点信息

图1 多枝柽柳采样点地理分布图Fig.1 Geographic distribution of T. ramosissima
1.2 SSR标记的多态性检测
使用植物基因组DNA提取试剂盒(DP305,天根生化科技有限公司,北京)提取多枝柽柳基因组DNA,利用1%琼脂糖凝胶电泳和超微量紫外分光光度计(Nano Drop 2000c,赛默飞世尔科技有限公司,马萨诸塞州,美国)进行DNA质量检测,最终获得191个合格的DNA样本,储存于-20 ℃备用。
本研究所用SSR分子标记为本实验室前期开发所得的14个EST-SSR标记(表2)。使用前在每个标记的上游引物5′端加FAM、HEX或ROX荧光标记,用于后期毛细管电泳检测。以多枝柽柳DNA为模板,在ABI2720 PCR扩增仪中采用25 μL混合反应体系进行PCR扩增。PCR反应体系为:1 μL模板DNA(50 ng/μL),12.5 μL Premix Taq(宝生物工程有限公司,大连),0.5 μL上游引物,0.5 μL下游引物和10.5 μL ddH2O。PCR扩增程序为:94 ℃ 5 min;94 ℃ 30 s,引物最佳退火温度(50~60 ℃)30 s,72 ℃ 30 s,35个循环;72 ℃ 10 min终止反应。扩增产物经1%琼脂糖凝胶电泳检测后,委托擎科生物科技有限公司(北京)在内标为GS500LIZ的ABI 3730 DNA分析仪(赛默飞世尔科技有限公司,马萨诸塞州,美国)上进行毛细管电泳检测。
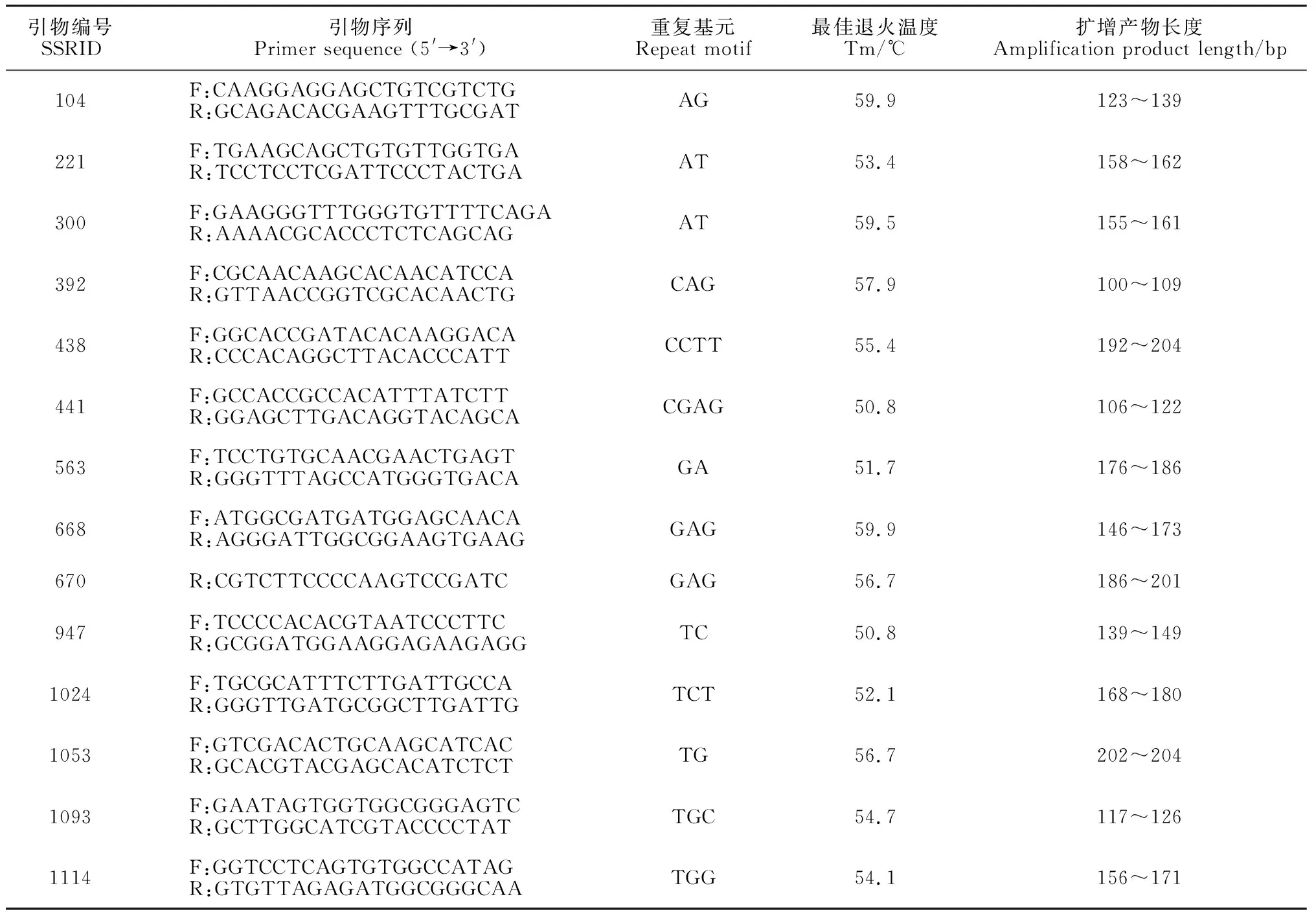
表2 14个多态EST-SSR标记的基本信息
1.3 数据处理
