增温对鼎湖山混交林中4种优势树种生物量分配和养分积累的影响
2021-08-09谭钠丹李旭吴婷列志旸刘旭军刘世忠陈平刘菊秀
谭钠丹, 李旭, 吴婷, 列志旸, 刘旭军, 刘世忠, 陈平, 刘菊秀
增温对鼎湖山混交林中4种优势树种生物量分配和养分积累的影响
谭钠丹1,2, 李旭1, 吴婷1, 列志旸1, 刘旭军1, 刘世忠1, 陈平2, 刘菊秀1*
(1. 中国科学院华南植物园退化生态系统植被恢复与管理重点实验室,广州 510650;2. 仲恺农业工程学院,广州 510225)
为了解未来全球变暖对南亚热带森林生态系统物种组成的影响,在广东鼎湖山采用沿海拔梯度垂直移位的方法,研究了模拟增温对木荷()、红枝蒲桃()、红锥()和马尾松()等4种优势树种的生物量分配模式和养分(氮和磷)积累的影响。结果表明,增温使大气平均温度增加(1.28±0.60)℃,土壤平均温度增加(1.04±0.30)℃;6 a长期增温使木荷和马尾松的树高增长率分别显著提高83.0%和52.1%,基径增长率分别显著提高37.1%和76.9%,二者的茎质比都显著增加,根质比、叶质比和根冠比都显著降低,养分积累量分别显著增加100.3%和185.7%;但增温对红枝蒲桃和红锥的树高、基径、生物量分配模式和养分积累无显著影响。因此,6 a增温对4种优势树种生物量分配模式和养分积累的影响具有一定的种间差异,木荷和马尾松由于生物量和养分积累量较高,所以在长期增温条件下可能具有较强的适应性,这种差异可能会对我国南亚热带混交林中的群落结构和功能带来潜在影响。
移位增温; 生物量分配格局; 植物养分积累;鼎湖山
随着大量化石燃料的燃烧和土地使用的变化,温室气体的浓度急剧增加,导致大气温度显著升高,全球气温较工业革命前已经增加1℃[1–3]。其中热带和亚热带地区增温幅度为1.3℃~5.0℃,显著高于全球平均水平[4]。森林作为全球陆地生态系统的主体,具有分布广、生产力高及固碳能力强等特点,在陆地碳循环中发挥着重要作用。热带森林的碳储量和净初级生产力在陆地生态系统中的占比更是高达25%和33%,它们拥有丰富的生物多样性,并提供广泛的生态系统服务[5–6]。相比于高纬度森林,低纬度森林植被热适应的生态位相对狭窄,由于热带树种可能更容易受到持续变暖的影响,因此对未来全球变暖的适应能力可能远小于高纬度地区[7–8]。全球气候变暖必然会对热带森林生态系统造成巨大威胁,因而亟需在热带及亚热带地区开展增温试验[9–11]。
各器官之间的生物量分配模式,即叶质比(LMR)、茎质比(SMR)、根质比(RMR)和根冠比(R/S)可以反映植物对资源获取和利用之间的协调配比[12–13]。研究表明,增温会改变森林土壤中养分有效性和植物光合速率,进而影响植物生物量积累和植物对养分的吸收及分配[14–17]。Yasuaki等报道,当温度升高时,植物的热适应能力和生物量分配模式的可塑性有助于维持其碳平衡[18]。有研究表明, 植物可通过改变生物量分配模式来提高获取各种资源的能力,以满足在增温条件下增长的生长速率[19–20]。不同植物的生物量变化对增温的响应也不同,增温显著增加了山油麻()的树高和生物量[21];曾贞等[22]报道增温不改变桑树()幼苗的生物量;也有研究表明,增温会降低植物生物量[23]。增温对植物生物量的影响还与增温时间有关,短期增温和长期增温对植物生物量的影响不一致[24–27]。养分积累量与植物自身的生长状况紧密相关,是植物生长潜力的重要指标[28–29]。由于气候变暖而引起的植物生物量和养分吸收能力的变化将直接影响其养分积累量[30–32],从而进一步影响植物生长。目前有关植物养分积累量对增温响应机理的研究相对较少,且多数是基于中高纬度地区和农田[33–35],给研究结果带来很大的限制性,前期的研究结果能否应用到低纬度森林中还无法验证。
