基于广义可加模型分析温湿度因子对斑翅果蝇田间种群动态的影响
2021-08-13赵远鹏李正跃张立敏
陈 晓,赵远鹏,李 欣,肖 春,李正跃,张立敏**
(1.云南农业大学植物保护学院,昆明 650201;2. 省部共建云南生物资源保护与利用国家重点实验室,昆明 650201;3. 昆明英茂花卉产业有限公司,昆明 650000;4. 云南农业大学大数据(信息工程)学院,昆明 650201)
斑翅果蝇DrosophilasuzukiiMatsumura,又称樱桃果蝇,隶属双翅目Diptera环裂亚目Cylorrhapha果蝇科Drsophilidae果蝇属Drosophila水果果蝇亚属Sophophora(孙鹏等,2011)。斑翅果蝇寄主范围非常广泛,已知寄主涉及100多种,主要包括蓝莓、樱桃、草莓、葡萄等(Kenisetal.,2016),且该果蝇的主要寄主植物为高密度单一种植模式,并且该果蝇能以其它野生寄主和观赏植物作为庇护场所和再侵染源(Klicketal.,2012),所以容易导致该果蝇种群数量在种植区域内快速增长甚至爆发。近年来,随着各国之间的贸易往来加深,在世界范围内,斑翅果蝇除了在适宜的自然或管理环境中扩散,也随着人为的商品运输及交通工具的携带进行传播,还有以具潜在风险的有害生物为载体进行协同扩散,利用广泛分布于边境地带的贸易商品进行扩散,更有甚者附带于农副产品和废弃物中进行快速地分布扩散(Hauseretal.,2009)。现已有多个国家相关检疫部门高度重视斑翅果蝇日渐增长的危害,发布了相关预警或将其列入检疫性有害生物名单(孙鹏等,2011)。
光照、温度、湿度、降雨等气象因子可综合作用于昆虫的生长发育、生殖繁衍、行为活动、生存分布等,其中温度是影响上述生物学指标的关键气象因子,自然环境中温度的变化直接影响昆虫新陈代谢、生长以及繁殖等生命活动的速率,从而对昆虫的发生数量、体形大小、行为变异以及活动期长短产生十分重要的直接作用(石保坤,2012);环境湿度(包括空气相对湿度和土壤含水量)的变化可直接导致昆虫体内含水量的变化,影响虫体的水分平衡、生长发育及繁殖速率、抗逆性、行为诱导信号等,进而对其个体发育及群体发生产生影响(常晓娜等,2008)。因此,外界环境温度、湿度的变化是昆虫种群随着季节消长变化的主要因素,且温度与湿度因子联合作用下可增强上述现象的表达,在高温和低相对湿度的双重影响下,不育或可育斑翅果蝇的生存和繁殖均受到负面影响(Krügeretal.,2021)。