使用GeneMarker V2.6.0(SoftGenetics,宾夕法尼亚州立大学,美国)对毛细管电泳结果进行矫正和等位基因判读。用PowerMarker V3.0(http://www.powermarker.net/)分析各SSR的多态性信息含量(PIC);利用GenAlEx V6.502(http://biology.anu.edu.au/GenAlEx)计算等位基因数(Na)、期望杂合度(He)、观测杂合度(Ho)、Shannon’s信息指数(I)、种群间多态位点百分率(P)、分子方差分析(AMOVA)、遗传分化系数(Fst)、近交系数(Fis)和基因流(Nm);利用PopGene V1.32(http://www.ualberta.ca/~fyeh/)统计Nei’s遗传距离和遗传相似性系数。采用R包ecodist进行Mantel检验,分析群体遗传距离与地理距离的相关性。
使用Structure V2.3(http://pritch.bsd.uchicago.edu/structure.html)分析多枝柽柳的群体遗传结构。参数设置为:K为2~12,“Length of burnin period”为10 000,“Number of MCMC replications after burnin”为100 000,每个K值单独运行20次。结果上传至Structure Harvester(http://taylor0.biology.ucla.edu/structure Harvester/)计算最佳K值。基于PopGene V1.32得到的群体间遗传距离,利用FAMD V1.31(http://www.opensource.org/licenses/artistic-license-2.0.php)和MEGA V10.0.4(http://www.megasoftware.net)进行PCoA分析和非加权配对法(UPGMA)聚类分析,并采用最大似然法构建进化树。
2 结果与分析
2.1 EST-SSR标记的遗传多样性指数
