黄瓜黄叶基因YL的精细定位及候选基因预测
2021-08-28熊良荣张克岩何欢乐潘俊松
潘 明,熊良荣,张克岩,潘 健,何欢乐,潘俊松,蔡 润,王 刚
(上海交通大学农业与生物学院,上海200240)
叶是植物进行光合作用、呼吸作用和营养转化的重要器官,叶色直接影响植物光合作用的效率。叶色的变化是由基因型和生长环境决定的[1]。叶色是植物广泛存在的突变性状之一,具有易于鉴定且突变频率高的特点。植物叶色是叶绿体色素共同作用的结果,通常以绿色为常见表型。叶色变化是植物衰老的象征,严重影响植物的光合作用,会造成作物产量降低[2]。叶色突变体可成为研究叶绿素合成与代谢等生理过程的优良材料[3]。在研究较多的模式植物中,如水稻[4-5]、小麦[6]、拟南芥[7-8]、番茄[9-10]、油菜[11]、大白菜[12]等,已鉴定出了大量表型各异的叶色突变体。这些叶色突变体有助于研究光合作用相关基因的功能[3]。例如,在水稻中,已经发现200多个叶色突变体,154个相关基因被分别定位到12条染色体上,其中53个已经被克隆[13]。在大豆中,25个影响叶绿素含量的核基因已经被定位和鉴定[14]。玉米的叶色突变体研究也较多,已报道的玉米叶色突变基因有166个,其中,斑叶突变基因26个,黄绿叶突变基因64个,条纹叶突变基因58个,白叶突变基因18个。在黄绿叶突变体中,发现8个突变(nec1、nec2、nec3、nec4、nec5、nec6、nec∗-1487、nec∗-N1119B)为致死型突变[15]。
与以上植物相比,黄瓜(Cucumis sativusL.)中有关叶色突变体的研究还相对滞后,目前,只报道了10余个叶色突变体,如黄叶转绿(v)、黄叶转绿-1(v-1)、黄色子叶-1(yc-1)、黄色子叶-2(yc-2)、白化致死(pl)、白化转绿(albin)、光敏白化致死突变(ls)、叶绿素缺陷突变(cd)、黄转绿白斑叶(vvi)、金黄叶(yp)、金黄色子叶(gc)和黄绿叶(gyl)[16-20]等,其中有些基因已经被克隆。如Miao等[19]利用一个淡黄色子叶和第一个真叶的绿色叶片突变体9110GT检测到一个单隐性基因(v-1)控制了绿色叶片性状,并通过高分辨率遗传图谱确定了一个v-1候选基因。Gao等[20]发现了一个黄瓜叶绿素缺乏的突变株c528,在整个发育过程中具有金黄叶色,通过多个群体的分离分析表明,该突变是隐性遗传,绿色对金黄色具有完全显性。对更多叶色黄化突变体进行研究,可为深入了解黄瓜叶色突变形成的生理及分子机理奠定基础。
本研究以稳定遗传的黄叶突变体yl为父本、黄瓜自交系S52为母本,构建F2分离群体,通过F2群体不同叶色分离比,确定该突变性状的遗传类型,并利用BSA法结合F2群体分析确定叶色基因染色体位置,开发新的遗传标记确定叶色基因精确物理区间,通过重测序数据的比对分析,推测候选基因。
1 材料与方法
1.1 试材及取样
以华北型黄瓜自交系WD1为材料,采用EMS诱变方式获得黄叶突变体yl,经上海不同季节和地点种植后鉴定为一个稳定遗传的突变体。2018年春以黄叶突变体yl为父本、S52为母本,杂交获得F1。2018年秋对F1进行自交获得F2群体。2019年春在上海交通大学农业与生物学院育苗间分批次种植277个F2单株,10株S52、10株黄叶突变体yl和10株F1,观察统计叶色性状用于遗传分析。所有材料均单株采样,用于混合分组分析法(BSA)分析和基因定位。
1.2 黄叶突变体的性状调查
将黄叶突变体yl、S52、F1种子各10粒,以及277粒F2种子分两批播种于32孔穴盘中,培养在育苗间,温度(昼∕夜)25∕18℃,光强12 000 lx,昼长16 h,定期浇水。对该群体的初生子叶、完全伸展的子叶,以及新生的真叶进行观察记录。
1.3 利用BSA法进行DNA池间多态性标记的筛选及基因连锁关系分析
从F2群体中随机挑选5个黄叶植株等量混合构建黄叶突变体DNA池,等量混合5个绿叶植株的DNA构建绿叶野生型DNA池。以2个DNA池为模板,用平均分布在黄瓜7条染色体上的235对SSR引物进行池间多态性引物筛选。以F2群体植株DNA为模板,用筛选得到的多态性标记进行扩增分析,标记与基因的连锁关系。
1.4 黄叶突变体DNA混池基因组重测序和候选基因预测
在F2中选取20株黄叶表型植株叶片抽提DNA,等量混合后组成DNA混池,委托北京百迈客生物科技有限公司进行基因组重测序,样品的测序深度为20X。参考基因组为华北型黄瓜9930(http:∕∕cucurbitgenomics.org∕organism∕2)。采用GENEIOUS软件,将定位区间内所有基因的基因组序列在混池基因组序列、黄瓜9930参考基因组序列、突变体来源亲本WD1和群体亲本S52重测序基因组序列中进行比对,统计亲本和黄叶植株混池材料中的碱基突变类型,预测候选基因。
2 结果与分析
2.1 黄瓜黄叶突变体yl的表型分析
为鉴定黄叶突变体yl的表型,将该突变体分别于不同季节、不同地点进行种植,经多次调查、统计发现,黄叶突变体yl为表型稳定遗传的突变材料。黄叶突变体yl在子叶初长和子叶完全展开的时候表现为黄叶,随着真叶破心以及子叶的生长,每片新长出的真叶叶色均为黄色。待新的真叶长出后,低节位的真叶开始转绿,直至完全转变为绿色。