在解释生物学原理和自然种群规律方面,非线性模型均优于线性模型,非线性模型已经广泛应用于昆虫生长发育和种群动态研究。早期研究人员对昆虫种群动态研究以描述性为主,并进行种群聚集与扩散迁移的研究,现在则应用模型模拟来研究昆虫与环境之间的相互作用关系,目的是为了加强理解昆虫种群的生命系统,改进对昆虫种群的监测调查和防治的具体措施。而关于斑翅果蝇种群动态模型研究的文章较少,例如,整合与温度相关的生命表数据并通过矩阵投影模型来估算斑翅果蝇的种群动态(Wimanetal.,2014);或应对入侵北美地区的斑翅果蝇开发了一个连续的时间阶段种群结构模型(Langilleetal.,2016)。Hastie和Tibshirani(1986)提出并发展了广义可加模型(generalized additive models, GAM),该模型可拟合响应变量与解释变量之间的非线性关系,但模型需要考虑曲线的拟合优度、光滑度,需要克服解释变量之间共曲线性关系导致的系数标准误偏差,并利用两个解释变量之间的Pearson相关系数R来判别两者之间的相关性程度。若要求得到的拟合曲线平稳变化,不出现非剧烈快速波动的情况,则需要通过响应变量与解释变量之间的关系曲线粗糙度来度量拟合光滑度(欧阳芳和戈峰,2013)。
本研究在昆明市云南农业大学后山设置诱捕点,调查统计2016年至2017年连续两年斑翅果蝇成虫的种群动态变化,明确调查期内斑翅果蝇成虫种群数量动态,分析斑翅果蝇成虫种群数量分布,通过广义可加模型(GAM)分析斑翅果蝇成虫种群数量与温湿度因子间的关系,并判别是何种关键因子显著影响斑翅果蝇成虫种群数量动态变化。不仅从生态学角度探寻了温湿度因子对斑翅果蝇成虫种群动态消长规律起到重要作用,也为后续研究斑翅果蝇产生何种适应性应对环境因子变化提供了重要的依据。
1 材料与方法
1.1 试验地点
2016至2017年,在云南农业大学校园后山自然果林(N25°8′16″,E102°45′15″,海拔1 970 m)进行连续两年的斑翅果蝇成虫种群动态调查。所选果林树势良好,主要以梨树为主,夹杂少量桃树,果林无化学药剂喷施,地面生长有龙葵、蛇莓等野生浆果。
1.2 田间诱捕及温湿度记录
诱剂配制比例参照优化配比,即糖(g)∶酒(mL)∶醋(mL)∶水(mL)=1∶3∶1∶6(伍苏然等,2007)。采用五点取样法,将自制诱捕器放置在离地面1.2~1.5 m处,不同取样点之间距离至少间隔10 m。温湿度记录仪(GGL-20)由深圳宇问加壹传感系统有限公司生产提供,悬挂位于果林较为隐蔽处。
