湿地松人工林土壤呼吸及其组分对模拟酸雨的响应*
2021-09-04房焕英肖胜生余小芳欧阳勋志秦晓蕾
房焕英 肖胜生 余小芳 熊 永 欧阳勋志 秦晓蕾
(1.江西农业大学林学院 南昌 330045;2.江西省水土保持科学研究院 南昌 330029;3.江西省土壤侵蚀与防治重点实验室 南昌 330029;4.江西师范大学 南昌 330022;5.江西省水利规划设计研究院 南昌 330029)
自20世纪以来,随着社会的进步和工业的发展,大气酸沉降问题随之出现,并对全球生态系统产生了重大影响(Reayetal.,2008;Enzaietal.,2016),其中,酸沉降激增导致的土壤酸化能强烈改变陆地生态系统碳汇强度及分布格局(Piaoetal.,2009),进而增加碳循环-气候反馈评估的不确定性。作为陆地生态系统碳循环的关键过程之一,土壤呼吸是土壤碳排放的最主要途径(Metcalfeetal.,2011)。森林是陆地生态系统的主体,土壤呼吸速率微小的变化都可能影响到陆地生态系统土壤碳积累(Scharlemannetal.,2014);然而,由于区域气候、土壤营养、植被类型、酸雨沉降量和持续时间等不同,有关酸雨影响森林土壤呼吸的研究结论差异较大,主要表现为可抑制(李一凡等,2019a)、促进(Zhouetal.,2016)或不影响(Chenetal.,2020;Zhengetal.,2019)等。随着研究不断深入,土壤微生物呼吸和植物根系呼吸是土壤呼吸的两大重要组分已成为共识(土壤动物呼吸较弱,一般忽略不计),二者发生机制不同,对酸雨的响应也存在差异(庞蕊等,2017),因此,区分土壤呼吸各组分对理解土壤呼吸机制及其环境变化响应非常关键。
东南部亚热带是我国酸雨最严重的区域(张勇等,2011),了解东南部亚热带森林土壤呼吸对酸雨的响应规律,对认识全球碳平衡和未来气候变化具有重要意义。鉴于此,本研究以我国东南部亚热带地区栽植面积较大的湿地松(Pinuselliottii)林为对象,连续定位观测土壤呼吸速率,分析不同强度酸雨对土壤总呼吸速率(Rs)及其组分(微生物异养呼吸速率Rh和根系自养呼吸速率Ra)的影响规律,以期进一步了解森林土壤呼吸过程,为酸沉降胁迫下的森林管理提供科学依据。
1 研究区概况
2 研究方法
2.1 试验设计

根据研究地点气象站多年资料,4—7月(雨季)降雨约占全年的50%,8—10月(一般为台风雨)降雨约占全年的20%,11月至翌年3月(旱季)降雨约占全年的30%(Liuetal.,2016),酸沉降试验尽量参照该降雨比例,分别于2015年4、8和11月在各样地喷洒50 L不同浓度的酸溶液,喷洒时尽量使酸溶液完全渗入土壤,所有处理均在自然条件下进行,目前降雨视为现实情境背景值,模拟酸雨量为酸雨增量。
2.2 土壤呼吸分离与速率测定
采用壕沟法分离土壤呼吸组分(Jassaletal.,2006)。每块样地随机设置3个1 m × 1 m 样方(去根处理)作为土壤异养呼吸(或称微生物呼吸)速率(Rh)测定点,在样方四周挖掘深100 cm、宽20 cm壕沟,将玻璃纤维薄片放入沟中以阻止根向样方内生长,然后把沟填平。在每个Rh测定点附近设置1个50 cm × 50 cm小样方作为土壤总呼吸速率(Rs)测定点。通过有根和无根样方土壤呼吸速率的差值确定土壤自养呼吸(或称根系呼吸)速率(Ra)。将PVC呼吸圈(高10 cm、内径20.4 cm)沿样地等高线上、中、下位置布设,呼吸圈露出土壤表面2~3 cm。为减小样方内死亡根系分解造成的微生物呼吸速率升高,在土壤呼吸速率正式测定前5个月,即2014年7月进行分离工作(前期在湿地松人工林地监测表明,隔离5个月后,沟内死亡根系大部分已分解)。
采用LI-8100A土壤碳通量自动测量系统(LI-COR Inc.,Lincoln,NE,USA)测定土壤呼吸速率。测定期为2015年1—12月,每月2次,选取晴朗天气08:30—11:30进行。为分析土壤呼吸速率的短期效应,酸处理后1周内每天测定1次(视天气而定)。每个采样点每次有效测定时间为2 min,3次重复。土壤呼吸速率测定前一天,清除呼吸基座内的活体植物,以确保测定的呼吸速率不受植物影响。测定土壤呼吸速率时,采用LI-8100A系统自带的土壤温湿度探头同步测定5 cm深处土壤温度和10 cm深处土壤湿度(通过烘干法校正)。
2.3 土壤样品采集与测定
2015年1、4、6、8、11月,在每块样地内按照“S”形设置5个采样点,去除地表凋落物,用取土钻按0~5和5~10 cm进行分层取样。将样品带回实验室过2 mm钢筛,一部分新鲜土样置于4 ℃冰箱内冷藏保存,另一部分土样进行风干处理。