我国亚热带森林被称为“北回归线荒漠带上的绿洲”,分布着世界上现存面积最大、最典型的常绿阔叶林。针阔叶混交林是我国南亚热带针叶林向地带性常绿阔叶林演替的中间林分类型,也是我国南亚热带地区主要林分类型,其中木荷()、红枝蒲桃()、红锥()和马尾松()等为优势种。本文通过垂直移位增温试验探究长期增温对这4种南亚热带优势树种生物量分配和养分积累量变化的影响,并试图揭示植物生物量分配与其养分积累量对增温的响应机制,为预测未来南亚热带混交林生产力及其群落结构提供参考。
1 研究区概况
研究区位于广东省中部鼎湖山森林生态系统定位研究站(112°30′39″~112°33′41″ E,23°09′21″~ 23°11′30″ N),为典型的亚热带季风气候。年平均气温为20.9℃。年降水量为1 927 mm, 降雨多集中于3-8月份。针阔叶混交林为鼎湖山海拔300 m处的主要林型。土壤主要类型为赤红壤,土层厚40~80 cm, 土壤pH值为4.6,表土层有机质含量为4.3%[36]。
2 材料和方法
2.1 试验地设计
我们选择位于鼎湖山海拔300 m的五棵松样地的针阔叶混交林为研究对象,2012年1月,在海拔300 m处的五棵松混交林中和30 m处的季风林旁的苗圃地,分别选择30 m×30 m区域,去除区域内乔木层和灌木层,使该空旷地带暴露于充足的阳光中。在2个海拔的空旷地上分别修建3个OTC (open-top chambers)(长×宽×高=3 m×3 m×0.8 m)。OTC样方地下部分四周及底部铺上水泥和瓷砖,防止隔离样方内土壤与周围土壤的水分和养分交换(图1)。

图1 样方构造示意图
每个OTC的土壤分3层(5、20和40 cm层)观测土壤温度(109, Campbell Scientific, Lincoln, USA)和湿度(CS616, Campbell Scientific, Lincoln, USA)。在样地旁的平整地上架设离地2 m高度的地面气象观测系统,观测大气温度和湿度(HMP155A, Vaisala, Helsinki, Finland)[37]。
2012年4月,从位于海拔约300 m的混交林中分别收集不同层次(0~20、20~40、40~70 cm)的土壤,分层混匀后填埋于海拔300和30 m的所有OTC内。土壤填埋后,根据植物在混交林中的分布比例和范围,选取1 a生, 基茎和树高基本一致的6种混交林常见树苗,包括木荷、红枝蒲桃、红锥、马尾松、短序润楠()和山血丹(),每种植物选择6株树苗,先在混交林中的统一位置进行驯化,再移植进填埋了混交林的土壤OTC内,并且按照相同株行距随机定植。由于短序润楠和山血丹生长缓慢,叶片稀少,不足以达到叶片采样的条件。故本研究只选用木荷、红枝蒲桃、红锥和马尾松4种南亚热带混交林优势乔木树种[38]。
2.2 生物量分配和养分积累量的测定
2014年12月、2015年12月(3 a短期增温)和2018年6月(6 a长期增温)分别测量每个OTC内所有植株的基径(mm)和高度(cm),每个OTC中采用全收割法, 每种植物随机收割1棵,以减少树种间的遮蔽和竞争,同一OTC中收割植物的位置基本一致,并将所有砍伐的植株分成根、茎和叶。将采集的植物器官样品在65℃下烘干,测定干质量,根据经验公式=a(2)b估算每株幼苗的生物量,其中为生物量,为基径,为树高,a和b是回归系数(表1)。
生物量分配的计算公式:根质比(RMR)=根生物量/总生物量;茎质比(SMR)=茎生物量/总生物量;叶质比(LMR)=叶生物量/总生物量;根冠比(R/S)=根生物量/(叶生物量+茎生物量)。
用Kjeldahl[37]方法测量叶、茎和根的N浓度; 样品用H2SO4-H2O2消煮后,用光度法测量叶、茎和根的P浓度。单株N积累量(g)=叶N浓度×叶生物量+茎N浓度×茎生物量+根N浓度×根生物量;单株P积累量(g)=叶P浓度×叶生物量+茎P浓度×茎生物量+根P浓度×根生物量。
2.3 数据分析