春、冬两季因自然环境气温较低,诱剂不易挥发,故收集频率为1周一次,夏、秋两季因自然环境气温较高,诱剂易挥发,故收集频率为 2或 3 d一次。将诱捕到的所有昆虫样本带回实验室,在解剖镜下进行分类鉴定,并统计斑翅果蝇雌雄成虫数量。
1.3 广义可加模型
广义可加模型分析可分为以下3个步骤,依次为:变量预分析、模型构建、模型预测。广义可加模型是根据具体情况将广义线性模型和可加模型结合起来,其中,广义线性模型是改变响应变量条件期望的函数形式,将它记为g(muY),其中muY=E(Y/X1,…,Xn),可表示为:g(muY)=α0+α1X1…+αnXn+ε,还可用非参数的形式来描述响应变量条件期望与解释变量的对应关系,用f(x)表示,可表示为:E(Y/X)=f(x),将其推广到多个解释变量时,则记为可加模型E(Y/X1,…,Xn)=f(X1)…+f(Xn)。将两种方式结合起来,则广义可加模型可表达为:g(muY)=α0+f1(X1)…+fn(Xn)+ε。其中,在g(muY)项中,muY是Y的期望值,g(…)是连接函数,α0是截矩,在fn(Xn)项中,fn(…)是解释变量Xn的单变量函数,ε为随机变量。模型评价指标选用广义交叉验证值(Generalized Cross-Validation, GCV),GCV值最小的模型最优(欧阳芳和戈峰,2013)。
1.4 数据分析
使用Excel对数据进行整理和预处理,利用R(version 4.0.5)对斑翅果蝇种群数量数据进行对数预处理,mgcv数据包拟合广义可加模型并进行分析。
2 结果与分析
2.1 成虫田间种群发生动态
2016年至2017年,两年中全年均有斑翅果蝇发生,斑翅果蝇成虫种群数量均从6月开始明显增加至10月种群数量回落,高峰期分别出现在7月、8月、9月,在11月至次年5月斑翅果蝇成虫种群数量都维持在较低水平,尤其在2-5月种群数量呈全年最低水平。雌、雄成虫的种群动态变化也呈现先升高后降低的趋势;其中,雄成虫种群动态变化与总种群动态变化趋势一致。2016年种群变化趋势呈单峰型,而2017年呈双峰型;其中,雌成虫种群变化趋势在两年中均呈单峰型,最高峰分别出现在2016年7月、2017年8月。2017年斑翅果蝇成虫数量,尤其是高峰期数量相较于 2016年明显降低。2016年、2017年最高峰期斑翅果蝇成虫总量分别为587头、236头,雄成虫数量分别为356头、130头;雌成虫数量分别为312头、106头(图1)。
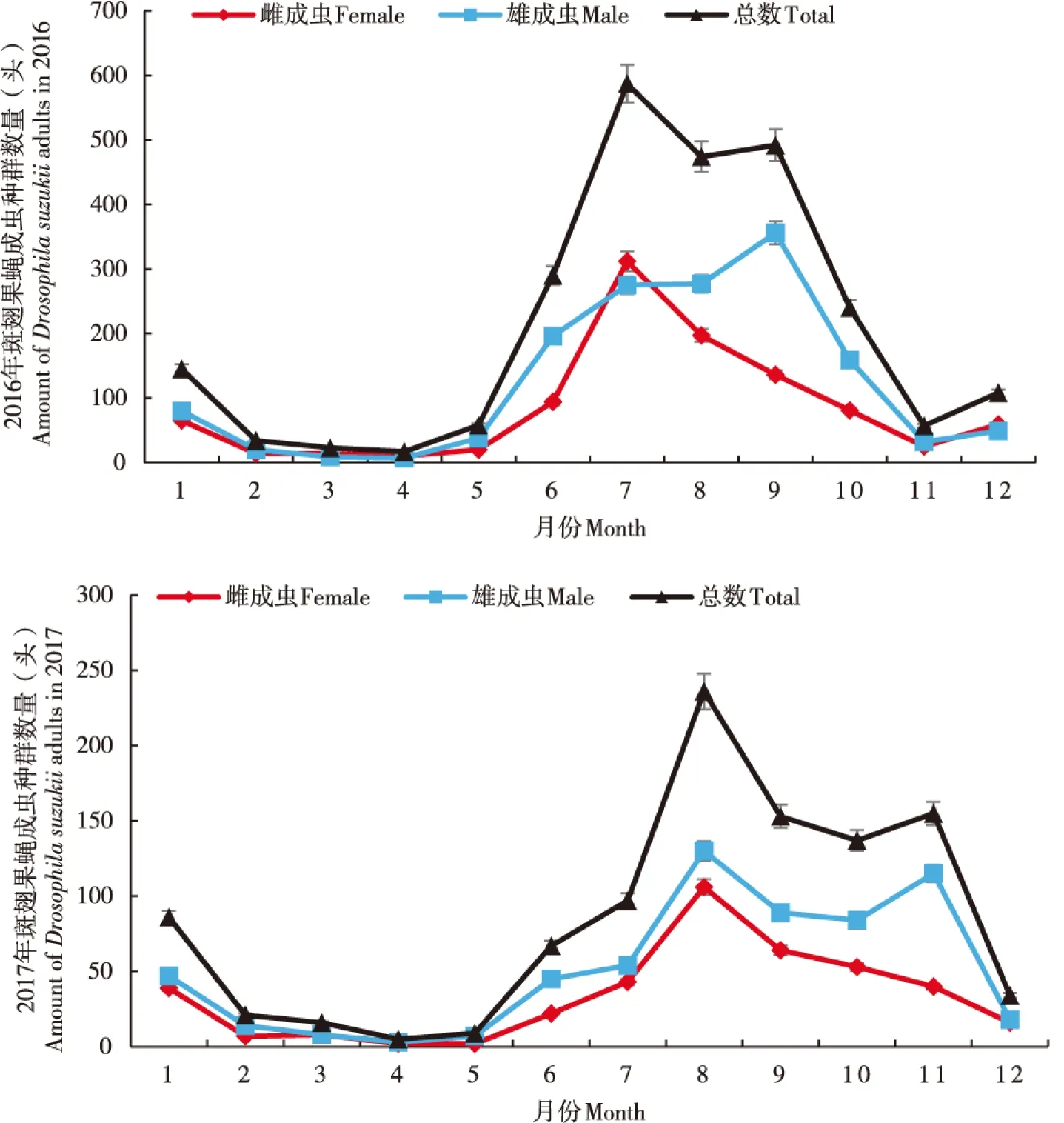