根深蒂固的应试观念影响了高中生的思想教育,要改变这种现状,要认识到应试体制已不适应时代的发展要求。学校要树立新时代的人才培养观,要建立科学的思想教育管理体系,尽快推进应试教育向素质教育转变。要立足于人的发展,将人的素养提升置于首位,要重视学生潜能的发掘、个性的培养。
2.4 数据分析
采用 Excel 软件进行数据基础分析,SPSS软件进行统计分析,Sigmaplot软件进行制图。使用最小显著差异法(LSD)比较不同数据间的差异,Pearson分析方法检测指标间的相关性,多元逐步回归统计分析土壤呼吸的主要影响因子。
采用经验指数模型描述土壤呼吸速率(R,μmol·m-2s-1)随土壤温度(T,℃)的变化(Shietal.,2009):
R=aebT。
(1)
土壤温度敏感系数(Q10)的计算方法(崔海等,2016)为:
Q10=e10b。
(2)
土壤湿度(W,%)和土壤呼吸速率R的关系采用二项式表示(崔海等,2016):
R=aW2+bW+c。
(3)
土壤呼吸速率与土壤湿度、土壤温度的复合关系采用非线性模型拟合(彭信浩等,2018):
R=aebTWc。
(4)
运用赤池信息准则(Akaike information criterion,AIC)衡量统计模型的拟合效果(巫志龙等,2019):
(5)
式中:RSS为残差平方和;n为样本量;K为模型自变量个数。
3 结果与分析
3.1 土壤呼率季节变异
不同酸处理下,Rs、Rh和Ra季节动态类似,均呈夏季高、冬季低的单峰模式(图1)。Rs峰值出现在8月,为5.86 μmol·m-2s-1;最低值出现在1月,为1.45 μmol·m-2s-1。
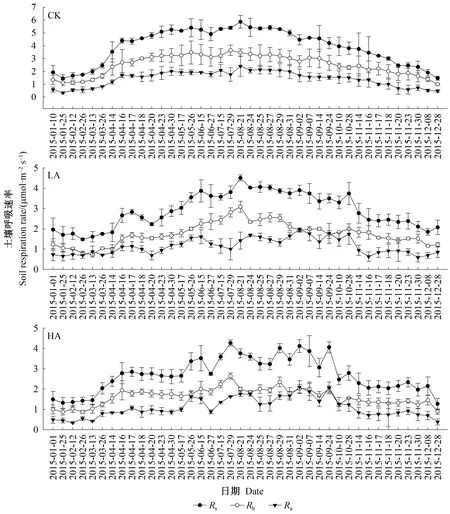
图1 不同酸处理下土壤呼吸速率动态变化Fig.1 Dynamic changes of soil respiration rate under different acid treatments
不同酸处理虽未显著改变Rs、Rh和Ra的季节变化模式,但影响土壤呼吸季节波动,土壤总呼吸速率的变异系数由CK处理的35%降至LA和HA处理的29%和31%。
不同酸处理下,土壤呼吸速率季节变异与土壤温度动态变化基本一致(图2)。各处理的土壤温度差异很小,但土壤湿度存在一定差异。LSD分析表明,与CK相比,LA处理样地的土壤湿度显著偏低(P<0.05),HA与LA处理样地的土壤湿度也有显著差异(P<0.05)。

图2 不同酸处理下土壤温湿度动态变化Fig.2 Dynamic changes of soil temperature and moisture under different acid treatments
3.2 土壤呼吸对模拟酸雨的响应
从年均值来看,不同酸处理对Rs、Rh和Ra均存在抑制效应(图3)。与CK(3.91 μmol·m-2s-1)相比,LA和HA处理后Rs显著降低25.83%和30.95%(P<0.05),Ra由1.41 μmol·m-2s-1(CK)显著下降19.15%(LA)和23.40%(HA)(P<0.05);Rh由2.50 μmol·m-2s-1(CK)显著下降29.60%(LA)和35.20%(HA)(P<0.05)。