用SPSS 20.0 (IBM)统计软件对数据进行统计分析,采用Sigmaplot 12.5 (Systat Software Inc)软件绘制图表。采用多因素方差分析法(Multi-Way repeated measures ANOVA)分析数据间的差异显著性;采用单因素方差分析法(One-Way ANOVA)分析增温与对照间的差异。显著性水平设定为=0.05。

表1 增温处理植物的生物量估算方程
3 结果和分析
3.1 增温对大气温度、土壤温度和含水量的影响
从图2可见,2014年1月至2018年11月,增温和对照样地的大气平均温度分别为23.15℃和21.87℃,增温样地升高了(1.28±0.60)℃。增温处理下3个层次的土壤平均温度为22.35℃,对照土壤为21.31℃,增温样地升高了(1.04±0.30)℃。增温样地土壤平均湿度为17%,对照样地为22%,增温样地下降了5%。
3.2 增温对基径和树高的影响
4种植物的基径与株高受到增温、物种、测定年份的影响,温度对植物基径与树高的影响也与树种有关(表2,<0.05)。3 a的短期增温下,4种植物的基径和树高都增加,马尾松基径比对照显著增加了87.0%和87.5%,树高显著增加65.7%和49.8%;木荷的基径在增温第2年显著增加了33.3%,树高在第2和3年显著增加了61.1%和65.1%。在6 a长期增温下,与对照组相比,木荷的基径和树高分别显著增加了37.1%和83.0%,马尾松的基径和树高分别显著增加了76.9%和52.1%;而红锥的树高和基径都有减少的趋势,红枝蒲桃的基径也减少(表3)。
3.3 增温对生物量分配模式的影响
结果表明(表2),增温、年份及其交互作用对植物的总生物量均产生显著影响(<0.05)。从图3可见,2014、2015和2018年木荷总生物量分别比对照显著增加了93.8%、73.7%和100.7%;马尾松分别显著增加了395.2%、341.1%和277.3% (<0.05)。但增温6 a的红锥和红枝蒲桃则分别下降了32.4%和18.9%。
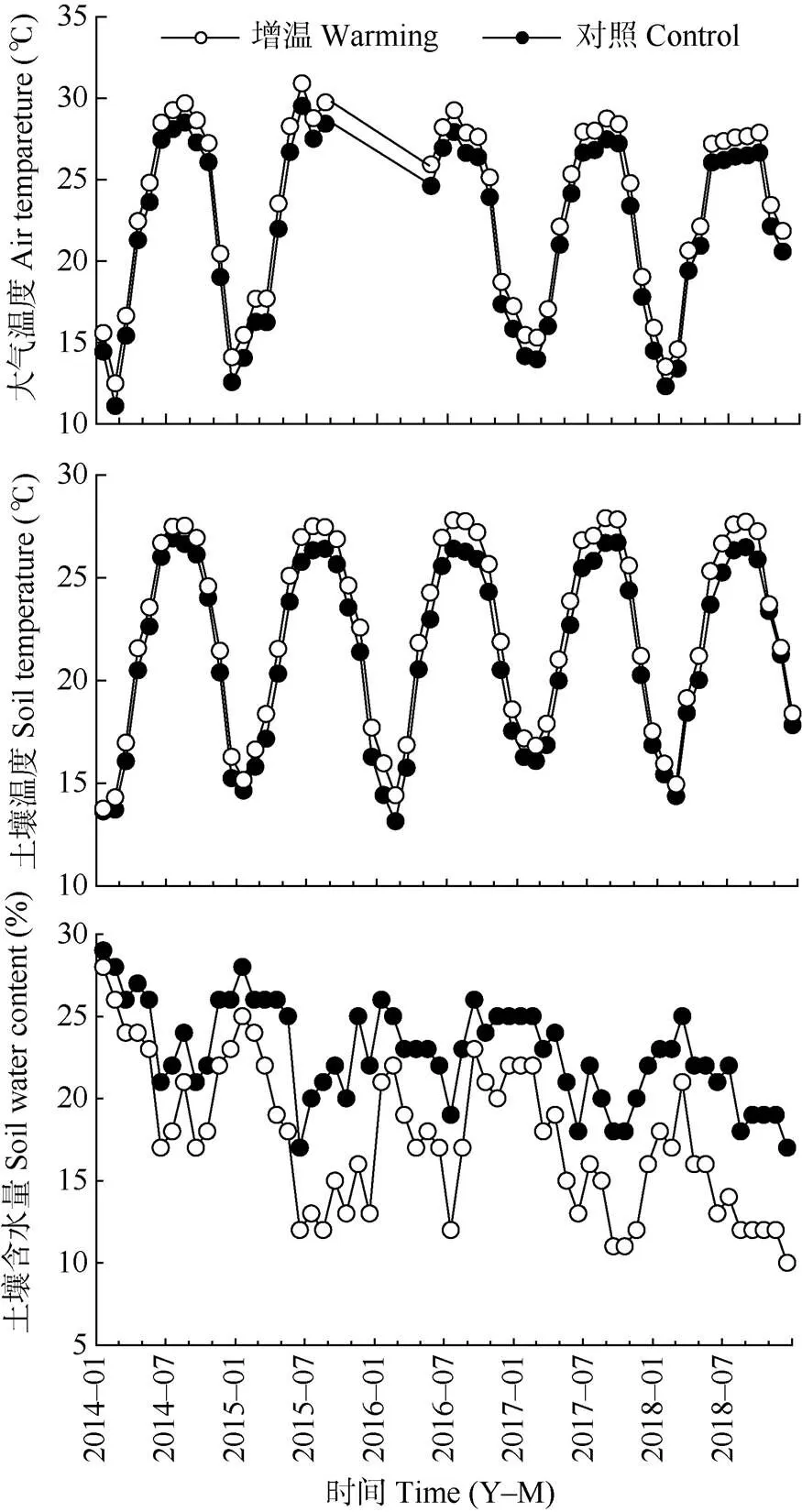
图2 样地大气温度、土壤温度和土壤湿度的动态变化
增温使植物各器官生物量分配模式发生显著变化(表2,<0.05)。增温2、3和6 a,木荷的根质比分别显著下降了7.4%、7.7%和12.5%,叶质比显著下降了15.6%、13.8%和27.1%,茎质比显著上升了17.1%、13.3%和15.7%;马尾松的根质比显著下降了16.7%、18.8%和14.3%,叶质比显著下降了4.2%、4.3%和7.7%,茎质比显著上升了8.8%、8.3%和6.3% (图4,<0.05);短期增温,红枝蒲桃与红锥的根质比和叶质比都呈下降趋势,但增温6 a, 红锥的根质比升高。
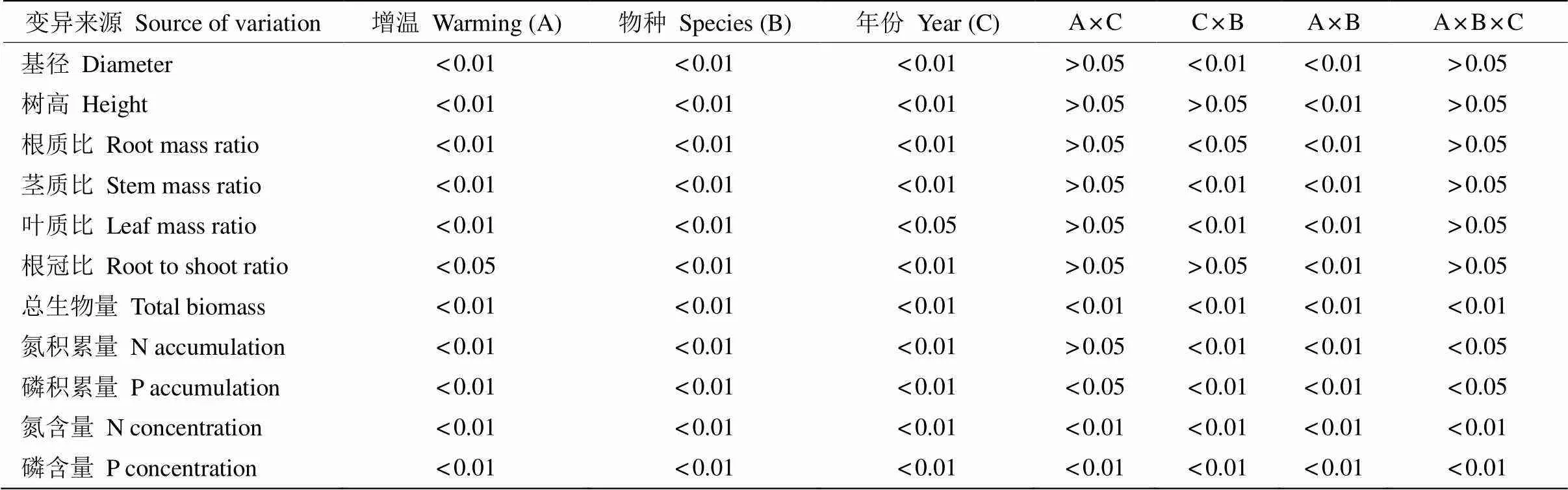
表2 增温对植物生物量分配和养分积累量影响的多因素方差分析(P)
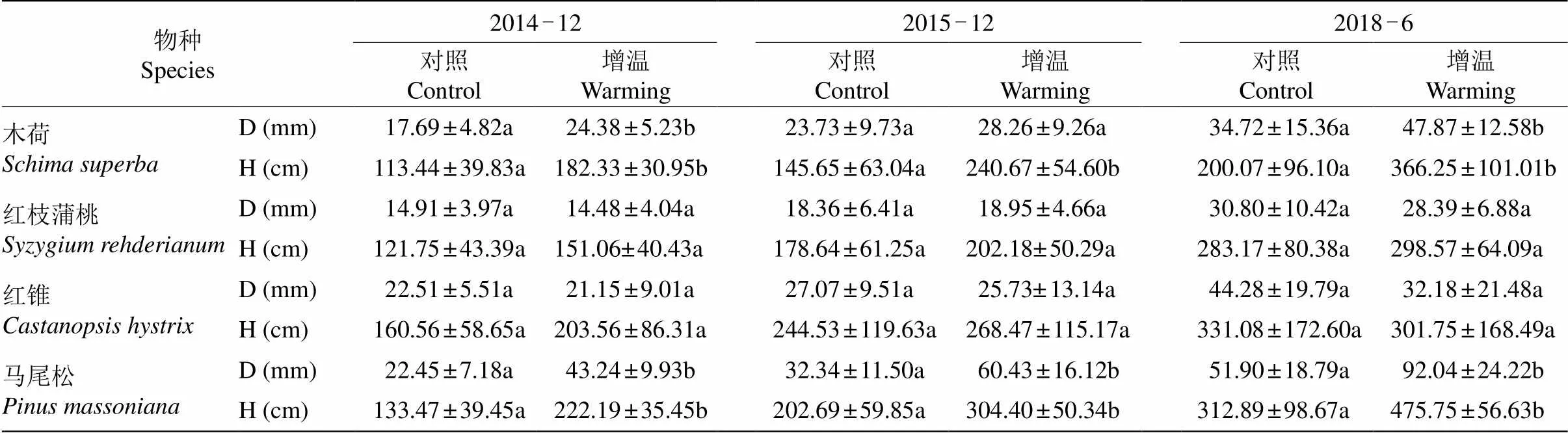
表3 增温对植物基径和树高的影响
=6; D: 基径; H: 树高; 相同时间同行数据后不同字母表示差异显著(<0.05)。下表同。
=6; D: Diameter; H: Height; Data followed different letters at the same line and time indicated significant difference at 0.05 level. The same is following Tables.
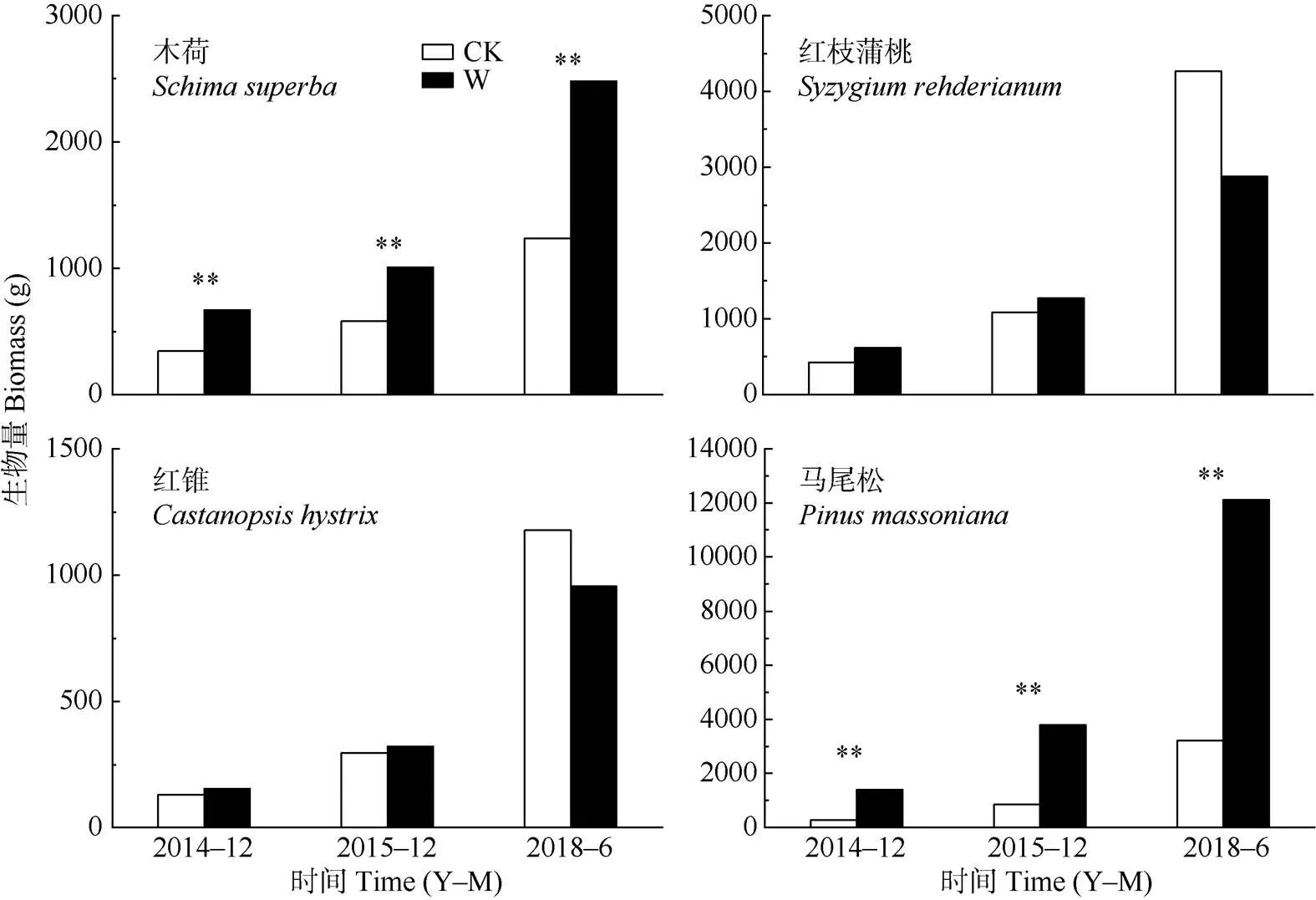
图3 增温对4种树种生物量的影响。*: P<0.05; **:P<0.01; CK: 对照; W: 增温。
根冠比受增温、物种及其二者间的交互效应影响,也与年份有关(表2,<0.05)。增温2、3和6 a,木荷的根冠比分别显著降低了10.8%、11.4%和12.9 %;马尾松显著降低了21.7%、23.8%和23.5% (图4,<0.05);增温6 a红锥的根冠比有所增加。