图1 2016-2017年斑翅果蝇成虫种群动态Fig.1 Population dynamics of Drosophila suzukii adults from 2016 to 2017
2.2 广义可加模型分析温湿度因子与斑翅果蝇成虫种群数量之间的关系
2.2.1变量预分析
对2016-2017年斑翅果蝇雌成虫、雄成虫和总种群数量取对数并绘制QQ图和频次分布图(图2、图3、图4),并初步判定种群数量(响应变量)的分布特征为正态分布类型,Identity link可作为连接函数,进而将种群数量作为相应变量Y,温度(Temperature)、湿度(Humidity)因子作为解释变量构建模型。
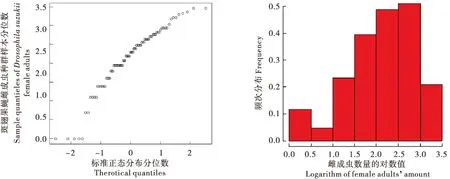
图2 斑翅果蝇雌成虫种群数量正态QQ图和频次分布图Fig.2 Normal QQ plot and frequency distribution diagram of Drosophila suzukii female adults
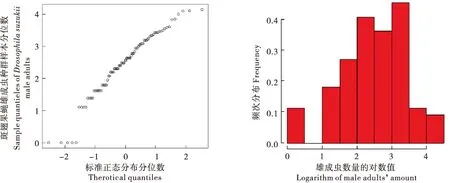
图3 斑翅果蝇雄成虫种群数量正态QQ图和频次分布图Fig.3 Normal QQ plot and frequency distribution diagram of Drosophila suzukii male adults
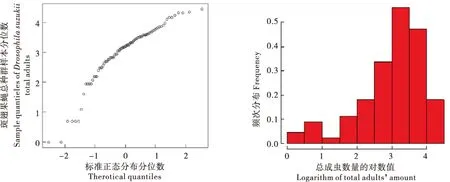
图4 斑翅果蝇总种群数量正态QQ图和频次分布图Fig.4 Normal QQ plot and frequency distribution diagram of Drosophila suzukii total adults
2.2.2模型构建
分析调查周期内温度与湿度因子间Pearson相关系数,得知两个解释变量间Pearson相关系数小于0.5,故本研究将斑翅果蝇雌成虫、雄成虫、总种群数量作为响应变量,温度(Temperature)、湿度(Humidity)作为解释变量构建广义可加模型。对于多变量数据集,模型构建时可构造一个或多个可行模型,以不同解释变量的组合构成多个分析模型。因此,初步构建响应变量与解释变量间的分析模型是包括多个可能模型的模型组,再通过模型评估筛选最优拟合结果。
g(Female)=αF+f1F(Temperature)+f2F(Humidity)+εF
g(Male)=αM+f1M(Temperature)+f2M(Humidity)+εM
g(Total)=αT+f1T(Temperature)+f2T(Humidity)+εT
