秋季施肥对毛白杨苗木质量、造林效果和养分回流的影响*
2021-09-04王苗苗李国雷彭玉信刘春和赵建松王书红王长伟赵蕊蕊
王苗苗 刘 勇 李国雷,2 彭玉信 刘春和 赵建松 王书红 蕫 彪 王长伟 赵蕊蕊
(1.北京林业大学省部共建森林培育与保护教育部重点实验室 北京 100083;2.城乡生态环境北京实验室 北京 100083;3.北京市黄垡苗圃 北京 102601)
苗木质量是决定造林成功与否的首要条件(Cuestaetal.,2010;Villar-Salvadoretal.,2015),培育大规格且养分(主要是氮)富足的苗木有利于提高造林成活率和生长表现(Olietetal.,2009;2013;Villar-Salvadoretal.,2012)。在苗圃阶段的苗木培育过程中,当苗木进入硬化期(生长后期)后,生物量继续增长,若此时停止施肥,苗木从土壤中获取的养分减少,会导致苗木体内养分稀释(Ritchieetal.,1980),从而降低苗木质量。研究发现,秋季施肥能够有效提高养分贮存(李国雷等,2011;魏红旭等,2011),是一种理想的稳态养分加载方式(Villar-Salvadoretal.,2012;2015),且合理的秋季施肥方案(顶芽形成后开始施肥,施用氮磷钾全肥,肥料溶液浓度小,施肥4~6周)还可提高苗木抗寒性(Dumroese,2003;Floistadetal.,2004;Islametal.,2009;李国雷等,2011;Lietal.,2012)。目前,苗木秋季施肥方案的研究主要集中于常绿树种(Birchleretal.,2001;Boivinetal.,2002;2004;Southetal.,2002;Rikalaetal.,2004;Islametal.,2009;Andiviaetal.,2011;Olietetal.,2011;刘洲鸿等,2002;李珊等,2018),而对落叶树种研究较少(刘勇等,2000;Lietal.,2012;2014;Zhuetal.,2013;Wangetal.,2016)。Li等(2012;2014)研究苗木硬化期不同水平氮处理得出,秋季施肥可显著提高长白落叶松(Larixolgensis)和栓皮栎(Quercusvariabilis)苗木的根生物量积累和氮贮存。王佳茜(2019)通过同位素标记(15N)发现,栓皮栎苗木移栽初期,新组织生长所需氮的78%~98%来源于硬化期的氮贮存,且贮存量越高,对翌年新生的贡献越大。Wang等(2016)研究认为,中、高施肥量对栓皮栎苗木移栽后的生长、养分贮存和吸收具有促进作用。然而,Andivia等(2011)却认为,秋季施肥对冬青栎(Quercusilexspp.ballota)造林成活率和生长无影响。因此,有必要将苗木质量与造林效果相结合,进一步探究秋季施肥对落叶树种的影响。
苗木生长发育所需养分除从外界获取外,也可通过自身养分回流(nutrient resorption,NuR)获得。养分回流一般指叶片衰落时,将氮、磷等回流至其他组织,以满足生长、发育和繁殖的养分需求,通常用养分回流效率(nutrient resorption efficiency,NuRE)表征(Aerts,1996;Brantetal.,2015;Yuanetal.,2015)。Vergutz等(2012)研究得出,陆地生态系统植物叶片氮、磷和钾的回流率高达62.1%、64.9%和70.1%。植物通过养分回流,既可减少凋落物带来的养分流失,提高竞争力,还可降低对土壤养分的依赖,是一种重要的养分维持策略(nutrient conservation strategy)(Aertsetal.,2000;Vergutzetal.,2012;Tangetal.,2013)。植物生长速度不同,对养分需求不同(翟明普等,2016),养分回流也可能存在差异。Nambiar等(1991)研究发现,不同肥力条件下新西兰辐射松(Pinusradiata)的生长速率与养分回流呈正相关;而Harvey等(1999)则认为,不同施肥与干旱处理下杂交杨(Populus)养分回流的变化与生长速率无关。秋季施肥通过提高苗木养分贮存,可能促进翌年生长,然而,这种因苗圃施肥带来的翌年生长差异是否影响其养分回流尚未可知。
毛白杨(Populustomentosa)是我国北方特有的乡土树种,也是我国华北地区平原绿化、绿色通道和用材林基地建设的主要树种(李吉跃等,2012)。三倍体毛白杨(triploidP.tomentosa)是朱之悌等(1998;2002)采用细胞染色体部分替换和染色体加倍等技术对毛白杨进行遗传改良获得的新品种,与自然二倍体相比,其具有速生、优质、抗病、材质白、纤维长等特点,被广泛应用于造林、工业制浆和造纸等,并已在华北地区大面积栽培(张平冬等,2011;宋曰钦等,2015)。本研究以三倍体毛白杨苗木为试验材料,探究秋季施肥对苗木质量、养分贮存和造林效果的影响,明确苗木生长与养分回流的内在联系,确定最佳施肥水平,以期为苗木培育精准施肥提供参考。
