β-伴大豆球蛋白通过上调NOD样受体蛋白-3表达诱导猪小肠上皮细胞焦亡
2021-08-09孙智峰刘羽佳李思婷谢维娜单兴根丁红研王希春吴金节
王 蕾 孙智峰 刘羽佳 李思婷 谢维娜 单兴根 田 朋 杨 悦 丁红研 李 玉 王希春 吴金节
(安徽农业大学动物科技学院,合肥 230061)
β-伴大豆球蛋白(β-conglycinin,7S)是大豆中贮藏的一种主要蛋白质,分子质量为140~180 ku,为大豆总蛋白质含量的10.0%~12.7%,由α′亚基、α亚基和β亚基组成三聚体结构,分子质量分别为68、72和52 ku,易引起幼龄动物免疫球蛋白E(IgE)型过敏反应[1]。动物吸收大豆球蛋白的主要部位在小肠,小部分见于胃和大肠中[2-3]。幼龄动物采食β-伴大豆球蛋白后,肠道出现明显炎症浸润,使小肠通透性升高,损坏肠黏膜屏障,引起小肠炎症性疾病,严重损伤机体健康,降低动物生产性能[4-5]。肠上皮细胞(IEC)层暴露于肠腔内容物中,参与营养物质的选择性吸收,并通过相邻IEC之间的紧密连接(TJ)的表达,对肠腔内容物的被动细胞旁通透性起到屏障作用。本课题组前期研究证实,β-伴大豆球蛋白降低猪小肠上皮细胞(IPEC-J2)TJ的表达,提高细胞通透性,降低细胞线粒体膜电位,抑制细胞增殖,引起IPEC-J2损伤和凋亡[6-8];在前期用透射电镜观察β-伴大豆球蛋白引起IPEC-J2损伤和凋亡的过程中,常发现高浓度(10 mg/mL)的β-伴大豆球蛋白组中细胞形态异常肿胀、细胞膜破裂、细胞质流出、线粒体结构异常,细胞形态高度疑似细胞焦亡的外部形态。
细胞焦亡又称细胞炎性坏死,是一种程序性细胞死亡,表现为细胞不断胀大直至细胞膜破裂,导致细胞内容物的释放进而激活强烈的炎症反应。NOD样受体蛋白-3(NLRP-3)是细胞内的一种蛋白质复合体,主要功能是活化半胱氨酸天冬氨酸蛋白酶-1(Caspase-1)进而间接调控白细胞介素-1β(IL-1β)的成熟及分泌,诱导细胞焦亡,引起炎症反应。因此,本试验拟通过构建慢病毒靶向siRNA-NLRP-3干扰载体转染IPEC-J2后再添加10 mg/mL的β-伴大豆球蛋白,分析β-伴大豆球蛋白是否通过NLRP-3/Caspase-1/消皮素D(GSDMD)信号通路引起仔猪小肠上皮细胞焦亡,介导过敏性炎症的发生,有助于认识其在仔猪肠道过敏反应发生发展和转归中的作用,为临床防治β-伴大豆球蛋白诱导的仔猪过敏性肠道疾病提供新思路。