其中,g(Female)、g(Male)、g(Total)分别是指斑翅果蝇雌成虫、雄成虫以及总种群动态的连接函数;α是函数截矩;在fn(Xn)项中,fn(…)是解释变量Xn的单变量函数;ε是随机变量。3个下角标F、M、T分别是指雌成虫、雄成虫以及总种群。
模型参数拟合结果表明,温湿度因子显著影响了2016-2017年斑翅果蝇田间成虫种群动态(P<0.05),雌成虫、雄成虫、总种群数量的GAM模型参数α分别为2.21346、2.55606和3.08316;雌成虫、雄成虫和总种群数量与温湿度因子关联的GCV值分别为0.57299、0.74501和0.64611,其中雌成虫的GCV值最小,因此说明斑翅果蝇雌成虫种群数量与温湿度因子进行拟合的模型最优(表1)。
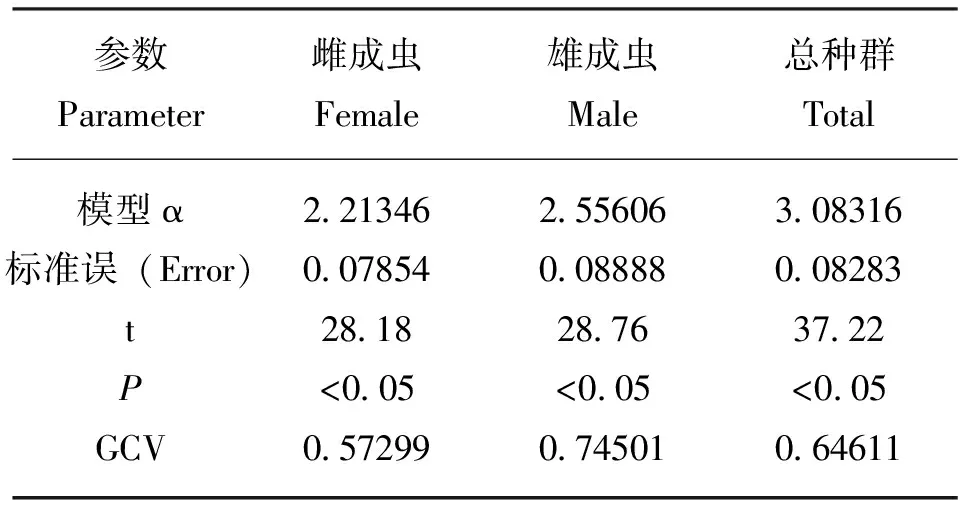
表1 基于温湿度因子的斑翅果蝇种群动态广义可加模型参数拟合及检验
2.2.3模型预测
斑翅果蝇雌成虫种群动态与温湿度因子之间呈非线性相关,其中雌成虫种群发生在温度23℃以下呈局部负相关,在23℃以上呈局部正相关且上涨趋势较为平缓;雌成虫种群发生与湿度呈正相关,但在湿度变幅大的情况下种群发生增量小(图5)。斑翅果蝇雄成虫的种群动态与温湿度因子之间呈非线性相关,雄成虫种群发生在温度23℃以下呈局部负相关,在23℃以上呈局部正相关,且上涨趋势较为明显;雄成虫种群发生与湿度呈正相关,但在湿度变幅大的情况下种群发生增量小,斑翅果蝇总种群发生与雄成虫种群发生表现基本一致(图6,图7)。
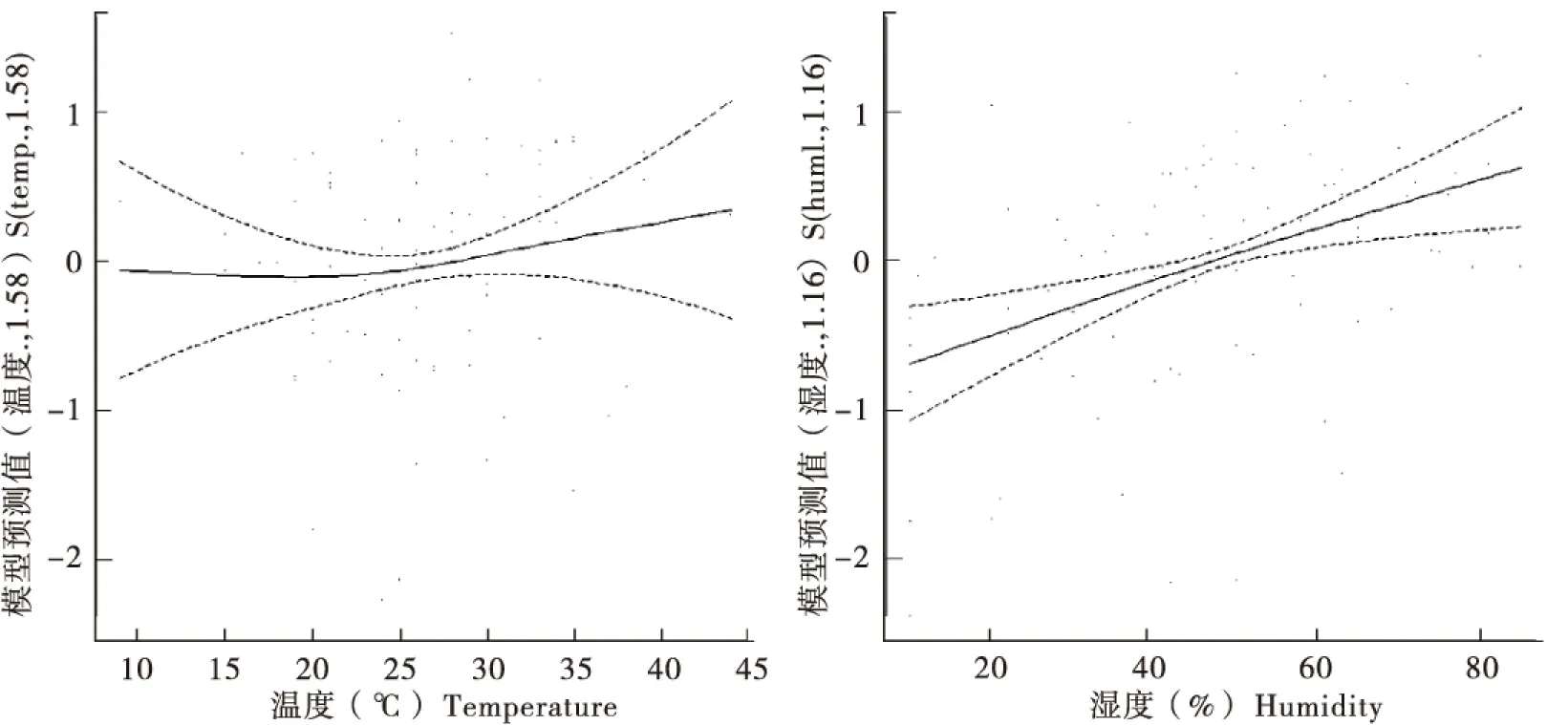
图5 斑翅果蝇雌成虫种群数量与温湿度因子之间的相关性Fig.5 Population sizes of Drosophila suzukii female adults correlated with temperature-humidity factor注:Y轴标题括号中的数值为模型拟合的光滑度参数,该数值接近1,预测变量与目标变量间的相关关系越接近水平直线,反之曲线呈现不同相关关系。图6-图7同。Notes : The value in the brackets of Y-axis heading is the smoothness parameters of the prediction model. If the smoothness parameter is close to 1, the relationship between