处理松材线虫病疫木的木腐真菌筛选*
2021-08-09王旻嘉叶建仁涂煜昇杜方平
王旻嘉 叶建仁 涂煜昇 杜方平
(1.南京林业大学林学院 南方现代林业协同创新中心 南京 210037;2.安徽金寨县金山寨食(药)用菌种植专业合作社 金寨 237300)
松材线虫病(Bursaphelenchusxylophilus)又称松树萎蔫病(pine wilt disease,PWD),是一种短时间内就可导致松属(Pinus)树种大量萎蔫死亡的毁灭性病害,全球已有52个国家将其作为检疫性病虫害(Motaetal.,2009)。该病害起源于北美,目前分布于美国、加拿大、墨西哥、日本、韩国、中国、葡萄牙和西班牙等(Inacioetal.,2015;王曦茁等,2018),对北美洲本土松种未造成明显危害,对亚洲国家松树危害最为严重(Alexanderetal.,2011)。1982年中国在江苏南京首次发现松材线虫病(孙永春,1982),其后病害不断扩散蔓延。据国家林业和草原局2020年第4号公告,我国松材线虫病疫区已有18个省(区、市)、666个县级行政区、4 333个乡镇级行政区,1年内新增85个县级疫区,发生面积已超过110万hm2(董瀛谦等,2020;潘佳亮等,2021),中国已成为全球松材线虫病发生最严重的地区。随着疫情的发展,病害寄主植物、传播媒介昆虫和适生范围都在不断变化,病情呈现由点状分布向片状发展,逐渐向西向北呈现跳跃式扩散趋势(程功等,2015;李晓冬等,2020)。2017年,疫情发生区域已突破年均10 ℃等温线(理永霞等,2018),红松(Pinuskoraiensis)、落叶松(Larixspp.)成为病害新寄主,云杉花墨天牛(Monochamussaltuarius)成为松材线虫病在我国北方森林的传播新媒介(于海英等,2018;2019)。2019年,首次发现樟子松(P.sylvestrisvar.mongolica)自然感病(于海英等,2020)。全国6 000万hm2松林正面临着松材线虫病严重威胁,防控形势十分严峻。
人为因素造成疫木及其制品的扩散流通是导致我国疫情在短短四十年内蔓延爆发的主要原因(潘宏阳,2007;叶建仁等,2012)。针对这一现状,目前采取的主要防控措施包括检疫与监测、疫木除治和媒介昆虫防治等,其中疫木除治是从源头阻断该病害传播蔓延的关键环节(叶建仁,2019)。目前的疫木处理方法是就地粉碎或焚烧等(张丽荣等,2019;朱宁波,2019),但依然还是存在着疫木就地除治比较困难、不彻底,造成环境污染等问题(郑礼平,2017),故亟待寻找更加高效环保的疫木除害新技术,从源头上提高阻断病害扩散蔓延的效率。
疫木是由木材、病原线虫、媒介昆虫、真菌、细菌等构成的复杂的生物群落,木腐真菌作为群落中的分解者,具备在病死木上大量繁殖并通过分解木材实现物质循环的能力。已有研究表明,一些木腐真菌对松材线虫的生长和繁殖有较强的抑杀作用 (董锦艳等,2000;李玉中等,2020;徐红梅等,2018;张建平等,2003),但当前关于木腐菌作用于松材线虫的研究大多处于室内试验阶段,对松材线虫病疫木的处理相关研究多针对疫木伐桩(陈瑶等,2008a)以及茯苓的疫木种植(吴云忠,2013),直接针对伐倒木段除害的研究很少。前人利用菌袋培养菌种,培育耗时长且培养基成分要求高,成本昂贵,不适用于直接广泛推广。因此,寻找更多容易扩繁、定殖能力强、除害效果佳的木腐真菌,使其作用于疫木,改变病原线虫的适生环境,减少疫木中松材线虫数量,是实现疫木不下山,就地除治的可能突破口。本研究通过筛选具备处理松材线虫病疫木潜力的木腐真菌,探究不同菌株的田间效果,为研发松材线虫病疫木除治的新途径提供新思路。
1 材料与方法
1.1 试验地概况
试验地位于江苏省镇江市句容林场摇令口工区(119°12′E,32°05′N),地处长江南岸,江苏省西南部,属亚热带季风气候,四季分明,温暖湿润,雨量充沛,相对湿度76%,无霜期237天。年平均温度15.4 ℃,年极端气温最高39.7 ℃,最低-13.1 ℃,年平均降水量1 106 mm。试验所用疫木均为林区内15年生当年枯死的松材线虫病黑松(Pinusthunbergii)疫木。
1.2 供试材料
2019年4月从安徽省池州市九华山、江苏省南京市中山陵、江苏省镇江市下蜀林场采集腐朽松木样品,从中分离纯化得到147个真菌菌株(编号以J、H、S开头的菌株为分离自九华山样品,编号以Z开头的菌株为分离自中山陵样品,编号以X、W开头的菌株为分离自下蜀林场样品),保存于4 ℃冰箱备用。糙皮侧耳(Pleurotusostreatus)、硫磺菌(Laetiporussulphureus)来源于中国普通微生物菌种保藏管理中心(CGMCC),灰葡萄孢(Botrytiscinerea)来源于南京林业大学森林病理实验室。试验线虫为南京林业大学森林保护实验室保存的松材线虫AMA3种群。
1.3 木腐真菌的室内筛选