图1 黄瓜黄叶突变体yl的苗期表型Fig.1 Seedling phenotype of cucumber yellow leaf mutant yl
2.2 黄瓜黄叶突变体yl的遗传分析
杂交F1代(黄叶突变体yl×S52)的黄瓜植株均表现为野生型的绿色叶,未出现突变的黄色叶。将F1自交得到的F2群体单株分批次(第一批77株,第二批200株)种植后,观察亲本、F1及两批F2不同单株的叶色表型。如表1所示,F2群体中出现了正常的绿色叶和突变的黄色叶的表型分离,卡方检测结果显示正常绿叶色与黄色叶的分离比符合3∶1,说明该黄叶突变性状受一对隐性核基因控制,为质量性状。

表1 黄瓜黄叶突变体yl F2群体叶色性状分离比Table 1 Segregation ratio of leaf color traits of yellow leaf mutant yl in F2 population(S52×yl)
2.3 黄瓜叶色基因YL的染色体定位
为了初步筛选与YL基因相连锁的分子标记,在F2分离群体中,分别取5株黄叶突变植株和5株绿叶野生型植株的叶片提取DNA,等量混合后分别构建黄叶突变体DNA池和绿叶野生型DNA池。基于BSA法,在2个DNA池间对平均分布在黄瓜7条染体上的235对SSR引物进行多态性筛选,发现标记4号染色体的SSR26406、SSR23603、SSR15682、SSR15420、SSR05416这5对引物在2个DNA池间有多态(表2,图2)。利用以上5对引物对F2群体中的20株隐性单株进行分析,发现这5对引物标记均与YL基因连锁。同时也发现,YL基因被定位在黄瓜4号染色体分子标记SSR15682和SSR05415之间(图3,图4)。

图2 DNA混池间多态性SSR标记分析Fig.2 Analysis of polymorphisms between DNA pools by SSR markers

图3 SSR15420在亲本S52、WD1和F2群体中20个隐性单株的PCR分析Fig.3 PCR analysis of SSR15420 in parents S52,WD1 and 20 recessive individuals of F2 population

图4 黄瓜黄叶基因YL在黄瓜四号染色体上的遗传定位Fig.4 Genetic mapping of yellow leaf gene YL on cucumber chromosome 4

表2 基因池间多态性SSR标记Table 2 Polymorphic SSR markers between two DNA pools
2.4 黄瓜叶色基因YL的精细定位
为了进一步缩小定位区间,在初步定位的区间内新开发并筛选了具有亲本多态性的插入∕缺失(InDel)标记InDel4-1、InDel4-2、InDel4-3、InDel4-4、InDel4-5、InDel4-6、InDel4-7、InDel4-8、InDel4-9和InDel4-10。用这些多态性的InDel标记对277个F2单株进行分析,结果显示YL基因被定位在黄瓜4号染色体上的分子标记InDel4-9(图5)和InDel4-10(图6)之间,物理距离为936.39 kb(图7)。
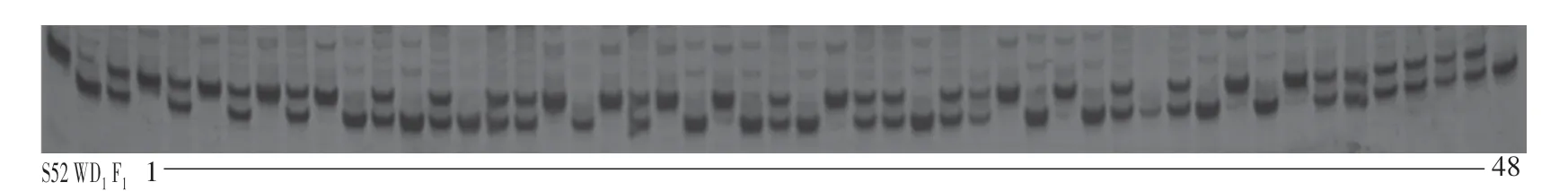
图5 InDel4-9在亲本S52、WD1和48株F2群体中的扩增结果Fig.5 Amplification of InDel4-9 in S52,WD1 and 48 individuals of F2 population