1 材料与方法
1.1 试验地概况与试验材料
试验地位于山东省聊城市冠县毛白杨国有苗圃(36°30′N,115°22′E,海拔37 m),属温带季风区域大陆性半干旱气候,光照充足,四季分明。年均气温12.8~13.3 ℃,年均降水量549.9~600 mm,无霜期198~227天。土壤为砂壤土,平均pH7.8,养分状况见表1(依据毛白杨根系分布特征分层取样)。供试材料为当年5月炼苗出圃的三倍体毛白杨组培苗“北林雄株1号”(P.tomentosa“Beilinxiongzhu 1”),由秋水仙碱溶液诱导银腺杨(P.alba×P.glandulosa)2n花粉与毛新杨(P.tomentosa×P.bolleana)杂交获得,并利用组培快繁技术无性快速扩繁(康向阳等,2000;王沛琦等,2014)。

表1 苗圃地和造林地土壤养分状况Tab.1 Soil nutrient status at nursery and field
2017年5月20日进行组培苗炼苗,1周后选择长势相近的幼苗[苗高H:(11.3±0.193)cm,地径D:(1.91±0.030)mm,数量n=30]移栽至露天苗圃地。6月7日(生长1周后)施用精准奥绿肥(Scotts Co.,Marysville,OH,USA)(15%N + 9%P2O5+ 12%K2O + TE,21 ℃释放期5~6个月),每株60 g,即纯氮每株9 g(曹帮华等,2004)。采用两侧穴施法单株施肥,穴深15 cm,距幼苗10 cm。育苗期间除草和浇水按需进行,保证不影响苗木生长。共育苗1 600株,株行距0.3 m×1.0 m。
1.2 试验设计
1.2.1 苗圃阶段 采用单因素完全随机区组设计,设置3个秋季施肥水平和1个对照,每株施肥0、5、10和20 g。选用水溶肥花多多1号(Peters®Professional,Scotts Co.,Marysville,OH,USA)(20%N + 20%P2O5+ 20%K2O + TE),浓度为41.6、83.3和166.6 g·L-1;对照施用等量自来水,每株20 mL。自9月2日起每周施肥1次,连续6周平均施入,每次施肥2天后浇水。试验共4个处理、4个区组,每小区100株苗(10行×10列,边缘的2行×2列为保护行)。
1.2.2 造林阶段 2018年3月下旬,每小区随机选取15株长势相近的苗木移栽至造林地。株行距3 m×3 m,3行×5列,完全随机排列。造林定植后立即浇1次透水,1周后浇第2次透水。生长季对苗木人工除草3次。
1.3 样品采集与分析
1.3.1 苗圃阶段苗木质量评价 秋季施肥前(9月1日),每小区选取5株标准苗木测定初始苗高(H0)、地径(D0),标准苗木指小区内中间6行×6列随机选择的健康苗木;生长季末(11月23日),再次测定苗高(H1)、地径(D1)。11月24日破坏取样,每小区选取5株苗木(因根系庞大,并未取得完整根系,仅根据造林移栽需求取得40 cm×40 cm×40 cm的主根系)。根系洗去基质和土壤,分离茎、根,65 ℃烘48 h,称茎生物量。按照组织部位将同一处理各小区的 5 株苗木样品混合,粉碎,过60目筛。称取样品0.2 g,采用浓H2SO4-H2O2法消化,凯氏定氮法测定全氮(UDK-152 全自动定氮仪,VELP Scientifica,Italy),钼锑抗比色法测定全磷(8453 型紫外分光光度计,Agilent 公司,USA),火焰光度计法测定全钾(SpectrAA220 型原子吸收光谱仪VarianInc.,USA)(Lowther,1980)。称取 0.1 g 混合干样,采用蒽酮比色法(8453 型紫外分光光度计,Agilent 公司,USA)测定可溶性糖和淀粉浓度。
1.3.2 造林后苗木生长评价 苗木定植后(3月30日),每小区选取中间列5株苗木作为标准苗木测定苗高(H2)、地径(D2);生长季末(11月24日),再次测定苗高(H3)、地径(D3)。茎体积(stem volume,SV,cm3)=1/3×3.14×(D/2)2×H。并计算造林成活率。
1.3.3 造林后苗木养分回流 根据2017年苗圃阶段的表现,选取施肥量每株10 g处理的苗木与对照比较,探究秋季施肥对造林后苗木养分回流的影响。生长盛期(8月20日),每小区选取中间列5株苗木向阳面中部摘取40片全展叶,混合;生长季末(11月23日),收集表层落叶。全部叶样65 ℃烘48 h,粉碎,过筛。氮、磷、钾测定方法同上。