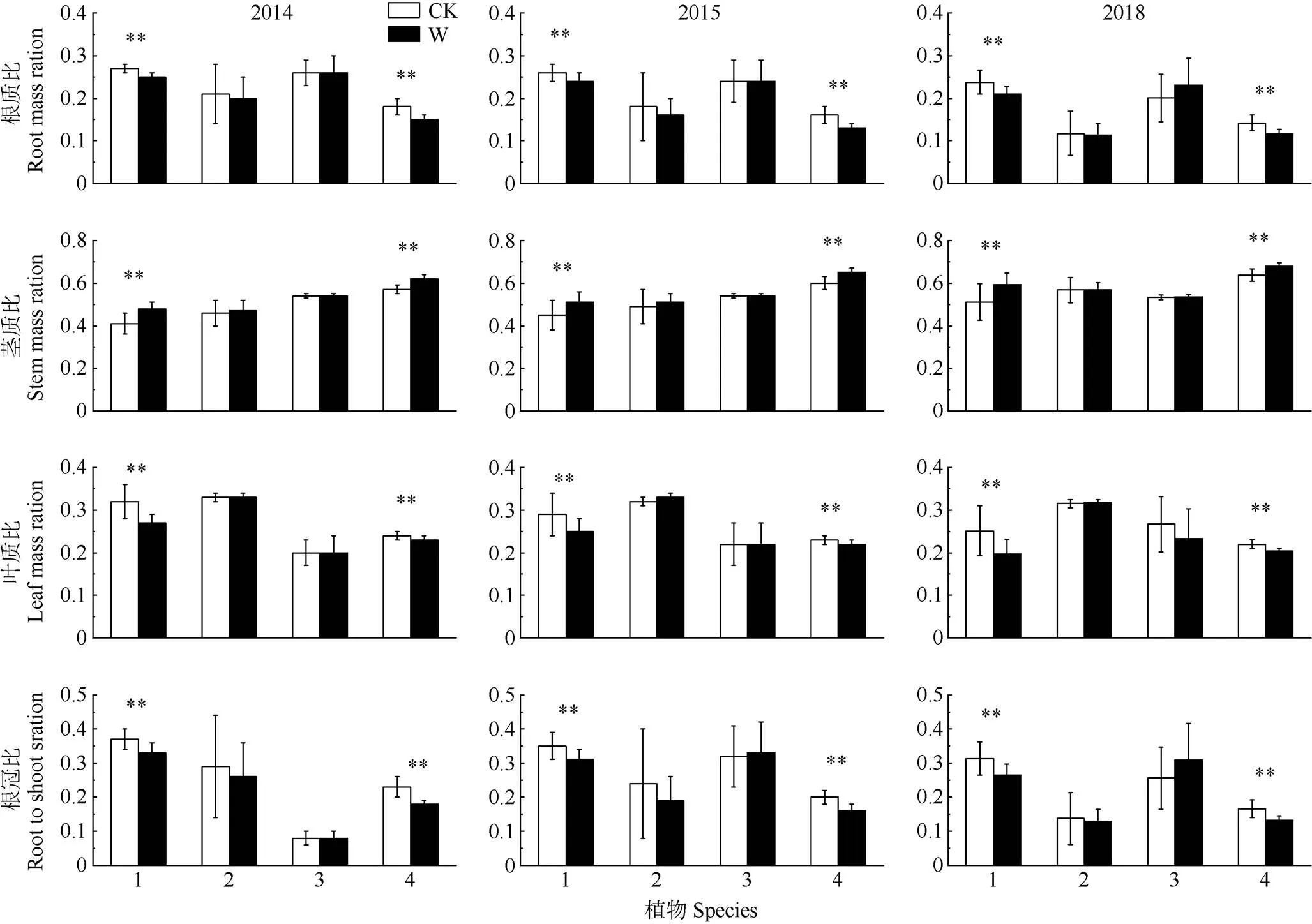
图4 增温对4种树种生物量分配的影响。1: 木荷; 2: 红枝蒲桃; 3: 红锥; 4: 马尾松; *: P<0.05; **: P<0.01; CK: 对照; W: 增温。下图同。
3.4 增温对N、P浓度及其积累量的影响
4种植物的N和P浓度受增温、年份及其交互作用的影响(表2,<0.05)。增温处理2 a,木荷、红枝蒲桃和红锥的N浓度分别比对照显著降低了21.3%、24.0%和28.7%,除木荷外,其他植物的P浓度都增加;增温处理3 a,木荷、红枝蒲桃和马尾松的P浓度分别比对照显著增加了41.8%、39.5%和26.0%;增温处理6 a,木荷和红枝蒲桃的N浓度都呈增加趋势,红锥和马尾松的下降;除马尾松的P浓度下降外,其他3种植物都上升。
从图5可见,增温处理下,植物的总N积累量发生显著变化(<0.05),增温处理2、3和6 a, 木荷的N积累量分别比对照显著增加了62.1%、84.4%和100.3%;马尾松分别显著增加了342.8%、324%和185.7% (<0.05)。然而,增温处理6 a的红锥和红枝蒲桃总N积累量分别下降了36.8%和6.2%。
从图5可见,与对照相比,增温处理2、3和6 a使木荷P积累量分别显著增加了74.3%、140.9%和142.3%,马尾松分别显著增加了409.7%、401.9%和201.4% (<0.05)。然而,在增温处理6 a的红锥和红枝蒲桃P积累量分别减少了5.6%和3.3%。
4 结论和讨论
4.1 增温对植物生物量及其分配模式的影响
本研究结果表明,增温处理2~3 a提高了4种植物的总生物量,6 a长期增温使木荷和马尾松的总生物量持续显著增加,而红枝蒲桃与红锥的却呈下降的趋势,其中红锥的生长受到一定程度抑制, 基径和树高均下降,这是因为增温初期提高了植物的光合能力和水分运输能力,同时增温导致土壤矿化速率加快,促进了植物对土壤养分的吸收,进而加速了植物的生长和总生物量的积累[39–43],这与Li等[44]的研究结果一致。马尾松是一种原生裸子植物,在森林早期演替阶段具有比其他共存种更高的竞争能力和生长速率[45]。增温6 a出现种间差异可能与不同植物对当地土壤水分条件的适应性差异有关[46]。Bowman[47]的研究表明,增温会降低水分利用率从而抑制植物生长。也有研究表明,长期增温可能会造成热带植物水分胁迫,进而导致地上生物量和碳储量减少[48]。长期增温使红枝蒲桃和红锥的总生物量下降,可能是其相对较低的生长速率和竞争资源的能力,为应对气候变化而调整自身生长策略。
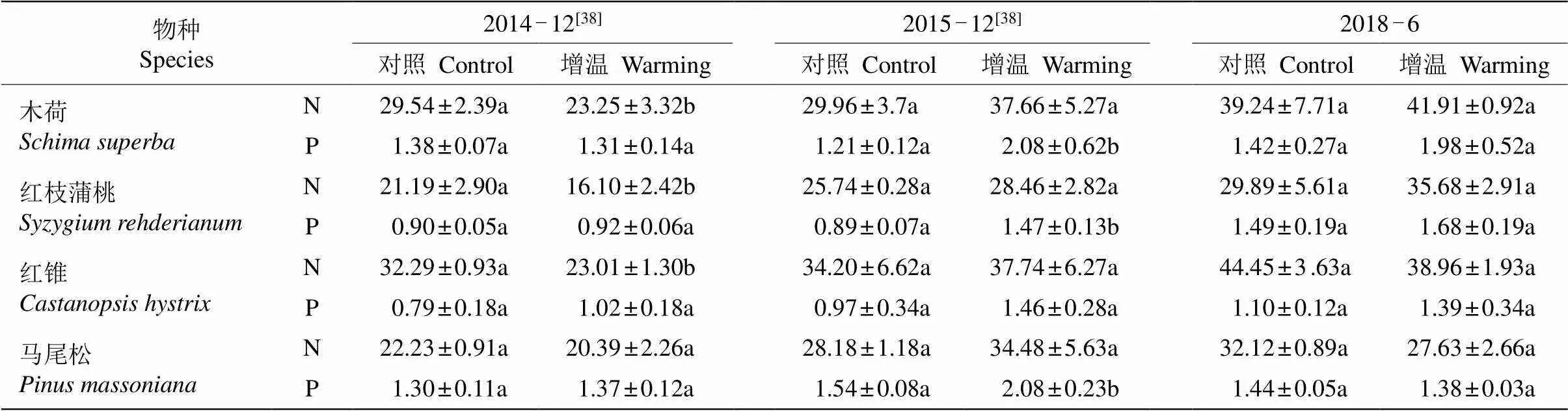
表4 增温对4种植物养分浓度的影响(n=3)
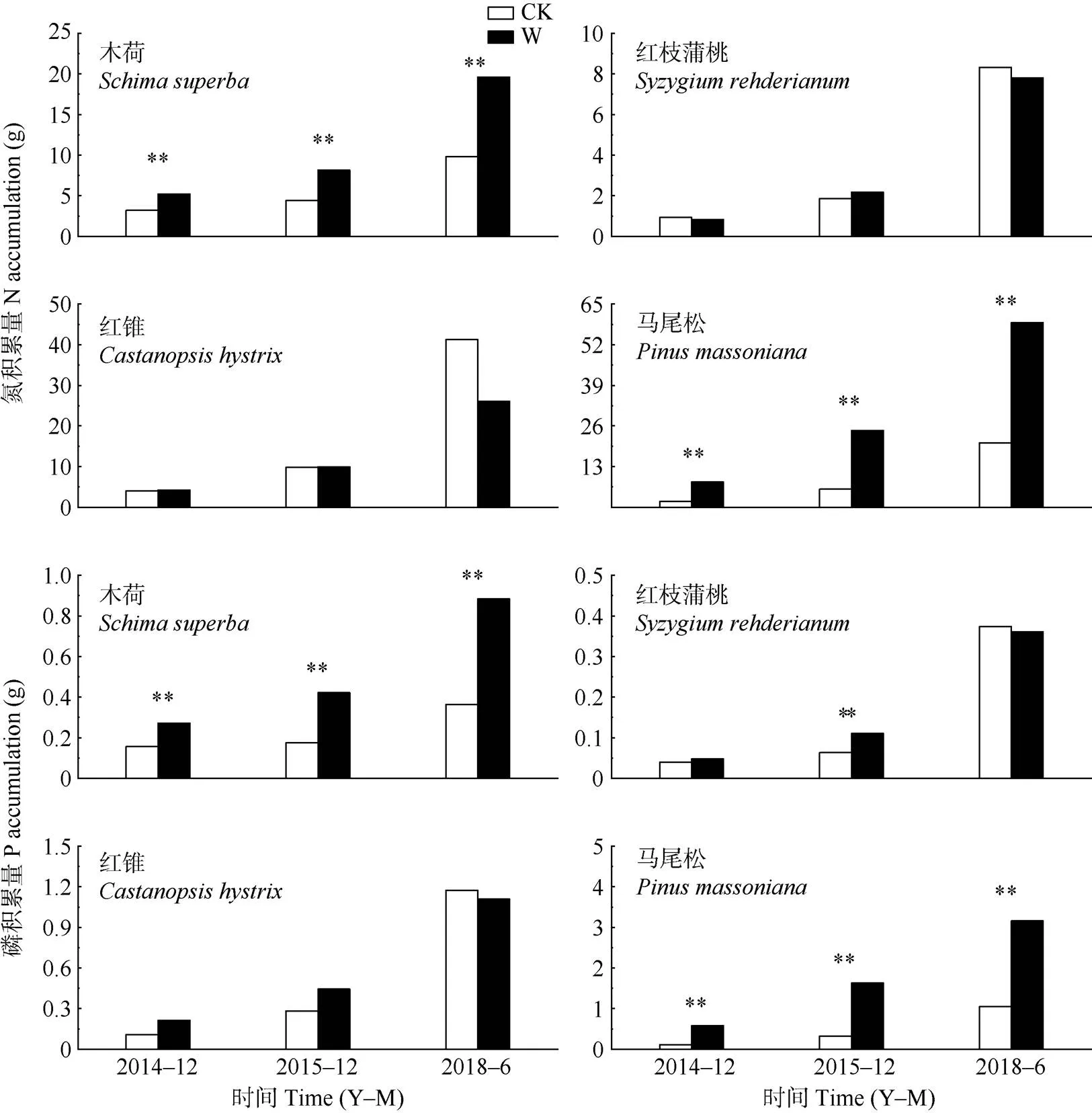
图5 增温对4种树种养分积累量的影响
本研究结果表明,短期增温能增加4种植物的茎质比,但叶质比、根质比和根冠比下降。6 a长期增温使木荷和马尾松的茎质比显著增加,叶质比、根质比和根冠比显著下降,红枝蒲桃和红锥的根质比和根冠比有所下降。在短期增温下,4种植物均以降低根和叶的生物量为代价,分配更多的生物量到茎,以木荷和马尾松表现显著。Lin等[49]分析了国内5种不同森林类型的生物量数据,认为增温显著提高了亚热带马尾松林中茎生物量的分配比例,这与本试验结果一致,叶旺敏等[50]的研究结果也支持这一观点。由于植物在增温前期快速生长,所以植物面临在有限的空间内获取更多光源和养分的激烈竞争,已有研究证实,植物为了获取更充足的光源,通常会提高茎的生物量以便于向外扩展生长空间[51]。6 a增温使生物量分配模式出现种间差异,可能是由于长期增温导致的土壤含水率下降,出现了土壤水分胁迫,其中木荷和马尾松能更好地适应这种水分条件的变化,如基径和树高持续显著增加,因此能够在样方中占据优势地位。