1 材料与方法
1.1 试验材料
β-伴大豆球蛋白购自中国农业大学食品工程学院,并进一步提纯至95%;IPEC-J2由武汉市农业科学院细胞库提供;RPMI 1640培养基购自美国Thermo公司;二辛可宁酸(BCA)蛋白浓度测定试剂盒购自Biosharp;β-肌动蛋白(β-actin)由爱必信(上海)生物科技有限公司提供;辣根过氧化物酶(HRP)标记山羊抗兔免疫球蛋白G(IgG)二抗和山羊抗鼠二抗购自Biosharp;NLRP-3-shRNA慢病毒干扰载体及其阴性对照NC-shRNA慢病毒载体由广州易锦生物技术公司构建合成。
1.2 试验方法
1.2.1 细胞培养与转染
将对数生长期的IPEC-J2以5×105个/孔的密度接种在6孔细胞板,置于37 ℃、5% CO2的恒温培养箱中培养,当细胞融合度达到30%~40%时,随机分为A、B、C组,每组3个重复。A组(对照组,无添加)每24 h更换新鲜培养液。依据预试验参数[感染复数(MOI)=100]进行病毒悬液的感染,B组(阴性对照组)添加慢病毒空载体,C组(干扰组)添加携带NLRP-3干扰片段的慢病毒载体,并加入5 μg/mL的聚凝胺辅助转染,于培养箱内培养12 h后更换新鲜培养液继续培养。72 h后收集各组IPEC-J2,并通过实时荧光定量PCR(RT-qPCR)和蛋白质印迹法(Western blot)验证携带NLRP-3干扰片段的慢病毒载体是否沉默NLRP-3基因。
1.2.2 RT-qPCR、Western blot验证目的基因NLRP-3沉默效果
收集A、B、C组细胞,并加入Trizol试剂提取总RNA,将RNA反转录合成cDNA作为模板,利用PCR仪扩增,以2-ΔΔCt法计算各组细胞中NLRP-3 mRNA相对表达量。NLRP-3 PCR引物序列为F:5′-GACCTCAGCCAAGATGCAA-3′,R: 5′-GACCTCAGCCAAGATGCAA-3′;甘油醛-3-磷酸脱氢酶(GAPDH)PCR引物序列为F:5′-TGACCCCTTCATTGACCTCC-3′,R:5′-CCATTTGATGTTGGCGGGAT-3′。