14个EST-SSR标记在191个多枝柽柳个体中共扩增出79个等位基因(Na),每个基因座上的等位基因数为3~7个(平均值为5.6),期望杂合度(He)在0.18~0.77间(平均值为0.55),观测杂合度(Ho)在0.05~0.51范围内(平均值为0.29),多态信息含量(PIC)在0.17~0.74间,平均值为0.51(表3)。

表3 14个多态EST-SSR标记的遗传参数
2.2 多枝柽柳种群的遗传多样性
各多枝柽柳种群的多态位点比率(P)平均值为0.970,期望杂合度(He)在0.368~0.565间(平均0.473),观察杂合度(Ho)在0.224~0.383间(平均0.294),Shannon’s指数(I)在0.680~1.037间(平均0.862),固定指数(F)在0.182~0.443间(平均0.332)。其中,pop2、pop3、pop5、pop6和pop12的Ne和He均较高,具有相对丰富的遗传多样性,而其余种群的Ne和He分别在1.830~2.160和0.368~0.470间,遗传多样性相对较低。各种群He、Ho以及I等遗传参数变化不大,遗传多样性相似(表4)。
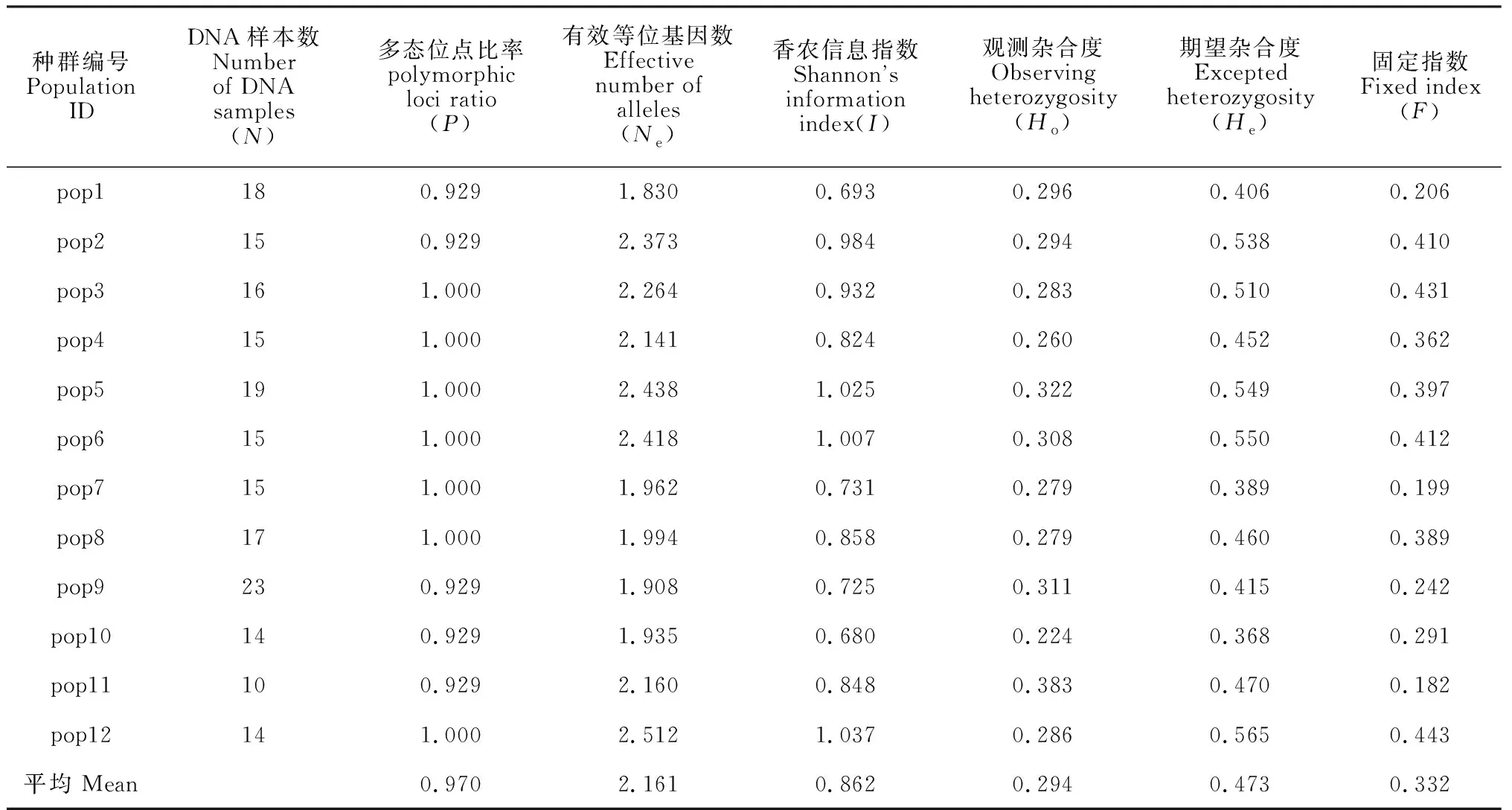
表4 12个多枝柽柳种群的遗传多样性
多枝柽柳种群内的近交系数(Fis)为0.366,群体间平均基因流(Nm)为4.425,种群间平均遗传分化系数(Fst)为0.079,各种群间的遗传分化系数(Fst)见表5。AMOVA分析结果显示,种群间遗传变异为14.0%,种群内遗传变异为86.0%,与上述种群遗传分化系数结果吻合(表6)。
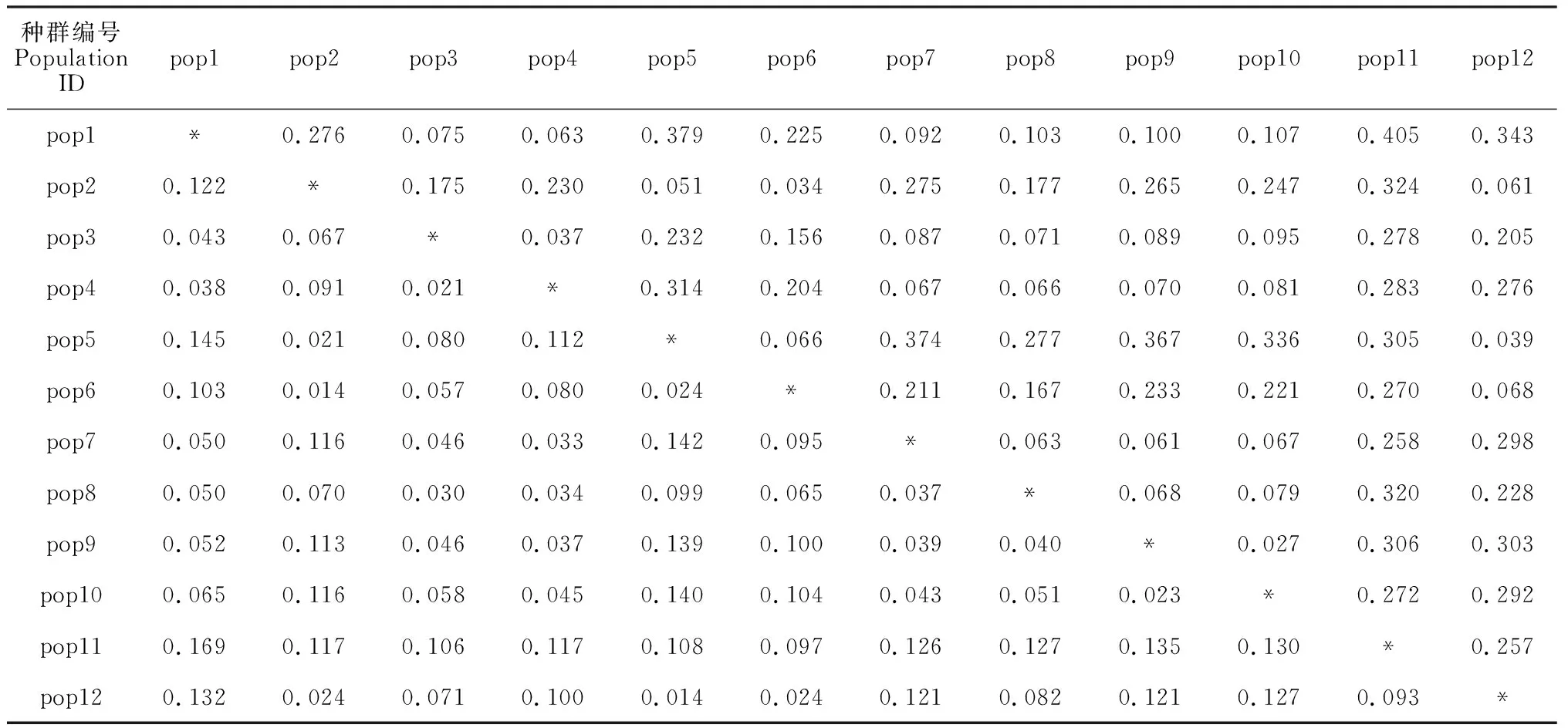
表5 多枝柽柳种群间的遗传分化系数(Fst)及Nei’s遗传距离

表6 多枝柽柳种群间和种群内分子变异的AMOVA分析