the predictor variables and dependent variables would be closer to a horizontal straight line, otherwise, the relationship would be different correlations. Same to Fig.6-Fig.7.
图6 斑翅果蝇雄成虫种群数量与温湿度因子之间的相关性Fig.6 Population sizes of Drosophila suzukii male adults correlated with temperature-humidity factor
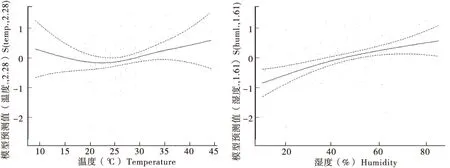
图7 斑翅果蝇总种群数量与温湿度因子之间的相关性Fig.7 Population sizes of Drosophila suzukii total adults correlated with temperature-humidity factor
3 结论与讨论
3.1 斑翅果蝇成虫全年种群数量动态规律及分析
种群动态调查是昆虫种群生态学研究的核心问题,尤其是为研究昆虫田间种群的适应性在环境因子变化下发生何种改变提供了基础和前提。国外已有对斑翅果蝇的种群动态变化及寄主的调查研究,例如,在比利时对斑翅果蝇种群发生进行调查,结果表明2014年的早春(非常温和的冬季之后),已经发现了斑翅果蝇的存在(Belienetal.,2014);研究发现从1月开始,当休眠状态下的斑翅果蝇雌成虫有效积温达到50~800℃·d时,则可在季节早期进行繁殖,而且可能会受到早期寄主植物可用性的限制(Wiman etal.,2016)。而斑翅果蝇在国内的研究主要集中在果园内进行种群动态的监测调查以及为害特性观察的研究,例如,调查研究发现陕西灞桥樱桃果园里斑翅果蝇为优势种(郭丽娜等,2014);陕西杨凌樱桃园中斑翅果蝇种群数量在10月中旬达到高峰期(常慧群等,2017);在斑翅果蝇发生的高峰期,云南石屏果园中的杨梅果实被害率可达100%(赵超等,2017)。在本研究中,2016-2017年调查发现斑翅果蝇成虫的种群数量通常在野生果林的果实进入成熟期后大幅增长,高峰期集中在 7月至9月,与不同引诱剂对贵州麻江田间蓝莓果蝇诱捕结果基本一致(徐芳玲等,2012),即8月蓝莓果实进入盛果期,成熟的蓝莓对果蝇的诱集数量呈现高峰值;在2月至5月,斑翅果蝇成虫种群数量动态均维持在较低水平,与陕西杨凌樱桃园中斑翅果蝇发生特点基本一致(杨静,2017);斑翅果蝇雄成虫以及总种群数量动态变化趋势基本同步,其中总种群发生动态与王华弟等研究浙江地区杨梅果蝇种群发生动态时监测得出的结果基本一致(王华弟等,2017)。