收集A、B、C组细胞,进行Western blot检测。每个聚丙烯酰胺凝胶电泳(SDS-PAGE)(12%)泳道加入20 μg的总蛋白电泳并转移至聚偏二氟乙烯膜(PVDF)膜上。PVDF膜4 ℃下与一抗孵育过夜后用Tris-HCl缓冲盐溶液+吐温(TBST)溶液清洗3次,进行二抗孵育。洗膜3次,放入凝胶成像系统显影拍摄,以β-actin作为内参蛋白校正试验误差。
1.2.3 试验分组与设计
依据RT-qPCR和Western blot结果得知转染携带NLRP-3干扰片段的慢病毒载体成功沉默目的基因NLRP-3后,将对数生长期的IPEC-J2以5×105个/孔的密度接种在6孔细胞板,置于37 ℃、5% CO2的恒温培养箱中培养。当细胞融合度达到30%~40%时,随机将细胞分为4组:A组为对照组(未转染),B组为阴性对照组(转染慢病毒空载体),C组为干扰组(转染携带NLRP-3干扰片段的慢病毒载体),D组为β-伴大豆球蛋白调节组(转染携带NLRP-3干扰片段的慢病毒载体后添加10 mg/mL的β-伴大豆球蛋白,继续培养24 h),每组设置3个重复,转染方法参照1.2.1。
1.2.4 细胞活性测定
按照6 000个/孔的密度将对数生长期中的IPEC-J2悬液(100 μL/孔)接种在96孔板中,置于37 ℃、5% CO2的恒温培养箱中培养。当细胞融合度达到30%~40%时,随机分为A、B、C、D组,按照试验分组与设计将B、C和D组分别进行慢病毒转染(MOI=100)处理后,在D组添加10 mg/mL的β-伴大豆球蛋白。继续培养24 h后,每孔加入10 μL的细胞增殖毒性检测试剂盒溶液,在细胞培养箱内继续孵育至出现明显的颜色反应。用酶标仪测定450 nm处的光密度(OD)值,检测细胞活性。
1.2.5 末端标记法(TUNEL)染色法观测细胞阳性表达
收集A、B、C和D组细胞用10%中性甲醛固定,按照脱氧核糖核苷酸末端转移酶介导的缺口TUNEL检测试剂盒说明书检测细胞死亡方式。采用红色荧光标记TUNEL阳性细胞,4,6-联脒-2-苯基吲哚(DAPI)染色细胞核,荧光显微镜下观察并采集图像。
1.2.6 Caspase-1和IL-1β含量测定
收集A、B、C和D组细胞至离心管,1 000 r/min离心5 min,磷酸盐缓冲液(PBS)洗涤3次。随后在每组样品中加入500 μL含0.1% TritonX-100的0.1 mol/L Tris-HCl(pH=7.4),冰水中超声裂解各组样品。收集细胞裂解液,1 000 r/min离心10 min后,吸取上清液,依据ELISA试剂盒说明进行试验,检测Caspase-1和IL-1β含量。