遗传距离分析显示(表5), pop1、pop3和pop4的遗传距离在0.063~0.075间; pop2、pop5、pop6和pop12的遗传距离在0.034~0.061内;pop7、pop8、pop9和pop10的遗传距离在0.061~0.067间,而pop11与其他种群间的遗传距离相对较远。
2.3 多枝柽柳的种群遗传结构
当K为4时,ΔK最大(图2,A),说明191份多枝柽柳可分为4个亚群:即pop1、pop3和pop4为Ⅰ组,pop7、pop8、pop9和pop10为Ⅱ组,pop2、pop5、pop6和pop12为Ⅲ组、pop11为Ⅳ组(图2,B)。
为进一步分析多枝柽柳种群间的遗传关系,利用12个种群间的Nei’s 遗传距离进行了UPGMA聚类分析(图3,A)和主坐标分析(PCoA)(图3,B)。结果显示,这些多枝柽柳种群均被分为4组:pop1、pop3和pop4为一组,pop2、pop5、pop6和pop12为一组,pop7、pop8、pop9和pop10为一组,pop11为一组,与遗传结构分析结果一致。Mantel检验表明各种群的遗传距离和地理距离不具相关性(r=0.194,P=0.120,图4)。
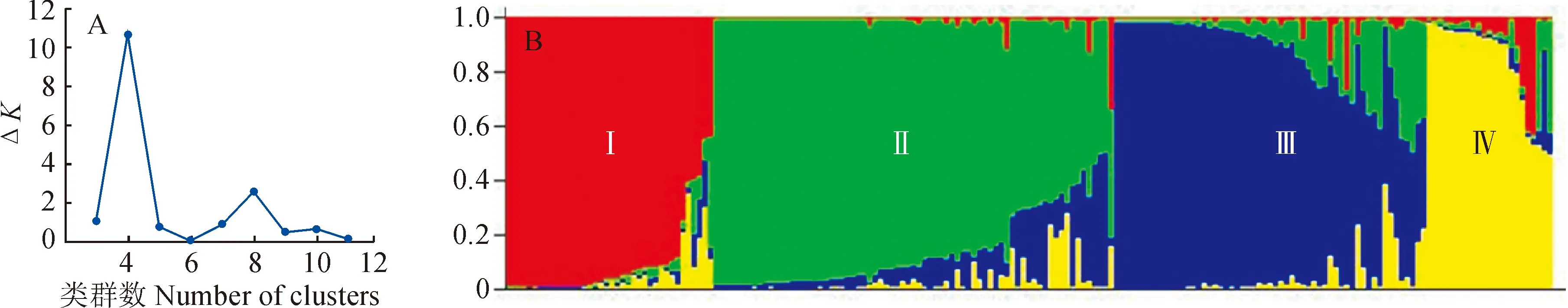
红色代表pop2、pop5、pop6和pop12;绿色代表pop7、pop8、pop9和pop10;蓝色代表pop1、pop3和pop4;黄色代表pop11图2 多枝柽柳亚类群数确定(A)及遗传结构分析(B)Red represents pop2, pop5, pop6 and pop12; green represents pop7, pop8, pop9 and pop10; blue represents pop1, pop3 and pop4; yellow represents pop11Fig.2 Subpopulation identifying (A) and population genetic structure analysis (B) of T. ramosissima