图3 不同酸处理下土壤呼吸速率变化Fig.3 Variation of soil respiration rate under different acid treatments
为进一步探讨土壤呼吸对模拟酸雨的响应,按四季进行对比(图3)。LA和HA处理均显著降低生长季Rs(P<0.05),但2种处理间差异不明显,这种抑制效应在春、夏季显著且大于秋季。非生长季(冬季)Rs小于生长季,酸处理后Rs变化不明显,LA处理后Rs有所增加,HA处理后Rs减小(P<0.05)。酸处理后Rs的季节差异也发生改变,CK处理后仅春、秋之间差异不明显,LA处理后季节差异加剧,四季之间均有显著差异(P<0.05),HA处理后季节差异降低,春、夏季与秋季均无明显差异。
在生长季,LA和HA处理均显著降低Rh(P<0.05),这种抑制效应在春、夏、秋季均有体现。其中,酸处理对Rh的抑制作用在夏季表现最强,且HA抑制程度显著高于LA(P<0.05),但春、秋季2种处理间无明显差异。生长季Ra对酸处理的响应规律与Rh相似,但LA、HA显著抑制Ra仅发生在春、夏季(P<0.05),且2种处理间无明显差异。在非生长季,酸处理后Rh响应不明显,但LA可显著促进Ra(P<0.05)。
3.3 异养呼吸占比对模拟酸雨的响应
在CK、LA和HA处理下,Rh在Rs中的占比均值分别为63.94%(59.04%~77.24%)、60.69%(45.48%~74.29%)和60.00%(49.03%~75%)(图4)。LSD分析表明,酸处理后Rh在Rs中的占比显著降低(P<0.05),但LA和HA间差异不显著(P>0.05),这说明酸处理对Rh的抑制作用大于Ra,即与根系呼吸相比,土壤微生物活动更易受酸处理的抑制。