1.2.7 透射电子显微镜观察细胞形态
收集A、B、C和D组细胞至离心管,1 000 r/min离心5 min,弃上清液,每管加入1 mL的4%多聚甲醛4 ℃下固定12 h后,PBS洗涤3次(4 ℃,5 000×g,15 min),之后用2%锇酸渗透酸固定4 h。PBS冲洗后,用30%~100%酒精梯度脱水,渗透包埋,切割超薄切片染色,JEM-1230透射电子显微镜观察并拍摄图片。
1.2.8 RT-qPCR检测NLRP-3、ASC、Caspase-1、IL-1β和GSDMDmRNA相对表达量

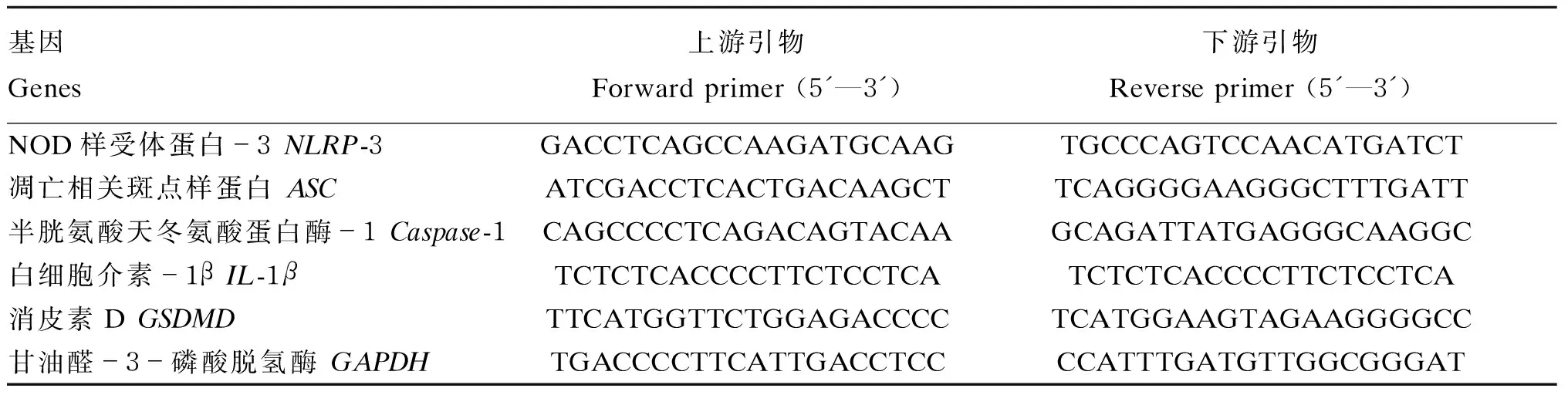
表1 基因引物序列参数
1.2.9 Western blot检测NLRP-3、ASC、Caspase-1、IL-1β和GSDMD蛋白表达水平
收集A、B、C和D组细胞,进行Western blot检验,以β-actin作为内参蛋白校正试验误差。
1.3 数据处理
试验数据用平均值±标准差表示,采用SPSS 17.0软件的ANOVA进行方差分析,最小显著差异(LSD)法进行显著性比较,P<0.05表示差异显著,P<0.01表示差异极显著。使用Graph Pad Prism 7.0软件绘制柱状图,使用Image J软件分析Western blot蛋白条带灰度值。
2 结果与分析
2.1 慢病毒转染IPEC-J2后成功沉默NLRP-3的表达
如图1、图2所示,与A组相比,B组细胞中NLRP-3 mRNA相对表达量和蛋白表达水平无显著差异(P>0.05),C组细胞中的NLRP-3 mRNA相对表达量和蛋白表达水平均极显著降低(P<0.01);与B组相比,干扰组细胞中的NLRP-3 mRNA相对表达量和蛋白表达水平均极显著降低(P<0.01)。结果表明,慢病毒转染IPEC-J2后成功沉默目的基因NLRP-3。
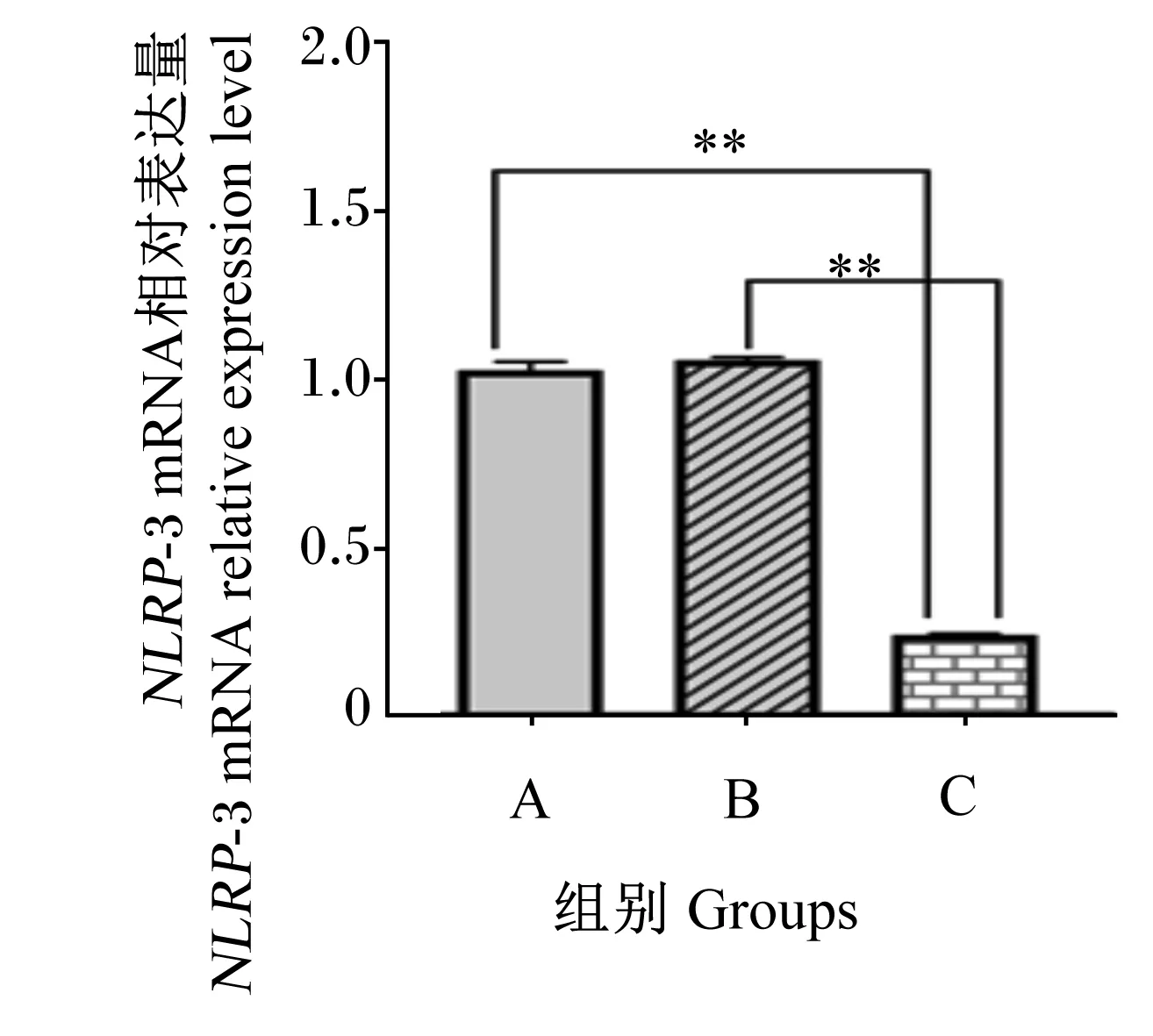
*表示与A组相比差异显著(P<0.05),**表示与A组相比差异极显著(P<0.01)。下图同。

图2 Western blot检测NLRP-3蛋白表达水平
2.2 β-伴大豆球蛋白降低IPEC-J2的细胞活性