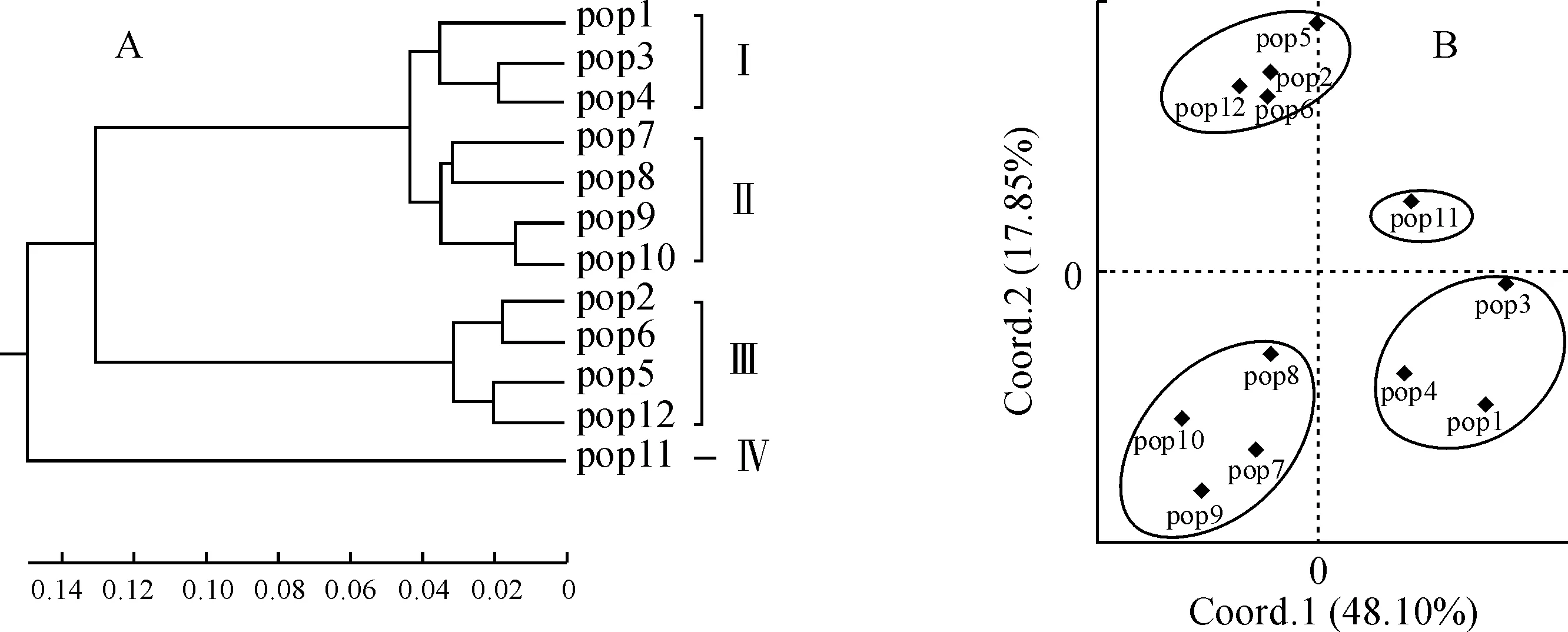
比例尺对应的是基于最大似然法的进化距离图3 基于遗传距离的多枝柽柳种群的UPGMA聚类分析(A)和主坐标分析(B)The scale bar corresponds to the evolutionary distance based on the Maximum Likelihood MethodFig.3 UPGMA dendrogram (A) and Principal Coordinates analysis (B) based on genetic distance of T. ramosissima populations
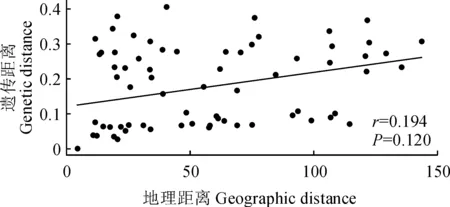
图4 多枝柽柳种群的遗传距离与地理距离的相关性分析Fig. 4 Correlation analysis between genetic distance and geographic distance of T. ramosissima populations
3 讨 论
遗传多样性是衡量一个物种适应环境变化的重要指标,是物种生存和进化的内在基础[37]。物种的适应性与遗传多样性密切相关,即遗传多样性越高,适应性越强[38-39]。本研究所用的14个SSR标记虽位于多枝柽柳基因序列内,但表现出中高度的多态性(平均He和平均Ho分别为0.55和0.29,平均PIC为0.51),一定程度体现了这些种群遗传变异信息丰富的特性。一般来说,一组等位基因的多态性百分率超过50%则说明它们具有相对丰富的遗传多样性[40-41]。