3.2 温湿度因子与斑翅果蝇成虫种群动态之间的相关性分析
温湿度变化对昆虫的生长发育速率、生殖能力及存活率等种群数量构成的关键因子产生重大影响,从而导致昆虫的种群数量发生变化(陈瑜和马春森,2010)。此前有研究报道,结合2016-2018年气温和降水量资料,分析大豆蚜虫Aphisglycines田间动态与气温和降水量的总体关系,结果得出气温以20℃为节点起伏,田间温度越高大豆蚜虫发生量越大,而降水量对大豆蚜虫田间种群动态影响不明显(邵珊珊等,2019)。滇中菜区小菜蛾Plutellaxylostella成虫种群消长动态与温度显著相关r=0.442,与降雨量成正相关r=0.283,但相关程度弱(赵雪晴等,2016)。假眼小绿叶蝉Empoascavitis的成虫虫口数量受田间气候条件影响较大,其中温度影响最大(李慧玲和林乃铨,2012)。本研究中以温度23℃为节点,斑翅果蝇成虫种群发生数量与23℃以下呈局部负相关,与23℃以上呈局部正相关,与湿度呈正相关,但在湿度变幅大的情况下种群发生增量小,得出湿度对斑翅果蝇的种群影响较温度的影响低,温度是影响斑翅果蝇成虫种群数量动态的关键生态因子,这一结果与上述温湿度因子对其他昆虫种群发生的影响研究结果趋向一致。
本研究中斑翅果蝇种群数量在高峰期的时段,正处于气温较高的夏季,同时果园也具备了数量、品种较多的寄主植物,而在种群数量维持在较低水平的其它季节,寄主植物的数量及品种也减少,后续深入探索可参考寄主等因子对斑翅果蝇种群发生的影响研究,例如,墨西哥果实结实季节(8月至次年5月)是影响当地田间斑翅果蝇季节性活动的最主要因素之一(Dominguez-Mongeetal.,2021);相同温度条件下,豇豆蚜虫Aphiscraccivora对于大豆(首选寄主)和棉花(次选寄主)表现不同,即在大豆中的内在增长率、繁殖率、每成年子代数和寿命均显著高于棉花,且温度波动到极端水平时,导致以棉花为食的蚜虫的上述各项参数迅速下降,而以大豆为食的蚜虫则没有下降,在一定程度上解释了夏初新疆棉区气温升高时豇豆蚜虫从棉花向大豆进行生态位转变的现象(Luetal.,2017)。因此,在全球变暖的背景下,除了对害虫进行种群动态建模和风险分析之外,同时也应探索寄主等因子影响害虫如何调节自身应对极端环境,或发生了何种生态位转变的研究。