如图3所示,与A组相比,B、C组细胞活性无显著差异(P>0.05);与B组相比,C组细胞活性无显著差异(P>0.05);与C组相比,D组细胞活性极显著下降(P<0.01)。
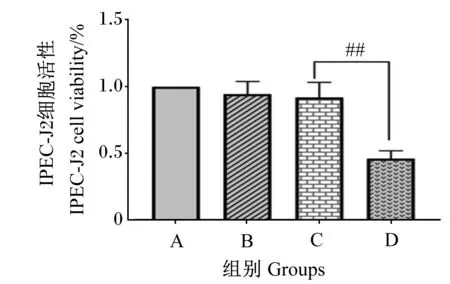
#表示与C组相比差异显著(P<0.05),##表示与C组相比差异极显著(P<0.01)。下图同。
2.3 β-伴大豆球蛋白促进IPEC-J2的阳性表达
如图4所示,与A组细胞相比,B、C组细胞无明显变化。与B组细胞相比,C组细胞无明显变化。与C组细胞对比,D组细胞密度降低,数量明显减少,细胞阳性表达明显增强。
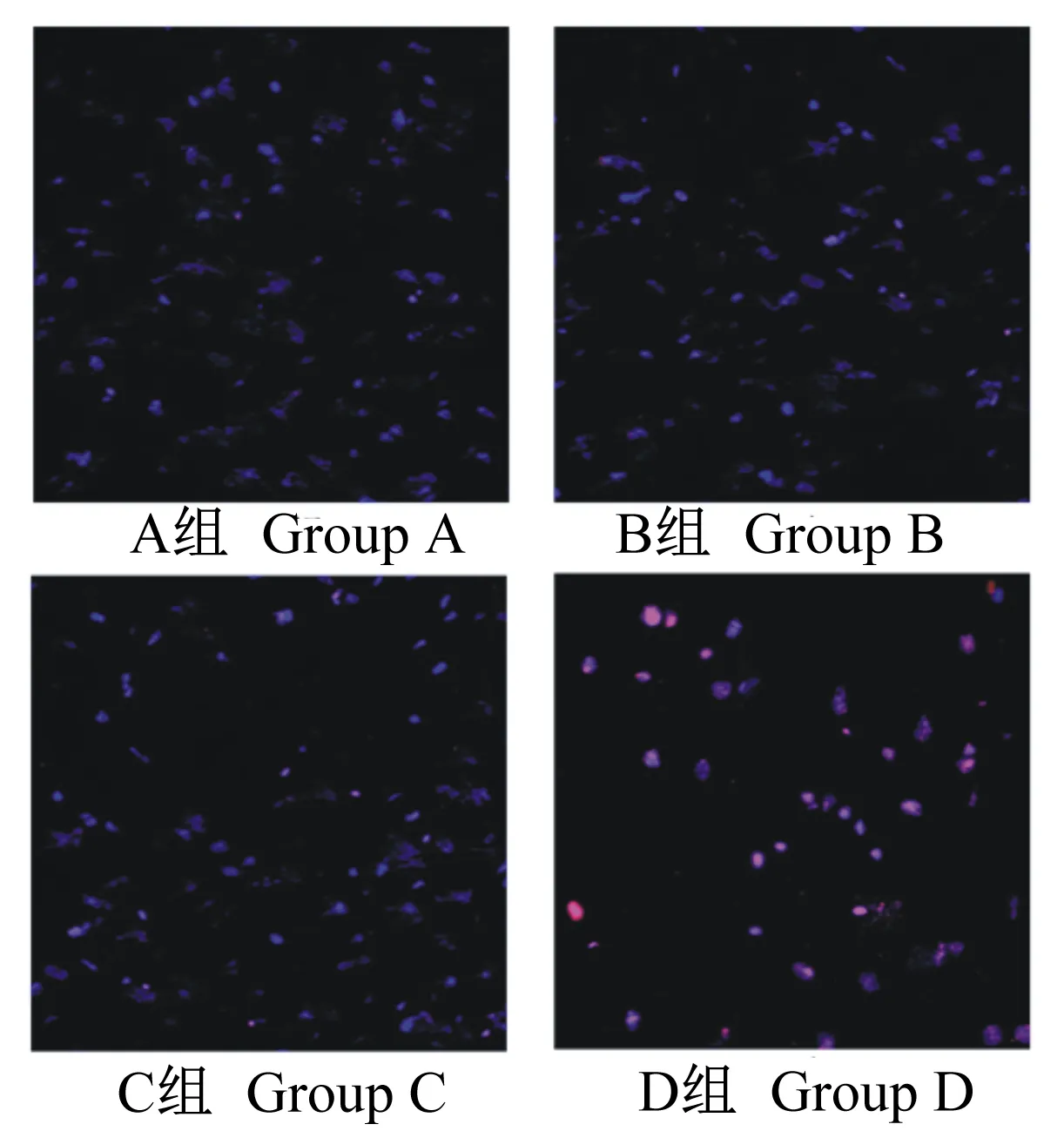
图4 TUNEL染色法观察各组IPEC-J2阳性表达
2.4 β-伴大豆球蛋白上调IPEC-J2细胞内Caspase-1和IL-1β的含量
如图5所示,与A组相比,B组细胞Caspase-1和IL-1β含量无显著差异(P>0.05),C组细胞Caspase-1含量显著降低(P<0.05),IL-1β含量极显著降低(P<0.01);与B组相比,C组细胞Caspase-1和IL-1β含量显著降低(P<0.05);与C组相比,D组细胞Caspase-1和IL-1β含量极显著升高(P<0.01)。
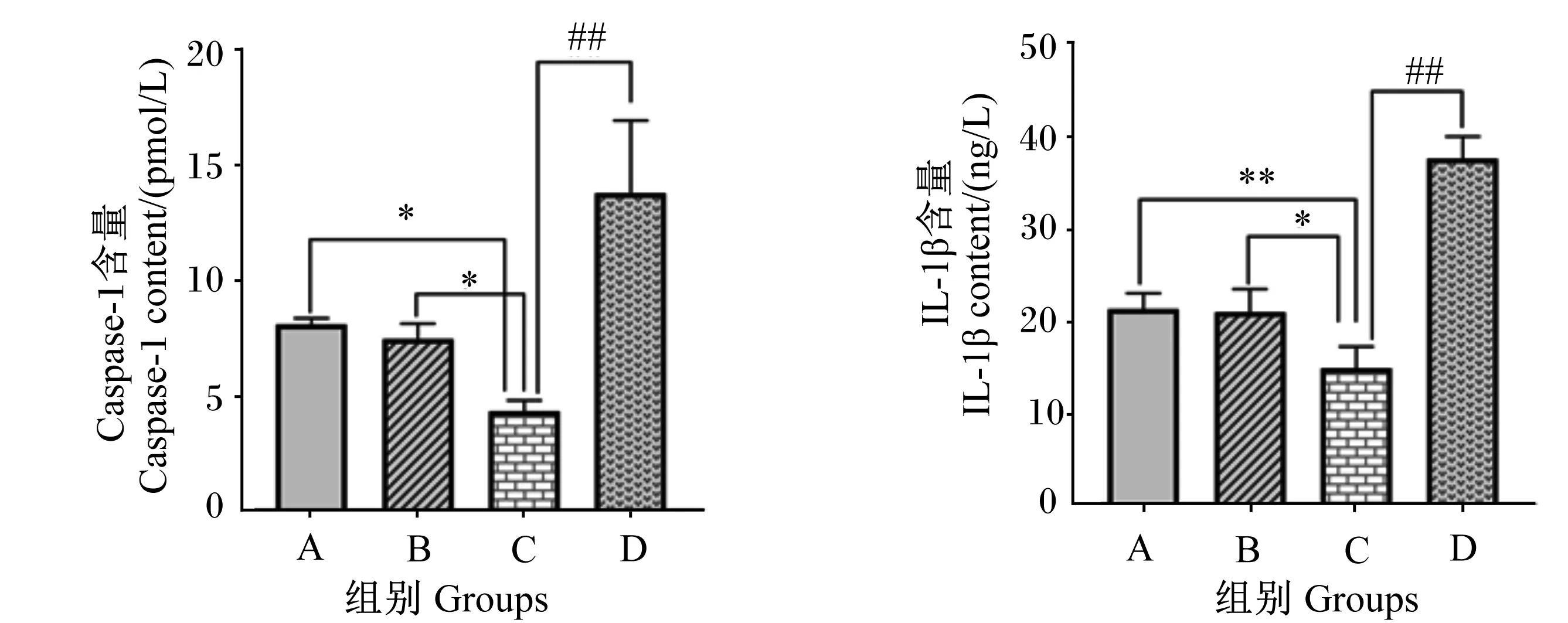
图5 β-伴大豆球蛋白对IPEC-J2细胞Caspase-1和IL-1β含量的影响
2.5 β-伴大豆球蛋白损伤IPEC-J2细胞的内部结构
如图6所示,与A组相比,B、C组细胞内部结构无明显变化;与B组相比,C组细胞内部结构无明显变化;与C组相比,D组细胞形态肿胀,细胞膜破裂出现孔洞、胞质疏松、胞质流出细胞膜,线粒体变性肿胀。
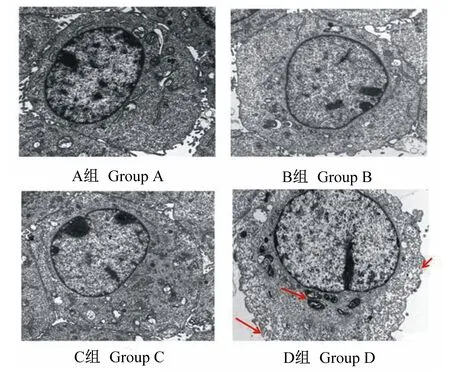
红色箭头所示:线粒体变性肿胀,细胞膜破裂出现孔洞、胞质流出。
2.6 β-伴大豆球蛋白上调IPEC-J2细胞NLRP-3、ASC、Caspase-1、IL-1β和GSDMD mRNA相对表达量