蒋志敏等[42]利用ISSR标记技术对黄河三角洲3个柽柳种群的90个个体进行了遗传多样性分析,发现10个标记的多态性比率为85%;赵景奎等[43]利用RAPD标记技术分析了黄河三角洲3个天然柽柳群体的遗传多样性和遗传分化,得到40.07%的多态百分比;叶春秀等[44]通过SSR标记分析了新疆塔河流域上、中、下游柽柳的种质资源遗传多样性,发现它们的多态性比率为79.49%。本研究发现,12个种群的平均多态位点比率为97.0%,表明额济纳湿地多枝柽柳种群具有丰富的遗传变异。综合Ne、He等种群遗传参数来看,pop2、pop3、pop5、pop6和pop12的遗传多样性相对较高(He均大于0.5),但所有种群的遗传多样性差别不大,暗示了它们中等水平遗传多样性的特征。
影响种群遗传结构的因素有繁育系统、地理分布、基因流等,其中基因流是种群遗传结构均质化的关键因素,能有效减少种群间的遗传变异[45]。本研究发现,多枝柽柳群体间的平均基因流(Nm)为4.425,说明所调查种群间存在频繁的基因交流,能有效减少遗传漂变对这些种群的影响,避免了群体间的遗传分化。固定指数(F)是衡量杂合子缺失或过剩的参数[46]。12个多枝柽柳种群的F值均大于0,表明它们处于杂合子缺失状态,存在一定程度的近交现象[47]。当然,这可能与本研究调查群体的地理距离相对较近有关。张娟等[38]对9个地理距离相对较远的新疆天然刚毛柽柳(Tamarixhispida)种群研究发现,可能因刚毛柽柳传粉媒介(风媒和虫媒等)的传播距离小,造成了采样群体间的基因流障碍,故而检测到群体间较高的分化程度。有趣的是,虽然基因流广泛,但可能与其传粉媒介的传粉能力和繁殖方式有关,多枝柽柳种群内的平均近交系数亦较高(Fis=0.366)。