红枝蒲桃和红锥则需要通过调整自身的生物量分配来维持其正常生长,而红锥可能对土壤水分的变化更敏感,所以将更多的生物量分配到根以寻找水资源[52–53]。这与植物最佳分配理论相吻合,即植物为了探索有限的环境资源,将在某种器官上投入更多比例[54]。红锥增加对根的分配可以提高对水和养分的吸收,但是这种生物量分配策略上的变化同时也导致其生长欠佳(如树高和基径的降低)。另外, 红锥具有根系较发达等特性[55–56],这可能也是红锥根质比和根冠比上升的原因之一。
4.2 增温对植物养分积累的影响
短期增温增加了4种植物的N和P积累量,这与其生物量增加和土壤养分供应有关。温度升高可以促进凋落物分解,使更多的养分释放出来供植物生长利用[57]。前期研究结果表明,增温增加了南亚热带森林土壤中有效P的供应和植物各器官的P浓度,但减少了南亚热带森林土壤中的N供应,4种植物各器官的N浓度也显著下降[37,58]。短期增温促进了植物生物量累积,进而促进了植物P积累量的增加。而增温条件下植物体内N积累量的变化主要取决于其生物量的变化,这与郭建平等[59]的研究结果一致。同时,增温条件下,南亚热带地区土壤中的N供应在一定程度上并不是限制加速植物生长的主要因素[60]。
6 a的长期增温增加了木荷和马尾松的N和P的积累量,而红锥与红枝蒲桃的N和P的积累量却呈下降趋势,主要原因可能是红锥与红枝蒲桃在长期增温下生物量呈减少的趋势,这与长期增温造成红锥和红枝蒲桃根系吸收能力减弱,从而导致植物缺乏养分或者水分有关。前期研究结果表明,增温加速了土壤蒸发和植物叶片蒸腾,植物可能通过土壤-植物-大气连续性地流失水分[61]。随着增温时间的延长,其蒸发需求也增加,进一步加重了森林生态系统的有效干旱[62–63]。张浩玮等[64]的研究表明水分胁迫显著降低了野生草地早熟禾整株及地上部分的N和P积累量。这也从一定程度上证明了水分胁迫会抑制植物N和P积累量,进而迫使植物通过改变N和P的分配与利用方式,以应对水分供应不足的环境做出适应性反映。4种植物的养分积累变化趋势表明,在全球变暖背景下,鼎湖山混交林中的红枝蒲桃和红锥对气候变化的适应幅度和竞争力要低于木荷和马尾松。王从容等[65]的研究也表明,在我国亚热带森林中,马尾松和木荷具有更强的生态适应性和竞争力。由此可见,长期增温对植物生长的影响是存在种间差异性的[66],而这种差异可能会进一步影响我国南亚热带混交林群落的物种组成。
增温对我国鼎湖山混交林中4种优势树种的生物量分配和养分积累都有显著的影响,但主要取决于植物种类。在6 a增温过程中,木荷和马尾松的生长、总生物量、N和P的积累量显著增加,茎质比显著增加,叶质比、根质比和根冠比显著下降, 这是由于木荷和马尾松将生物量更多投入到茎上供植物地上部分快速生长;而长期增温使红枝蒲桃和红锥总生物量、N和P的积累量相对减少,生物量更多投入到地下部分,根冠比有所上升,这是因为红枝蒲桃和红锥对土壤水分的适应范围更为狭窄, 竞争养分的能力较弱,因此需要调整其生物量分配策略来获取自身所需要的养分和水分。Schipper等[67]报道,温度升高对热带雨林树木的茎生物量产生显著的负面影响。而Gennaretti等[68]的研究表明,增温显著提高了北方森林植物的茎生物量。这种不同的生物量分配策略可以重新建立汇-库关系,并且可能对当地环境适应幅度广泛的树种更有益。由此可见,长期增温下,木荷和马尾松可能相比于红枝蒲桃和红锥更具有生长优势。在未来全球变暖的大背景下,植物的生物量分配模式和养分积累存在种间差异性,这种差异可能会给南亚热带混交林未来的群落物种组成带来潜在的影响。
[1] IPCC. Climate Change 2018: Special Report on Global Warming of 1.5℃ [M]. Cambridge: Cambridge University Press, 2018: 32.