如图7所示,与A组相比,B组细胞NLRP-3、ASC、Caspase-1、IL-1β和GSDMDmRNA相对表达量无显著差异(P>0.05);C组细胞NLRP-3、Caspase-1和GSDMDmRNA相对表达量极显著降低(P<0.01),ASC和IL-1βmRNA相对表达量显著降低(P<0.05)。与B组相比,C组细胞IL-1βmRNA相对表达量显著降低(P<0.05),NLRP-3、ASC、Caspase-1和GSDMDmRNA相对表达量极显著降低(P<0.01);与C组相比,D组细胞NLRP-3、ASC、Caspase-1、IL-1β和GSDMDmRNA相对表达量极显著升高(P<0.01)。
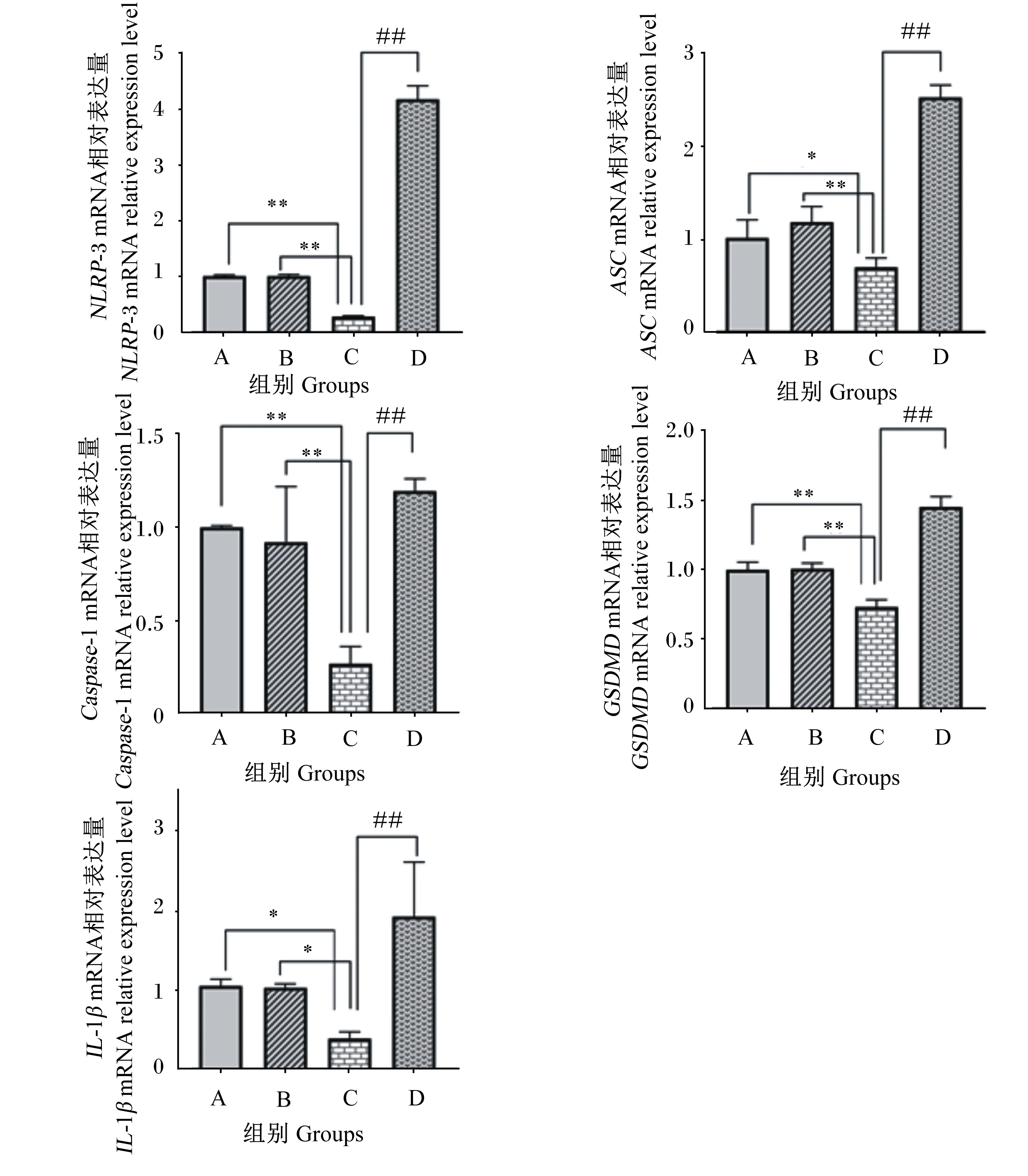
图7 β-伴大豆球蛋白对IPEC-J2中NLRP-3、ASC、Caspase-1、IL-1β和GSDMD mRNA相对表达量的影响
2.7 β-伴大豆球蛋白上调IPEC-J2细胞NLRP-3、ASC、Caspase-1、IL-1β和GSDMD蛋白的表达水平
如图8所示,与A组相比,B组细胞NLRP-3、ASC、Caspase-1、IL-1β和GSDMD蛋白表达水平无显著差异(P>0.05);C组细胞NLRP-3、ASC、Caspase-1、GSDMD和IL-1β蛋白表达水平极显著降低(P<0.01)。与B组相比,C组细胞Caspase-1蛋白表达水平显著降低(P<0.05),NLRP-3、ASC、GSDMD和IL-1β蛋白表达水平极显著降低(P<0.01)。与C组相比,D组细胞NLRP-3、ASC、Caspase-1、GSDMD和IL-1β蛋白表达水平极显著升高(P<0.01)。
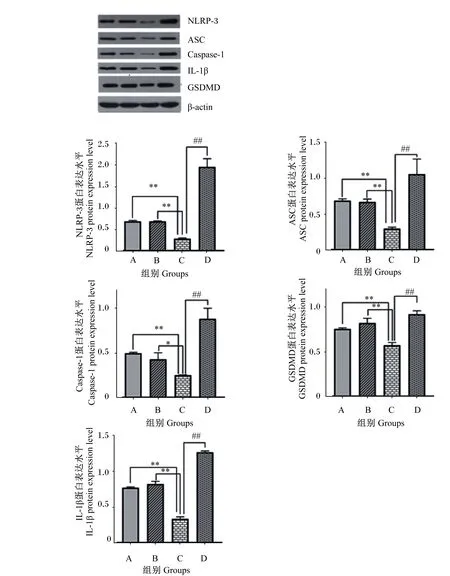
NLRP-3:NOD样受体蛋白-3 nod-like receptor pyrin domain-3;ASC:凋亡相关斑点样蛋白 apoptosis-associated speck-like protein;Caspase-1:半胱氨酸天冬氨酸蛋白酶-1 cysteine-containing aspartate-specific proteases-1;IL-1β:白细胞介素-1β interleukin-1β;GSDMD:消皮素D gsdermin D。
3 讨 论