养分回流效率(NuRE,%)=(Nugre-Nusen)/
Nugre×MLCF×100。
式中:Nugre为绿叶中养分浓度;Nusen为落叶中养分浓度;MLCF(mass loss correction factor)为生物量损失校正因子,值为0.784(Vergutzetal.,2012)。
1.4 数据统计与分析
采用 Excel2016整理数据,利用SPSS19.0(IBM,Chicago,USA)的“探索”和“转换”功能对数据进行检验和转换,确保数据满足正态性和同质性。通过单因素方差分析探究秋季施肥对苗高、地径、茎生物量以及氮、磷、钾、可溶性糖、淀粉浓度和养分回流效率的影响。运用广义二项分布非线性模型分析秋季施肥对苗木造林成活率的影响。如处理间差异显著,则基于最小二乘法(LSD)在 0.05 水平上进行多重比较。应用 SigmaPlot12.5(Systat Software,San José,CA,USA)作图,并分别建立造林后苗木生长与养分回流效率间的线性回归方程。
2 结果与分析
2.1 苗圃阶段苗木生长、养分贮存和非结构性碳水化合物水平
秋季施肥对苗圃阶段苗木生长的影响较小(表2)。与对照相比,秋季施肥后苗高、地径和茎生物量分别提高11.4%~20.6%、1.5%~10.5%和1.8%~6.4%。苗高随施肥量增加而增加,但地径和茎生物量在施肥量每株20 g时呈下降趋势。

表2 秋季施肥对苗圃阶段和造林后苗高、地径和茎生物量(茎体积)的影响Tab.2 F and P values for the effect of fall fertilization on seedling height,diameter and stem biomass(stem volume)increment at nursery and field
秋季同时施用氮、磷、钾肥,主要影响苗木氮贮存,而对磷、钾贮存的影响差异不显著(图1、表3)。随施肥量增加,苗木茎、根中的氮浓度呈先上升后下降的趋势,施肥量每株10 g时达到最大值。与对照相比,秋季施肥后苗木茎、根中的氮浓度分别提高2.4%~12.0%和17.4%~48.1%。

图1 秋季施肥对苗圃阶段苗木茎、根中氮浓度的影响Fig.1 The effect of fall fertilization on N concentration of stem and root at nursery
秋季施肥对非结构性碳水化合物水平的影响差异不显著(表3)。非结构性碳水化合物主要贮存在根中,可溶性糖和淀粉浓度是茎中的2倍。与对照相比,秋季施肥后根淀粉浓度增加17.9%~34.5%(表4)。

表3 秋季施肥对苗圃阶段苗木茎、根中养分浓度和非结构性碳水化合物水平的影响Tab.3 F and P value for the effect of fall fertilization on nutrient and non-structural carbohydrate concentrations in stem and root at nursery

表4 秋季施肥对苗圃阶段苗木茎、根中非结构性碳水化合物的影响Tab.4 The effect of fall fertilization on non-structural carbohydrate in stem and root at nursery
2.2 造林效果和养分回流
秋季施肥显著影响苗木造林成活率和生长表现(图2、表2)。随施肥量增加,苗高、地径和茎体积生长量整体表现为先上升后下降的趋势。与对照相比,施肥量每株10 g时苗木生长量达到最大(苗高、地径和茎体积生长量分别提高40.0%、80.4%和85.3%),而施肥量每株20 g时表现出抑制作用。与对照相比,秋季施肥对苗木造林成活率影响不显著(P>0.05),当施肥量每株20 g时苗木成活率最低(63.3%)。

图2 秋季施肥对造林后苗高、地径、茎体积生长量和成活率的影响Fig.2 The effect of fall fertilization on seedling height,diameter and stem volume increments,as well as survival rate at field 图中数值为均值 ± 标准误。不同小写字母代表不同处理间差异显著(LSD检验,P<0.05,n=4)。下同。Values are mean±SE.Different lowercase letters indicate significant difference among treatments(LSD test,P<0.05,n=4).The same below.