[2] ZHAO X Q, HUANG J, LU J, et al. Study on the influence of soil micro- bial community on the long-term heavy metal pollution of different land use types and depth layers in mine [J]. Ecotoxicol Environ Saf, 2019, 170: 218–226. doi: 10.1016/j.ecoenv.2018.11.136.
[3] ZHAO J X, LUO T X, WEI H X, et al. Increased precipitation offsets the negative effect of warming on plant biomass and ecosystem respi- ration in a Tibetan alpine steppe [J]. Agric For Meteorol, 2019, 279: 107761. doi: 10.1016/j.agrformet.2019.107761.
[4] CHEN F S, Niklas K J, Liu Y, et al. Nitrogen and phosphorus addi- tions alter nutrient dynamics but not resorption efficiencies of Chinese fir leaves and twigs differing in age [J]. Tree Physiol, 2015, 35(10): 1106–1117. doi: 10.1093/treephys/tpv076.
[5] Pan Y D, Birdsey R A, Fang J Y, et al. A large and persistent carbon sink in the world’s forests [J]. Science, 2011, 333(6045): 988– 993. doi: 10.1126/science.1201609.
[6] Cleveland C C, Townsend A R, Taylor P, et al. Relation- ships among net primary productivity, nutrients and climate in tropical rain forest: A pan-tropical analysis [J]. Ecol Lett, 2011, 14(9): 939–947. doi: 10.1111/j.1461-0248.2011.01658.x.
[7] Dusenge M E, Way D A. Warming puts the squeeze on photo- synthesis-lessons from tropical trees [J]. J Exp Bot, 2017, 68(9): 2073– 2077. doi: 10.1093/jxb/erx114.
[8] Mau A C, Reed S C, Wood T E, et al. Temperate and tropical forest canopies are already functioning beyond their thermal thresholds for photosynthesis [J]. Forests, 2018, 9(1): 47. doi: 10.3390/f9010047.
[9] Allen C D, Breshears D D, McDowell N G. On undere- stimation of global vulnerability to tree mortality and forest die-off from hotter drought in the Anthropocene [J]. Ecosphere, 2015, 6(8): 1– 55. doi: 10.1890/ES15-00203.1.
[10] Duan H L, Huang G M, Zhou S X, et al. Dry mass production, allocation patterns and water use efficiency of two conifers with different water use strategies under elevated [CO2], warming and drought con- ditions [J]. Eur J For Res, 2018, 137(5): 605–618. doi: 10.1007/s103 42-018-1128-x.
[11] Cavaleri M A, Reed S C, Smith W K, et al. Urgent need for warming experiments in tropical forests [J]. Glob Change Biol, 2015, 21(6): 2111–2121. doi: 10.1111/gcb.12860.
[12] Wu T, Liu S Z, Lie Z Y, et al. Divergent effects of a 6-year warming experiment on the nutrient productivities of subtropical tree species [J]. For Ecol Manag, 2020, 461: 117952. doi: 10.1016/j.foreco.2020.117952.
[13] Qi Y L, Wei W, Chen C G, et al. Plant root-shoot biomass allocation over diverse biomes: A global synthesis [J]. Glob Ecol Conserv, 2019, 18: e00606. doi: 10.1016/j.gecco.2019.e00606.
[14] Kasurinen A, Koikkalainen K, Anttonen M J, et al. Root morphology, mycorrhizal roots and extramatrical mycelium growth in silver birch (Roth) genotypes exposed to experimental warming and soil moisture manipulations [J]. Plant Soil, 2016, 407: 341–353. doi: 10.1007/s11104-016-2891-4.
[15] Wang P, Heijmans M M P D, Mommer L J, et al. Belowground plant biomass allocation in tundra ecosystems and its relationship with temperature [J]. Environ Res Lett, 2016, 11: 055003. doi: 10.1088/ 1748-9326/11/5/055003.
[16] Yu L, Song M Y, Xia Z C, et al. Elevated temperature differently affects growth, photosynthetic capacity, nutrient absorption and leaf ultrastructure ofandunder intra- and interspecific competition [J]. Tree Physiol, 2019, 39(8): 1342–1357. doi: 10.1093/treephys/tpz044.
[17] Rustad L E, Campbell J L, Marion G M, et al. A meta-analysis of the response of soil respiration, net nitrogen mineralization, and aboveground plant growth to experimental ecosystem warming [J]. Oecologia, 2001, 126: 543–562. doi: 10.1007/s004420000544.
[18] AKAJI Y, Inoue T, Tomimatsu H, et al. Photosynthesis, respi- ration, and growth patterns ofseedlings in relation to growth temperature [J]. Trees, 2019, 33: 1041–1049. doi: 10.1007/ s00468-019-01840-7.
[19] Wang L, Niu K C, Yang Y H, et al. Patterns of above- and belowground biomass allocation in China’s grasslands: Evidence from individual-level observations [J]. Sci China Life Sci, 2010, 53(7): 851– 857. doi: 10.1007/s11427-010-4027-z.
王亮, 牛克昌, 杨元合, 等. 中国草地生物量地上-地下分配格局: 基于个体水平的研究[J]. 中国科学: 生命科学, 2010, 40(7): 642–649.
[20] Ruiz-Vera U M, Siebers M, Gray S B, et al. Global warming can negate the expected CO2stimulation in photosynthesis and produc- tivity for soybean grown in the midwestern United States [J]. Plant Biol, 2013, 162: 410–423. doi: 10.1104/pp.112.211938.
[21] Li X J, Liu X F, Lin C F, et al. Effects of experimental soil warming on plant biomass allocation during the early stages of succession in a subtropical forest in China [J]. Acta Ecol Sin, 2017, 37(1): 25–34. doi: 10.5846/stxb201607261529.
李晓杰, 刘小飞, 林成芳, 等. 土壤增温调节中亚热带森林更新初期植物生物量分配格局 [J]. 生态学报, 2017, 37(1): 25–34. doi: 10. 5846/stxb201607261529.
[22] Zeng Z, Huan H H, Liu G, et al. Effects of elevated temperature and CO2concentration on growth and leaf quality ofseedlings [J]. Chin J Appl Ecol, 2016, 27(8): 2445–2451. doi: 10.13287/ j.1001-9332.201608.022.
曾贞, 郇慧慧, 刘刚, 等. 增温和升高CO2浓度对桑树幼苗的生长和叶片品质的影响 [J]. 应用生态学报, 2016, 27(8): 2445–2451. doi: 10.13287/j.1001-9332.201608.022.
[23] Zhang J W, Dong S T, Wang K J, et al. Effects of increasing field temperature on photosynthetic characteristics of summer maize [J]. Chin J Appl Ecol, 2008, 19(1): 81–86.
张吉旺, 董树亭, 王空军, 等. 大田增温对夏玉米光合特性的影响 [J]. 应用生态学报, 2008, 19(1): 81–86.
[24] Xu Z F, Hu T X, Zhang L, et al. Short-term gas exchange responses ofto simulated global warming in a timber-line ecotone, eastern Tibetan Plateau, China [J]. Chin J Plant Ecol, 2010, 34(3): 263– 270. doi: 10.3773/j.issn.1005-264x.2010.03.003.
徐振锋, 胡庭兴, 张力, 等. 青藏高原东缘林线交错带糙皮桦幼苗光合特性对模拟增温的短期响应 [J]. 植物生态学报, 2010, 34(3): 263–270. doi: 10.3773/j.issn.1005-264x.2010.03.003.
[25] Xu M H, Liu M, Zhai D T, et al. Dynamic changes in biomass and its relationship with environmental factors in an alpine meadow on the Qinghai-Tibetan Plateau, based on simulated warming experiments [J]. Acta Ecol Sin, 2016, 36(18): 5759–5767. doi: 10.5846/stxb20150417 0794.
徐满厚, 刘敏, 翟大彤, 等. 青藏高原高寒草甸生物量动态变化及与环境因子的关系——基于模拟增温实验 [J]. 生态学报, 2016, 36(18): 5759–5767. doi: 10.5846/stxb201504170794.
[26] Kudo G, Suzuki S. Warming effects on growth, production, and vegetation structure of alpine shrubs: A five-year experiment in northern Japan [J]. Oecologia, 2003, 135: 280–287. doi: 10.1007/s00442-003- 1179-6.