王仲礼等[39]和Bawa等[48]发现,柽柳的传粉媒介是以小型蝇类和蜂类为主,兼有多种昆虫传粉;王仲礼等[39]和张娟等[38]发现柽柳的繁育方式为自交亲和,但缺乏自花传粉机制,不存在无融合生殖现象,为兼性异交。因此可推测,如果出现地理距离隔离或长期缺乏长距离传粉者,会导致多枝柽柳遗传多样性下降,出现近交衰退现象。
群体遗传结构是遗传变异在群体内、群体间的分布以及在时间上的变化,研究群体遗传结构有助于种质资源的保护、利用和改良[49-50]。本研究发现,12个多枝柽柳种群可划分为4个亚群。一般来说,长寿和异交物种的遗传变异主要发生在种群内,而自交物种的遗传变异多发生在种群间[51]。12个多枝柽柳种群的平均遗传分化系数(Fst)为0.079,说明在总遗传变异中,7.9%来自种群间,92.1%来自种群内,与AMOVA分析结果相似(14.0%-vs-86.0%)。这与其他柽柳科植物种群间的遗传分化情况一致,如长叶红砂(Reaumuriatrigyna)(Gst=0.1425)[45]、红砂(Reaumuriasoongorica)(Gst=0.1931)[52]、疏花水柏枝(Myricarialaxiflora)(Gst=0.224)[53]等。蒋志敏[42]、赵景奎等[43]和叶春秀等[44]同样发现柽柳的遗传变异主要发生于种群内部,而种群间遗传分化程度相对较低。
多枝柽柳以耐干旱、耐盐碱、繁殖迅速等特点,被视为阿拉善荒漠区的重要造林绿化树种,是延缓绿洲荒漠化的重要防线,保持较高的遗传多样性是该植物遗传资源保护与开发利用的核心。本研究发现,尽管所调查的多枝柽柳种群具有较丰富的遗传多样性,但它们均呈现出一定程度的杂合子不足及近交现象,对该地区多枝柽柳遗传多样性的长期维持构成了潜在不利影响。因此,在开发与利用该种质资源的同时,应考虑维持和创造多枝柽柳群体间的基因流动,尽量减少人类活动对其生存环境的干扰和破坏,保护和修复已受损的湿地生态系统。