消化免疫系统尚未发育完善的仔猪采食β-伴大豆球蛋白后,小部分未分解的β-伴大豆球蛋白通过肠上皮细胞进入机体,引发由特异性抗体IgE介导的急性过敏反应,提高肠道通透性,损伤肠黏膜[1]。IPEC-J2位于仔猪小肠第1层,在构成完整的肠道管腔环境过程中发挥重要作用。研究发现,β-伴大豆球蛋白破坏IPEC-J2细胞骨架和紧密连接蛋白,降低IPEC-J2活性,抑制细胞生长增殖,诱导细胞凋亡[8-9]。IECs焦亡是区别于细胞凋亡、铁死亡、坏死和自噬的炎症性细胞死亡形式,并在急、慢性肠道损伤的发病过程中发挥关键作用[10]。NLRP-3是广泛存在于上皮细胞的炎性复合体,是引发细胞焦亡的关键因子[11-12],被激活后通过其N端的热蛋白结构域(PYD)与凋亡相关斑点样蛋白(ASC)的PYD域相连,诱导ASC的胱天蛋白酶募集结构域(CARD)招募并激活Caspase-1[13-15]。激活的Caspase-1切割GSDMD蛋白引发细胞焦亡[16]。
NLRP-3/Caspase-1在过敏性疾病的发生发展过程中发挥重要作用[17-18]。本试验利用慢病毒转染IPEC-J2后,与A组相比,B组细胞的NLRP-3 mRNA相对表达量和蛋白表达水平无显著差异,而C组细胞的NLRP-3 mRNA相对表达量和蛋白表达水平极显著降低,结果表明转染慢病毒空载体对IPEC-J2的NLRP-3 mRNA相对表达量无显著影响,而携带NLRP-3干扰片段的慢病毒载体转染IPEC-J2可以显著提高基因的转染效率并能够成功沉默目的基因NLRP-3。利用RT-PCR和Western blot验证慢病毒转染成功沉默目的基因NLRP-3后将样本分为4组(A、B、C、D组)的试验中,B组细胞的Caspase-1含量以及NLRP-3、ASC、Caspase-1 mRNA相对表达量和蛋白表达水平与A组相比无显著差异;C组细胞的Caspase-1含量以及NLRP-3、ASC和Caspase-1 mRNA相对表达量和蛋白表达水平显著低于A组及B组细胞;这说明沉默IPEC-J2的NLRP-3基因后,可以显著下调NLRP-3通路下游的ASC和Caspase-1基因的表达量。与C组相比,D组细胞(在沉默NLRP-3基因的IPEC-J2培养液中加入10 mg/mL的β-伴大豆球蛋白)的Caspase-1含量显著升高,NLRP-3、ASC和Caspase-1 mRNA相对表达量和蛋白表达水平显著升高。本试验结果说明,10 mg/mL的β-伴大豆球蛋白上调IPEC-J2NLRP-3的表达,上调的NLRP-3通过炎性复合体衔接蛋白ASC招募并激活Caspase-1。