图6 InDel4-10在亲本S52、WD1和部分48 F2群体单株中的扩增结果Fig.6 Amplification of InDel4-10 in S52,WD1 and 48 individuals of F2 population

图7 黄瓜黄叶基因YL在黄瓜4号染色体上的遗传定位Fig.7 Genetic mapping of yellow leaf YL gene on chromosome 4 of cucumber
2.5 黄叶突变体DNA混池基因组重测序和候选基因预测
将黄叶突变体DNA混池进行基因组重测序,将定位区间内所有基因的基因组序列在混池基因组序列、黄瓜9930参考基因组序列、突变体来源亲本WD1和群体亲本S52重测序基因组序列中进行比对,鉴定出定位区间内黄叶植株混池与黄瓜9930、亲本WD1和S52材料的非同义突变。结果表明:在黄瓜9930背景下,黄叶突变体yl存在突变位点25个,WD1存在突变位点23个,S52存在突变位点59个(图8)。通过比对混池DNA和WD1基因组重测序数据,发现4个黄叶突变体yl特有的错义突变位点,这4个特有的错义突变分布在Csa4G291360和Csa4G297530这2个基因中。再通过比对混池材料和S52亲本(表3),发现2个黄叶突变体yl特有的错义突变位点,这2个突变位点仍位于Csa4G291360和Csa4G297530这2个基因,将上述4种突变形式与本实验室保存的所有重测序亲本数据进行比较,发现Csa4G291360基因的这3个突变位点均存在于其他非黄化黄瓜材料中,而Csa4G297530基因的突变形式还未在现有的重测序亲本中有发现,即排除Csa4G291360基因,且Csa4G297530符合EMS诱变的颠换特征。由此可以推测,Csa4G297530为候选基因。

图8 黄瓜黄叶突变体yl与WD1和S52之间的突变位点分析Fig.8 Analysis of mutation sites between yl mutant,WD1 and S52 in cucumber

表3 黄瓜黄叶突变体yl在定位区间内特异突变类型Table 3 Specific mutation types of yl mutants in the location region
3 讨论
目前研究的黄瓜叶色突变体相对较少,李万青等[21]报道了一个非致死性黄化突变体C528,其表型为黄色叶片,在全生育期表达,不随着发育而转变,整个植株表现为全株黄色;胡亮亮等[22]报道了一个黄化转绿突变体C777,表现为子叶抽出即为黄色,随着第一片真叶的长出,子叶慢慢转绿,随着每长出一片新真叶,低节位的真叶则转绿,突变体整个生育期都表现出此类转变。本研究的黄瓜黄叶突变体yl由EMS化学诱变产生,是一个稳定遗传的突变材料,利用图位克隆方法,将黄叶基因定位于黄瓜4号染色体分子标记InDel4-9和InDel4-10之间936.39 kb的物理区间内。通过重测序数据的比对,排除存在相同突变形式的基因,推测Csa4G297530为候选基因。根据前人对黄叶的研究,未曾报道过该基因与控制黄叶性状相关,因此本研究发现了一个新的控制黄叶的突变基因。
植物线粒体可以为叶绿体的光合作用提供CO2和H2O,还可以为叶绿体除光合作用以外的其他生命活动提供能量,如为叶绿体中DNA的转录和翻译等生命活动提供ATP。线粒体与叶绿体在功能上都可以进行能量转换,线粒体产生的ATP用于细胞的代谢,叶绿体产生的ATP只用于光合作用暗反应阶段。Csa4G297530基因是线粒体底物载体蛋白,包含IPR002067(线粒体载体蛋白)和IPR023395(线粒体载体域)。Dinakar等[23]报道了疑似通过刺激细胞活性氧的水平,线粒体呼吸的抑制能够降低叶片对CO2的固定能力。贾凌云等[24]研究发现,线粒体呼吸作用的抑制并不会改变光反应阶段的化学能转化,但降低了叶片的光合速率。Chen等[25]研究显示,线粒体解偶联蛋白对维持有效的光合作用是必要的。因此,Csa4G297530基因的突变可能会影响黄瓜的光合作用,造成黄叶表型的形成。