[27] Li J R, Liu Z H. High-cold meadow plants respond to long-term warming [J]. Qinghai Pratacult, 2017, 26(3): 13–18,24. doi: 10.3969/j. issn.1008-1445.2017.03.003.
李京蓉, 刘泽华. 高寒草甸植物对长期增温的响应 [J]. 青海草业, 2017, 26(3): 13–18,24. doi: 10.3969/j.issn.1008-1445.2017.03.003.
[28] de Boer H C, Deru J G C, Hoekstra N J, et al. Strategic timing of nitrogen fertilization to increase root biomass and nitrogen-use efficiency ofL. [J]. Plant Soil, 2016, 407: 81–90. doi: 10.1007/s11104-016-2917-y.
[29] Rogers C W, Dari B, HU G S, et al. Dry matter production, nutrient accumulation, and nutrient partitioning of barley [J]. J Plant Nutri Soil Sci, 2019, 182(3): 367–373. doi: 10.1002/jpln.201800336.
[30] Kuster T M, Schleppi P, Hu B, et al. Nitrogen dynamics in oak model ecosystems subjected to air warming and drought on two different soils [J]. Plant Biol, 2013, 15(S1): 220–229. doi: 10.1111/j. 1438-8677.2012.00686.x.
[31] Zhao S C, Xu X P, Wei D, et al. Soybean yield, nutrient uptake and stoichiometry under different climate regions of northeast China [J]. Sci Rep, 2020, 10(1): 8431. doi: 10.1038/s41598-020-65447-6.
[32] NISHITANI S, ISHIDA A, NAKAMURA T, et al. Functional differ- rences in seasonally absorbed nitrogen in a winter-green perennial herb [J]. Roy Soc Open Sci, 2020, 7(1): 190034. doi: 10.1098/rsos.190034.
[33] Dawes M A, Schleppi P, Hättenschwiler S, et al. Soil warming opens the nitrogen cycle at the alpine treeline [J]. Glob Change Biol, 2017, 23(1): 421–434. doi: 10.1111/gcb.13365.
[34] Hou Y, Wang K Y, Zhang C. Effects of elevated CO2concen- tration and temperature on nutrient accumulation and allocation inseedlings [J]. Chin J Appl Ecol, 2008, 19(1): 13–19.
侯颖, 王开运, 张超. 大气二氧化碳浓度与温度升高对红桦幼苗养分积累和分配的影响 [J]. 应用生态学报, 2008, 19(1): 13–19.
[35] Wang J Q, Li L Q, LAM S K, et al. Changes in nutrient uptake and utilization by rice under simulated climate change conditions: A 2-year experiment in a paddy field [J]. Agric For Meteorol, 2018, 250–251: 202–208. doi: 10.1016/j.agrformet.2017.12.254.
[36] WANG J, YU B Y, HUANG J G. Xylem formation and response to climate ofin Dinghushan Mountain [J]. J Trop Subtrop Bot, 2020, 28(5): 445–454. doi: 10.11926/jtsb.4204.
王婕, 余碧云, 黄建国. 鼎湖山锥栗木质部形成及其对气候的响应 [J]. 热带亚热带植物学报, 2020, 28(5): 445–454. doi: 10.11926/jtsb. 4204.
[37] Liu J X, Li Y L, Liu S Z, et al. An introduction to an experimental design for studying effects of air temperature rise on model forest ecosystems [J]. Chin J Plant Ecol, 2013, 37(6): 558–565. doi: 10.3724/ SP.J.1258.2013.00057.
刘菊秀, 李跃林, 刘世忠, 等. 气温上升对模拟森林生态系统影响实验的介绍 [J]. 植物生态学报, 2013, 37(6): 558–565. doi: 10.3724/ SP.J.1258.2013.00057.
[38] Li X, Lie Z Y, Wu T, et al. Effect of warming on nutrients concen- trations and stoichiometry of 4 tree species in south subtropical mixed forest [J]. Ecol Environ Sci, 2019, 28(5): 890–897. doi: 10.16258/j. cnki.1674-5906.2019.05.005.
李旭, 列志旸, 吴婷, 等. 增温对南亚热带混交林4个树种养分含量及化学计量的影响 [J]. 生态环境学报, 2019, 28(5): 890–897. doi: 10.16258/j.cnki.1674-5906.2019.05.005.
[39] Bremner J M, Mulvaney C S. Nitrogen-total [M]// PAGE A L, MILLER R H, KEENEY D R. Methods of Soil Analysis: Part 2. Chemical and Microbiological Properties: Agronomy Monograph No. 9. 2nd ed. Madison, Wisconsin: American Society of Agronomy, 1982: 595–624.
[40] Li Y Y, Liu J X, Zhou G Y, et al. Warming effects on photosynthesis of subtropical tree species: A translocation experiment along an alti- tudinal gradient [J]. Sci Rep, 2016, 6: 24895. doi: 10.1038/srep24895.
[41] Drake J E, Tjoelker M G, Varhammar A, et al. Trees tolerate an extreme heatwavesustained transpirational cooling and increased leaf thermal tolerance [J]. Glob Change Biol, 2018, 24(6): 2390–2402. doi: 10.1111/gcb.14037.
[42] Butler S M, Melillo J M, Johnson J E, et al. Soil warming alters nitrogen cycling in a New England forest: Implications for eco- system function and structure [J]. Oecologia, 2012, 168(3): 819–828. doi: 10.1007/s00442-011-2133-7.
[43] Liu J X, Liu S E, Li Y Y. Warming effects on the decomposition of two litter species in model subtropical forests [J]. Plant Soil, 2017, 420 (1/2): 277–287. doi: 10.1007/s11104-017-3392-9.
[44] Li Y Y, Zhou G Y, Liu J X. Different growth and physiological responses of six subtropical tree species to warming [J]. Front Plant Sci, 2017, 8: 1511. doi: 10.3389/fpls.2017.01511.
[45] Tang X L, Wang Y P, Zhou G Y, et al. Different patterns of ecosystem carbon accumulation between a young and an old-growth subtropical forest in southern China [J]. Plant Ecol, 2011, 212(8): 1385– 1395. doi: 10.1007/s11258-011-9914-2.
[46] Crous K Y, Quentin A G, Lin Y S, et al. Photosynthesis of temperatetrees outside their native range has limited adjustment to elevated CO2and climate warming [J]. Glob Change Biol, 2013, 19(12): 3790–3807. doi: 10.1111/gcb.12314.
[47] Bowman D M J S, Williamson G J, Keenan R J, et al. A warmer world will reduce tree growth in evergreen broadleaf forests: Evidence from Australian temperate and subtropical eucalypt forests [J]. Glob Ecol Biogeogr, 2014, 23(8): 925–934. doi: 10.1111/geb.12171.
[48] Sullivan M J P, Lewis S L, Affum-Baffoe K, et al. Long-term thermal sensitivity of Earth’s tropical forests [J]. Science, 2020, 368 (6493): 869-874. doi: 10.1126/science.aaw7578.
[49] LIN W Q, FANG Y R, XUE L. Forest biomass allocation vary with temperature in five forest types of China [J]. Int J Agric Biol, 2019, 21: 1043–1048.
[50] YE W M, XIONG D C, YAng Z J, et al. Effect of soil warming on growth and photosynthetic characteristics ofsaplings [J]. Acta Ecol Sin, 2019, 39(7): 2501–2509. doi: 10.5846/stxb 201801150110.
叶旺敏, 熊德成, 杨智杰, 等. 模拟增温对杉木幼树生长和光合特性的影响 [J]. 生态学报, 2019, 39(7): 2501–2509. doi: 10.5846/stxb 201801150110.
[51] Wang G G, Bauerle W L, Mudder B T. Effects of light acclimation on the photosynthesis, growth, and biomass allocation in American chestnut () seedlings [J]. For Ecol Manag, 2006, 226(1–3): 173–180. doi: 10.1016/j.foreco.2005.12.063.
[52] Wan S Q, Hui D F, Wallace L, et al. Direct and indirect effects of experimental warming on ecosystem carbon processes in a tallgrass prairie [J]. Glob Biogeochem Cycles, 2005, 19(2): GB2014. doi: 10. 1029/2004GB002315.
[53] Niu S L, Wu M Y, Han Y, et al. Water-mediated responses of ecosystem carbon fluxes to climatic change in a temperate steppe [J]. New Phytol, 2008, 177(1): 209–219. doi: 10.1111/j.1469-8137.2007. 02237.x.