秋季施肥显著影响绿叶磷(F=109,P<0.001)和落叶氮(F=408,P<0.001)、磷(F=60,P<0.001)、钾(F=36.6,P=0.001)的浓度(图3)。与对照相比,秋季施肥后苗木落叶氮和钾的浓度分别下降56.4% 和37.3%,而绿叶磷和落叶磷分别提高19.7%和21.6%。秋季施肥使苗木氮回流效率显著提高71%,而对磷回流效率无显著影响。

图3 秋季施肥对造林后苗木绿叶和落叶中养分浓度及养分回流效率的影响Fig.3 The effect of fall fertilization on nutrient concentration in green and senesced leaves as well as nutrient resorption efficiency at field图中数值为均值 ± 标准误。不同小写字母代表秋季施肥对绿叶养分浓度的显著影响,“*”代表秋季施肥对落叶养分浓度的显著影响。椭圆中的数值为养分回流效率,不同大写字母代表秋季施肥对养分回流的影响差异显著(LSD检验,P<0.05,n=4)。Values are mean±SE.Different lowercase letters indicate significant difference of nutrients in green leaves,“*”indicate difference of nutrients in senesced leaves.Values in oval indicate nutrient resorption efficiency and different capital letters indicate difference of nutrient resorption efficiency with or without fall fertilization(LSD test,P<0.05,n=4).
由造林后茎体积生长量与养分回流间的线性回归方程可知,氮回流效率与茎体积生长量呈显著正相关,而磷回流效率与茎体积生长量无关(图4)。此外,养分回流效率的变化并非受单一元素控制,与叶片中多种元素浓度变化相关(表5)。氮回流效率与落叶氮和落叶钾浓度呈负相关,与绿叶磷、落叶磷浓度呈正相关,磷回流效率与绿叶氮浓度呈正相关。
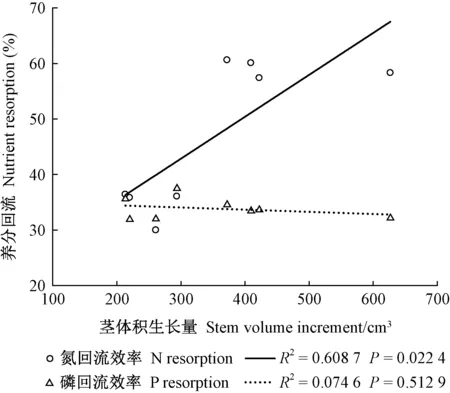
图4 造林后苗木茎体积生长量与养分回流间的线性关系Fig.4 Linear relationship between stem volume increment and nutrient resorption at field

表5 养分回流效率与叶片养分浓度变化间的线性关系Tab.5 Linear relationship between nutrient resorption and leaf nutrient concentration
3 讨论
3.1 秋季施肥对苗木质量和造林效果的影响
本研究表明,秋季施肥对苗圃阶段苗木生长的影响较小,但可显著提高苗木茎、根中的氮贮存,促进造林后的生长表现,与以往许多研究结果一致(Boivinetal.,2004;Rikalaetal.,2004;Islametal.,2009;Zhuetal.,2013)。然而,Boivin等(2002)对黑云杉(Piceamariana)进行“反指数”秋季施肥发现,苗木生物量积累比未施肥苗木下降13%~27%;Andivia等(2014)发现,秋季施氮或氮、磷、钾肥可显著提高冬青栎的生物量积累(13%~85%)。也有研究指出,苗圃阶段养分贮存的变化并不影响造林表现(Birchleretal.,2001;Andiviaetal.,2011)。树种特性、试验条件、施肥量和施肥方式等可能是造成上述差异的原因。本研究试验条件下,施肥量每株10 g时养分贮存浓度最高,田间表现也最好;施肥量每株20 g时造林后生长受抑,成活率最低。