图4 不同酸处理下土壤总呼吸中异养呼吸的占比变化Fig.4 Changes in the proportion of heterotrophic respiration in the total soil respiration under different acid treatments
3.4 土壤呼吸对模拟酸雨的响应机制
3.4.1 土壤呼吸与土壤水热因子的关系 不同酸处理下土壤呼吸速率与土壤温湿度的拟合模型见表1,AIC越小,说明模型拟合效果越好。由表1可知,不同酸处理下,Rs、Rh、Ra与土壤温度均呈极显著指数关系(P<0.01);土壤温度敏感系数(Q10)在LA和HA处理后降低;HA处理后,土壤湿度与Rs、Rh、Ra均极显著正相关(P<0.01),不同酸处理下,土壤湿度对土壤呼吸速率变异的解释程度小于土壤温度。但双因素模型拟合结果显示,土壤温湿度对土壤呼吸速率变化的共同解释程度与土壤温度差异较小。
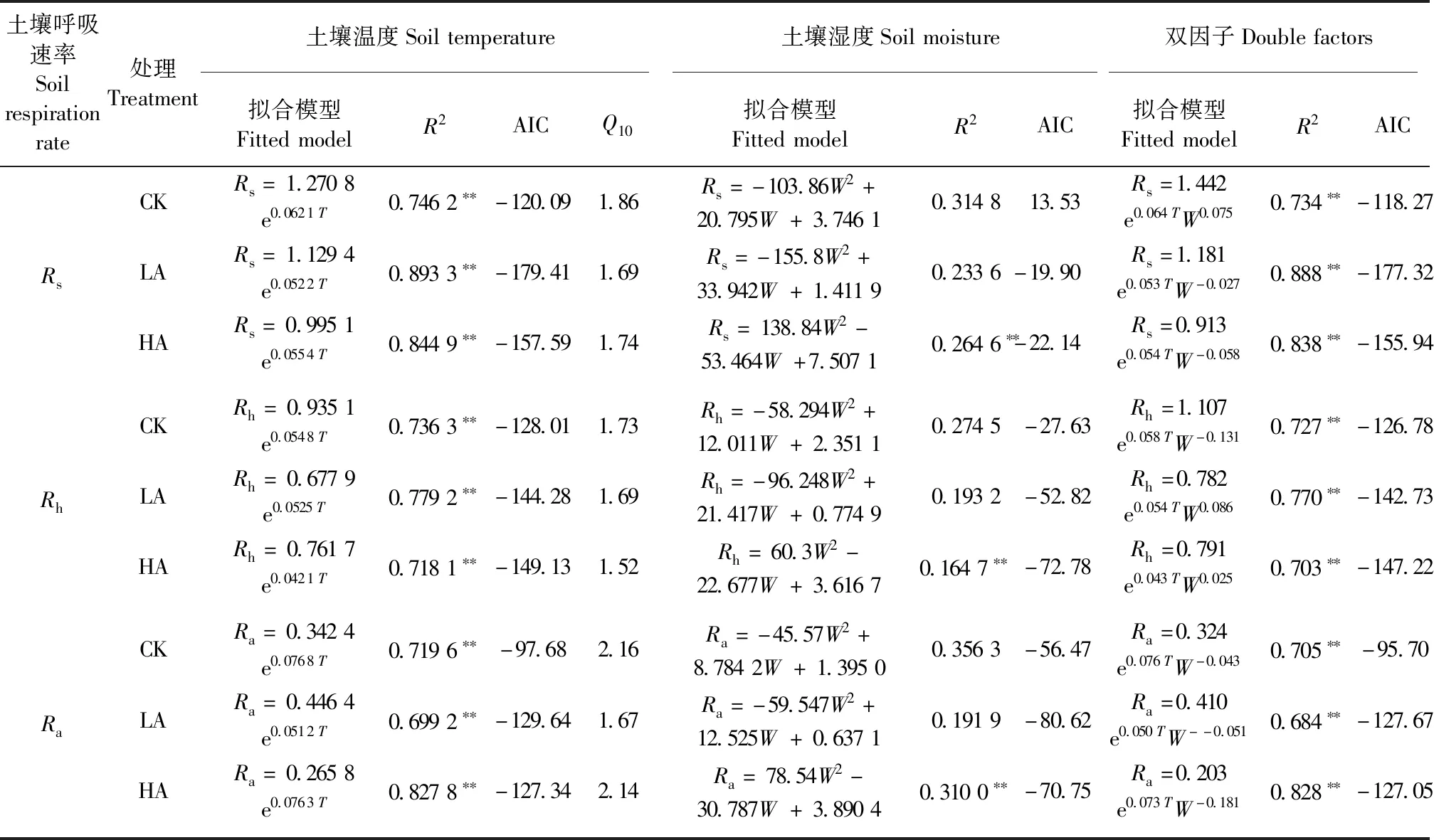
表1 不同酸处理下土壤呼吸速率与温湿度的拟合模型①Tab.1 Fitted model of soil respiration rate and soil temperature,soil humidity under different acid treatments