[54] Poorter H, Niklas K J, Reich P B, et al. Biomass allocation to leaves, stems and roots: Meta-analyses of interspecific variation and environmental control [J]. New Phytol, 2012, 193(1): 30–50. doi: 10. 1111/j.1469-8137.2011.03952.x.
[55] Ming A G, Liu S R, Mo H H, et al. Comparison of carbon storage in pure and mixed stands ofandin subtropical China [J]. Acta Ecol Sin, 2016, 36(1): 244– 251. doi: 10.5846/stxb201405211041.
明安刚, 刘世荣, 莫慧华, 等. 南亚热带红锥、杉木纯林与混交林碳贮量比较 [J]. 生态学报, 2016, 36(1): 244–251. doi: 10.5846/stxb 201405211041.
[56] Wu M, Deng P, Zhao Y, et al. Vertical distribution and physio- logical senescence characteristics of fine roots inMiq. plantations at different ages [J]. Chin J Ecol, 2019, 38(9): 2622– 2629. doi: 10.13292/j.1000-4890.201909.023.
吴敏, 邓平, 赵英, 等. 不同林龄红锥人工林细根垂直分布和衰老生理特征 [J]. 生态学杂志, 2019, 38(9): 2622–2629. doi: 10.13292/j. 1000-4890.201909.023.
[57] Litton C M, Giardina C P, Freeman K R, et al. Impact of mean annual temperature on nutrient availability in a tropical montane wet forest [J]. Front Plant Sci, 2020, 11: 784. doi: 10.3389/fpls.2020. 00784.
[58] Lie Z Y, Lin W, Huang W J, et al. Warming changes soil N and P supplies in model tropical forests [J]. Biol Fert Soils, 2019, 55(7): 751–763. doi: 10.1007/s00374-019-01382-7.
[59] Guo J P, Gao S H. Impacts of CO2enrichment and soil drought on C, N accumulation and distribution in[J]. J Soil Water Conserv, 2005, 19(2): 118–121. doi: 10.3321/j.issn:1009-2242.2005.02. 031.
郭建平, 高素华. 高CO2浓度和土壤干旱对贝加尔针茅C, N积累和分配的影响 [J]. 水土保持学报, 2005, 19(2): 118–121. doi: 10.3321/j. issn:1009-2242.2005.02.031.
[60] Liu J X, Li Y Y, Xu Y, et al. Phosphorus uptake in four tree species under nitrogen addition in subtropical China [J]. Environ Sci Pollut Res Int, 2017, 24(24): 20005–20014. doi: 10.1007/s11356-017-9633-x.
[61] Wu G L, Liu H, Hua L, et al. Differential responses of stomata and photosynthesis to elevated temperature in two co-occurring subtropical forest tree species [J]. Front Plant Sci, 2018, 9: 467. doi: 10.3389/fpls. 2018.00467.
[62] Restaino C M, Peterson D L, Littell J. Increased water deficit decreases douglas fir growth throughout western US forests [J]. Proc Natl Acad Sci USA, 2016, 113(34): 9557–9562. doi: 10.1073/pnas. 1602384113.
[63] Zhang X L, Manzanedo R D, D’Orangeville L, et al. Snowmelt and early to mid-growing season water availability augment tree growth during rapid warming in southern Asian boreal forests [J]. Glob Change Biol, 2019, 25(10): 3462–3471. doi: 10.1111/gcb.14749.
[64] Zhang H W, Bai X M, Fan J H, et al. Effect of water treatments on biomass, accumulation and allocation of nitrogen and phosphorus of[J]. Grassland Turf, 2018, 38(5): 8–15. doi: 10.3969/ j.issn.1009-5500.2018.05.002.
张浩玮, 白小明, 樊敬辉, 等. 不同水分处理对草地早熟禾生物量及N、P积累与分配的影响 [J]. 草原与草坪, 2018, 38(5): 8–15. doi: 10.3969/j.issn.1009-5500.2018.05.002.
[65] Wang C R, Li S Z, Yang X Y. Competitive patterns of pioneer species at different restoration levels in the subtropical red soil erosion and degradation region [J]. Chin J Appl Environ Biol, 2019, 25(2): 239–245. doi: 10.19675/j.cnki.1006-687x.2018.05044.
王从容, 李守中, 杨贤宇. 亚热带红壤侵蚀退化区不同恢复水平生境内先锋树种竞争特征 [J]. 应用与环境生物学报, 2019, 25(2): 239–245. doi: 10.19675/j.cnki.1006-687x.2018.05044.
[66] Cao J, Liu H Y, Zhao B, et al. Species-specific and elevation- differentiated responses of tree growth to rapid warming in a mixed forest lead to a continuous growth enhancement in semi-humid North- east Asia [J]. For Ecol Manag, 2019, 448: 76–84. doi: 10.1016/j.foreco. 2019.05.065.
[67] SCHIPPERS P, STERCK F, VLAM M, et al. Tree growth variation in the tropical forest: Understanding effects of temperature, rainfall and CO2[J]. Glob Change Biol, 2015, 21(7): 2749–2761. doi: 10.1111/gcb.12877.
[68] GENNARETTI F, GEA-IZQUIERDO G, BOUCHER E, et al. Ecophy- siological modeling of photosynthesis and carbon allocation to the tree stem in the boreal forest [J]. Biogeosciences, 2017, 14(21): 4851–4866. doi: 10.5194/bg-14-4851-2017.
Effects of Warming on Biomass Allocation Patterns and Nutrient Accumulations of Four Dominant Tree Species in Mixed Forest of Dinghushan, China
TAN Na-dan1,2, LI Xu1, WU Ting1, LIE Zhi-yang1, LIU Xu-jun1, LIU Shi-zhong1, CHEN Ping2, LIU Ju-xiu1*
(1. Center for Plant Ecology, Core Botanical Garden, Chinese Academy of Sciences, Guangzhou 510650, China; 2. Zhongkai University of Agriculture and Engineering,Guangzhou 510225, China)
In order to understand the effects of future global warming on species composition of forest ecosystems in the south subtropical region, the biomass allocation patterns and nutrient accumulations of four dominant tree species, such as,,and, were studied under simulated warming by translocating model forest from high altitude to low altitude in Dinghushan, Guangdong. The results showed that warming increases the average atmospheric temperature by (1.28±0.60)℃, and the average soil temperature by (1.04±0.30)℃. Forandunder warming for 6 years, the height significantly increased by 83.0% and 52.1%, and basal diameter by 37.1% and 76.9%, respectively, and the stem mass ratio significantly increased, but the ratios of root mass, leaf biomass and root to shoot significantly decreased, as well as nutrient accumulations increased significantly by 100.3% and 185.7%, respectively. However, warming had no significant effect on the height, basal diameter, biomass allocation patterns, and nutrient accumulations ofand. Therefore, the effects of 6-year warming on biomass allocation pattern and nutrient accumulation were different among four dominant tree species.andmight have strong adaptability under long-term warming due to their high biomass and nutrient accumulation. These differences might have a potential impact on community structure and function in subtropical mixed forests in China.
Warming; Biomass allocation pattern; Nutrient accumulations; Dinghushan
10.11926/jtsb.4325
2020–10–22
2021–01–05
广东省重点领域研发计划项目(2020B1111530004); 国家自然科学基金项目(41977287, 41991285); 广州市民生科技攻关项目(201903010021)资助
This work was supported by the Project for Key Research and Development in Guangdong (Grant No. 2020B1111530004), the National Natural Science Foundation of China (Grant No. 41977287, 41991285), and the Project for Livelihood Science and Technology in Guangzhou City (Grant No. 201903010021).
谭钠丹,硕士研究生,主要从事生态系统生态学研究。E-mail: tannd@scbg.ac.cn
E-mail: ljxiu@scbg.ac.cn
猜你喜欢
杂志排行

热带亚热带植物学报的其它文章
- 丛枝菌丝网络介导苜蓿植株间机械损伤信号的传递
- 粉叶小檗(小檗科)的形态变异式样及分类修订
- Pholidota advena (Par. & Rchb. f.) Hook. f., A Newly Recorded Species of Orchidaceae from China
- Agapetes nana, A Newly Recorded Species of Ericaceae from China
- Michelia taishanensis Y. H. Tong, X. E. Ye, X. H. Ye & Yu Q. Chen (Magnoliaceae), A New Species from Guangdong
- 基于热消散探针法的植物水分运输的研究进展