秋季施肥带来的苗木体内氮积累是以消耗非结构性碳水化合物为代价的(van den Driessche 1985;Margolisetal.,1986)。Margolis等(1986)研究表明,秋季施肥使花旗松(Pseudotsugamenziesii)体内氮浓度增加的同时也消耗了非结构性碳水化合物,细根中糖浓度下降尤为显著。刘勇等(2000)对毛白杨苗木进行秋季施肥处理,苗木茎内的可溶性糖含量比对照提高10.2%~32.7%。本研究中,秋季施肥后根淀粉浓度提高17.9%~34.5%,但差异不显著,且并未因氮贮存的提高而消耗,这可能与落叶树种秋季叶片凋落前较为集中的养分回流有关(Aerts,1996;Vergutzetal.,2012)。秋季施肥后,花旗松消耗根中的非结构性碳水化合物用于养分吸收(Margolisetal.,1986);而落叶树种在秋季养分回流的同时,衰落叶中的非结构性碳水化合物也可能回流至其他组织(Vergutzetal.,2012),用于养分吸收和翌年生长等(Millardetal.,2010;Villar-Salvadoretal.,2015)。目前,关于养分回流过程中非结构性碳水化合物变化的研究较少,今后可结合其变化与根部养分吸收进一步探究苗木养分回流与吸收的动态过程。此外,品种特性、试验条件、取样时间、肥料类型、施肥量和施肥方式等可能是造成本研究结果与刘勇等(2000)结果不同的原因。
3.2 秋季施肥对造林后苗木叶片养分浓度和养分回流的影响
本研究中,未施肥苗木造林后氮回流效率(34.5%)和磷回流效率(34.2%)低于木本植物的均值(48.4% 和53.3%)(Yanetal.,2017),也低于温带落叶植物均值(57.6%和 54.1%)(Vergutzetal.,2012),而施肥苗木氮回流效率(59%)高于平均水平。本研究中氮回流效率显著高于Salehi等(2013)不同杨树品种和无性系的回流效率(12.09%~18.93%),磷回流效率与其相似(32.46%~39.63%),但低于Harvey等(1999)不同杂交杨无性系的回流效率(57% 和64%)。不同的回流效率表明,养分回流存在树种或无性系特异性(species-or clones-specific)。此外,试验条件、树龄、取样时间和测定方法等也是造成差异的可能原因(Brantetal.,2015)。Harvey等(1999)研究发现,低氮浓度下钾不回流。本研究试验条件下,钾未回流,这可能与土壤中钾浓度高有关(表1),也可能与氮浓度低有关。
秋季施肥可显著提高氮回流效率(比对照提高71%),但对磷回流效率无显著影响。由苗圃秋季施肥引起的氮回流效率提高与造林后生长呈显著正相关,表明秋季施肥带来的养分贮存有利于造林后生长,且可促使苗木回流更多养分供后续需求,与以往研究结果一致(Pregitzeretal.,1990;Nambiaretal.,1991)。而Harvey等(1999)认为,生长速率对养分回流的作用较小;Salehi等(2013)指出,生长最优的美洲黑杨(Populusdeltoides)无性系“Harvard”回流效率最低。树种特性、树龄、试验条件、取样时间以及计算方法等可能是造成上述差异的原因。
本研究中,氮回流效率与落叶氮、落叶钾浓度呈负相关,与绿叶磷、落叶磷浓度呈正相关,磷回流效率与绿叶氮浓度呈正相关,这表明回流效率的变化并非受单一元素控制,而与叶片中多种元素的浓度变化有关,与See等(2015)研究结果相似。秋季施肥苗木落叶中的钾淋失相比未施肥苗木降低37.3%,可能与其生长快、需求量大有关。但本研究未考虑造林地土壤肥力对养分回流的影响。Hodge(2004)研究表明,土壤肥力高,植物从土壤中吸收的养分也高,可能会削弱对养分回流的依赖,养分吸收与回流间可能存在平衡机制(trade-off)。此外,关于施肥量对养分回流的影响也存在争议(Maoetal.,2013;Yuanetal.,2015;Kouetal.,2017)。因此,今后可结合造林地土壤肥力,进一步探究不同苗圃施肥水平对造林后养分吸收与回流影响的内在机制。
4 结论
本研究试验条件下,秋季施肥对苗圃阶段苗木生长的影响较小,但可显著促进苗木尤其是根系氮贮存和非结构性碳积累。秋季同时施用氮、磷、钾肥,主要提高氮贮存而不影响磷、钾贮存,表现出氮元素利用偏好性。翌年造林后,苗高、地径和茎体积生长量在施肥量每株10 g时达到最大,苗圃施肥效果得以充分体现,苗圃施肥量每株20 g的苗木造林后生长受抑、成活率最低(63.3%)。苗圃合理秋季施肥能够提高苗木氮贮存和非结构性碳积累,进而提高苗木翌年造林成活率和生长表现。施肥量每株10 g时最有利于苗木养分和非结构性碳水化合物贮存及造林后成活和生长,施肥量每株20 g时则产生负面效应。造林后,苗木生长量提高可提升叶片氮回流效率,降低落叶氮淋失。