表2 不同酸处理下土壤呼吸速率与土壤生化特性的皮尔逊相关性分析Tab.2 Pearson correlation analysis of soil respiration rate and soil biochemical characteristics under different acid treatments
3.4.3 土壤呼吸主要影响因素 多元逐步回归分析(表3)进一步表明,脲酶活性、蔗糖酶活性、TOC含量和pH是Rh的主要影响因子(P<0.05),可解释Rh变化的75.8%,脲酶活性、土壤温度和TOC含量是Ra的主要影响因子(P<0.05),可解释Ra变化的89.4%,土壤脲酶活性、蔗糖酶活性、TOC含量和pH是Rs的主要影响因子(P<0.05),可解释Rs变化的83.9%。

表3 土壤呼吸与影响因素的多元逐步回归分析Tab.3 Multiple regression analysis of soil respiration and influence factors
4 讨论
4.1 模拟酸雨对土壤呼吸及其组分的影响
湿地松人工林的土壤呼吸速率存在明显季节变化,对照样地为1.45~5.86 μmol·m-2s-1,高于北亚热带针阔混交林的土壤呼吸速率0.48~5.13 μmol·m-2s-1(陈书涛等,2017),低于西双版纳热带森林的土壤呼吸速率4.18~6.04 μmol·m-2s-1(王亚军等,2016),符合区域尺度上土壤呼吸主要受温度控制的结论(Tangetal.,2006)。酸处理对土壤呼吸季节特征的影响很小,但会显著(P<0.05)抑制土壤呼吸速率及其组分,且这种抑制效应随酸雨强度增加而增强,与大部分研究结果一致(Oulehleetal.,2011;Wuetal.,2016)。但也有研究发现,北亚热带次生林土壤呼吸速率和异养呼吸速率对酸雨响应不明显(陈书涛等,2017);喷施稀释硫酸对温带森林土壤呼吸速率也无显著影响(Zhengetal.,2018)等。这种差异可能与区域气候、土壤背景、植被类型、酸处理强度和持续时间等有关(Zhengetal.,2018;Sitaulaetal.,1995;梁国华等,2016a)。有研究指出,土壤呼吸速率受微生物、基质条件和环境状况等的共同作用,但这些因素同时受到林型影响(Xuetal.,2015;Wangetal.,2018)。Zhang等(2019)研究表明,土壤酸化对森林土壤呼吸速率的作用因森林类型而异,主要取决于土壤和气候条件。不同土壤类型对酸具有不同缓冲能力,这也是酸沉降初期土壤呼吸响应存在差异的主要原因,随着时间推移,酸雨效应出现累积,土壤呼吸会被显著(P<0.05)抑制(李一凡等,2019a);不过该结论需要更广泛、长期的野外试验进行验证。本研究得出,酸雨显著(P<0.05)抑制生长季土壤呼吸速率,非生长季却不明显,说明土壤呼吸速率对酸雨的响应存在季节性差异。张勇等(2001)、梁国华等(2016)也有类似发现,这可能是由于生长季水热充沛、光合作用较强,促使植物根系生长较旺盛,微生物活性较高,对酸雨的抑制响应也更明显。酸雨对异养呼吸速率的抑制现象体现在整个生长季(P<0.05),夏季表现最明显,且HA比LA作用显著;对自养呼吸速率的抑制作用发生在春、夏季,但LA和HA处理之间无明显差异,这可能是由于温度升高促使林木生长对自养呼吸速率产生积极作用,一定程度上可抵消酸雨带来的抑制效果。
4.2 土壤呼吸组分对模拟酸雨的响应机制


土壤水热因子也是影响土壤呼吸的重要非生物因素。本研究表明,土壤温度是土壤呼吸季节变异的主要因素,不同酸处理下土壤温度与土壤呼吸速率及其组分均有极显著的指数关系(P<0.01)。但土壤温度对土壤呼吸速率的影响存在阈值,阈值外二者指数关系不再成立(杨金艳等,2006)。土壤温度对土壤异养呼吸速率变异的解释程度略高于自养呼吸速率,LA处理增加了这种差异性,可能与LA处理下自养呼吸速率的Q10偏小有关,而HA处理提升了土壤温度对自养呼吸速率变异的解释,可能是由于酸处理下异养呼吸速率的Q10有所下降、自养呼吸速率的Q10较大所致。野外自养呼吸包含随植物生长而逐渐增加的呼吸量,可导致其对升温的响应程度更高(Jassaletal.,2006);异养呼吸Q10降低可能与升温促进活性碳库向钝性或缓性转移,引发土壤微生物可利用的活性碳源减少有关(Thornleyetal.,2001)。土壤湿度与土壤呼吸速率及其组分的相关性较弱,土壤温湿度的共同解释程度与土壤温度差异较小证实了这一观点。当土壤湿度变化对植物根系生长和土壤微生物活动影响较小时,不易观测到其对土壤呼吸的影响(余再鹏等,2014),但也可能被其他因子掩盖(吴亚华等,2016)。本研究还发现,LA处理显著降低土壤湿度(P<0.05),HA处理下土壤湿度与土壤呼吸速率及其组分呈显著二次曲线关系(P<0.05),尤其对自养呼吸变异的解释程度较高,这可能与酸雨输入的氮素促进树木蒸腾速率、土壤水分补充不足以抵消生长季的蒸散或非生长季根系自养呼吸增强进而增加水分利用率等有关(闫慧等,2013;王轶浩等,2012;张蕊等,2013)。Deng 等(2009)也指出,土壤水分消耗会导致土壤呼吸与土壤水分的相关性增强。目前,关于酸雨下土壤湿度降低现象的相关研究较少,将在以后研究中进一步佐证。
5 结论
1)模拟酸雨对湿地松人工林土壤呼吸的季节变化影响较小,土壤温度是导致其季节变化的主要驱动因素。
2)酸雨抑制土壤总呼吸速率及其组分,但这种抑制效应仅在生长季显著(P<0.05),在非生长季不明显,且异养呼吸速率对酸雨的抑制响应比自养呼吸速率强。
3)异养呼吸速率在土壤总呼吸速率中占比大于自养呼吸速率,但随着酸雨强度增加而减小,短期内异养呼吸速率仍是土壤总呼吸速率的主导成分。

由于土壤呼吸的影响因素多样,无机环境、生物过程、人为因素均可能成为主要影响因子,今后有待进一步延长模拟酸雨下土壤呼吸的监测时间,并加强各因素对土壤呼吸的交互作用等研究,以期深入了解酸雨加剧情况下森林土壤呼吸响应机制。