Zhang等[19]用卵蛋白诱导小鼠建立过敏性呼吸道炎症模型发现,致敏组小鼠NLRP-3、ASC和Caspase-1 mRNA相对表达量和蛋白表达水平显著上升;加入NLRP-3特异性抑制剂后NLRP-3、ASC和Caspase-1 mRNA相对表达量和蛋白表达水平显著升高下降,这与本试验结果相符合。
GSDMD是细胞焦亡的执行蛋白,被活化的Caspase-1切割后,释放其具有结合膜磷脂上膜打孔活性的N端结构域,在细胞膜上形成活性的孔隙,使得水分子等物质进入细胞内而引起细胞肿胀、细胞膜破裂,导致细胞焦亡[20-21]。Gan等[22]研究发现,NLRP-3特异性抑制剂可以抑制GSDMD的活化,并抑制IL-1β的过度释放,缓解细胞焦亡。本试验中,C组细胞IL-1β含量,GSDMD和IL-1βmRNA相对表达量和蛋白表达水平均显著低于A、B组细胞,说明沉默IPEC-J2的NLRP-3基因能够显著抑制GSDMD和IL-1β的表达,这与Liu等[23]通过敲除NLRP3基因显著减少IL-1β分泌和抑制呼吸道上皮细胞焦亡的结果一致。Song等[24]研究表明,Caspase-1切割GSDMD蛋白后释放炎性介质IL-1β,进一步级联扩大炎症反应,导致细胞焦亡和肠组织损伤。本试验中,与C组相比,D组细胞IL-1β含量,GSDMD和IL-1βmRNA相对表达量和蛋白表达水平显著升高,细胞活性显著降低,TUNEL染色IPEC-J2呈阳性表达,透射电镜观察可见细胞膜出现孔洞、细胞肿胀裂解、细胞质外溢、线粒体肿胀病变、线粒体嵴消失,细胞外部形态及内部结构呈现典型的细胞焦亡状态。Kim等[25]发现NLRP-3炎性小体的激活与过敏性重症/难治性哮喘的发生显著相关,进一步研究发现尘螨等过敏源通过NLRP-3/Caspase-1/GSDMD信号通路诱导人支气管上皮细胞焦亡,本试验结果与此一致。这表明β-伴大豆球蛋白显著上调NLRP-3 mRNA相对表达量,上调的NLRP-3激活其下游的Caspase-1/GSDMD信号通路,进而过度释放炎性介质IL-1β,诱导IPEC-J2焦亡。
4 结 论
β-伴大豆球蛋白通过上调IPEC-J2的NLRP-3表达,激活NLRP-3/Caspase-1/GSDMD信号通路,诱导IPEC-J2焦亡。