1.3.1 木质纤维素降解菌的初步筛选 将腐朽松木样品中分离得到的147个真菌菌株,打孔直径5 mm菌块分别接种到PDA-愈创木酚(0.04%)、PDA-苯胺蓝(0.1%)和羧甲基纤维素钠(CMC-Na)平板上,各设置3个重复,在25 ℃恒温暗培养5天,根据愈创木酚平板显色圈和苯胺蓝平板脱色圈大小,判断各菌株木质素降解能力。采用刚果红染色法,测量羧甲基纤维素钠平板菌落产生水解透明圈直径与菌落直径,根据比值大小判断各菌株纤维素降解能力(孙思琦等,2020)。综合3种选择培养基测定结果,筛选出优良木质纤维素降解菌株。
1.3.2 松材线虫平板繁殖量测定 将初步筛选出的木腐真菌接于PDA平板上,待菌株长满平板,每皿接入2 000条松材线虫,以灰葡萄孢菌作为阳性对照,每种真菌5皿重复。将供试虫株AMA3与不同木腐真菌在25 ℃恒温培养箱中共培养8天后,采用贝尔曼漏斗法对平板中的松材线虫进行分离,统计不同木腐真菌平板上松材线虫的数量变化。
1.3.3 木腐真菌腐朽能力测定 木腐真菌腐朽能力测定方法参照GB/T 13942—2009《木材耐久性能第1部分:天然耐腐性实验室试验方法》和骆静怡等(2015)方法加以改进:将黑松疫木段切割为2 cm×2 cm×2 cm木块作为腐朽试验试样,称量每块木材样品绝对干质量(精确到0.01 g)。室内条件下将无菌且具相同规格的黑松木块接种至长满木腐菌的广口瓶中,每个广口瓶接2个木块,每个菌株设置3个重复,4个月后用毛刷和流水将木块冲洗干净,除去表面菌丝,放入(103±2)℃烘箱烘至恒质量,计算不同真菌侵染下木块的失重率,反映不同木腐真菌的木材腐朽能力。
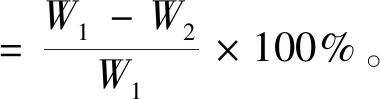
式中,W1:试验前木块绝干质量;W2:试验后木块绝干质量。
1.4 木腐真菌田间接种试验
参照蒙海勤等(2020)的方法加以改进,沿用液体PDA摇培菌丝接种方法。选择当年感病松材线虫病病死木,伐倒除侧枝,锯成75 cm长木段,每棵树3~5个木段。将所有砍伐木段按粗细大小均匀分组并编号,放置于林地上备用。用35 mm钻头在木段上均匀打4孔作为接种点,孔深约55 mm。接种前喷洒75%乙醇消毒,每个孔接种50 mL液体菌丝,用液体PDA浸湿的棉花球盖住孔口,透明塑料薄膜缠绕封口。试验设置7个木腐菌处理组,1个空白对照组,每个处理12个木段(来自3~4棵黑松疫木)。处理4个月后在垂直于接种点平面的木段侧面5点均匀钻孔取样,分析整段疫木内部松材线虫数量变化。取样结束后用油锯将75 cm木段横向一分为二,将所有处理组及对照组的其中一半木段(长约37.4 cm)分别放入8个20目2 m×2 m钢丝网罩中,扎丝封口,另一半木段置于网罩外。待天牛开始羽化,将8组网外木段纵向劈开找寻天牛蛹室,取天牛蛹室周围0.5~1 cm木样,分析蛹室周围木样内松材线虫数量。同时定期捕捉网罩内羽化的天牛成虫,分析天牛身上携带线虫数量。
所有取回的木样及天牛均通过贝尔曼漏斗法分离其中线虫数量,Leica DM500显微镜下计数,并将漏完虫的木样置于(103±2)℃烘箱中烘至质量恒定,称量木样绝干质量,计算每克木样中所含线虫量。
1.5 优良木腐真菌的初步鉴定
利用CTAB法提取优良木腐菌株基因组DNA,选用真菌通用引物ITS1/ITS4进行PCR扩增ITS序列。用浓度为1%的琼脂糖凝胶验证PCR产物,验证成功的产物由上海杰李公司测序。将测序序列在NCBI上进行BLAST比对。
1.6 数据统计与分析
利用IBM SPSS Statistics 20软件处理和分析数据,表格中计算结果均用mean±SEM表示,采用单因素方差分析(One-Way ANOVA)方法比较各处理间差异显著性,GraphPad Prism 6.01软件作图。
2 结果与分析
2.1 高效抑制疫木中松材线虫繁殖的木腐真菌筛选
2.1.1 具有木质纤维素降解能力的菌株筛选 147个真菌菌株中有12个真菌在愈创木酚平板上产生红棕色氧化带,9个真菌在苯胺蓝平板上产生脱色圈,14个真菌在刚果红羧甲基纤维素钠平板上产生透明圈(表1)。其中菌株J5-2、J22、J46、H12、S4、S8、S14、W4、W15、X4、Z1、Z17、Z25在1种以上的选择培养基中能表现出变色反应,说明这些菌株可以同时分泌多种木质纤维素降解酶系,具有较好的分解木质纤维素潜力。从单个培养基分析,在愈创木酚平板上菌株W15显色能力最强,其次为菌株W4、J46、Z25。在苯胺蓝平板上菌株Z43、S4、S8对苯胺蓝的脱色能力与其他菌株相比具有显著差异(P<0.05)。在羧甲基纤维素钠平板上菌株S17、W15、H12产生水解圈与菌落直径比值较大,说明菌株产生纤维素水解酶的含量较高。综合3种选择培养基变色反应结果,初步筛选Z1、Z17、Z25、Z43、J5-2、J22、J46、H12、S4、S8、S14、S17、X4、W4、W15共15个菌株进行下一步试验。
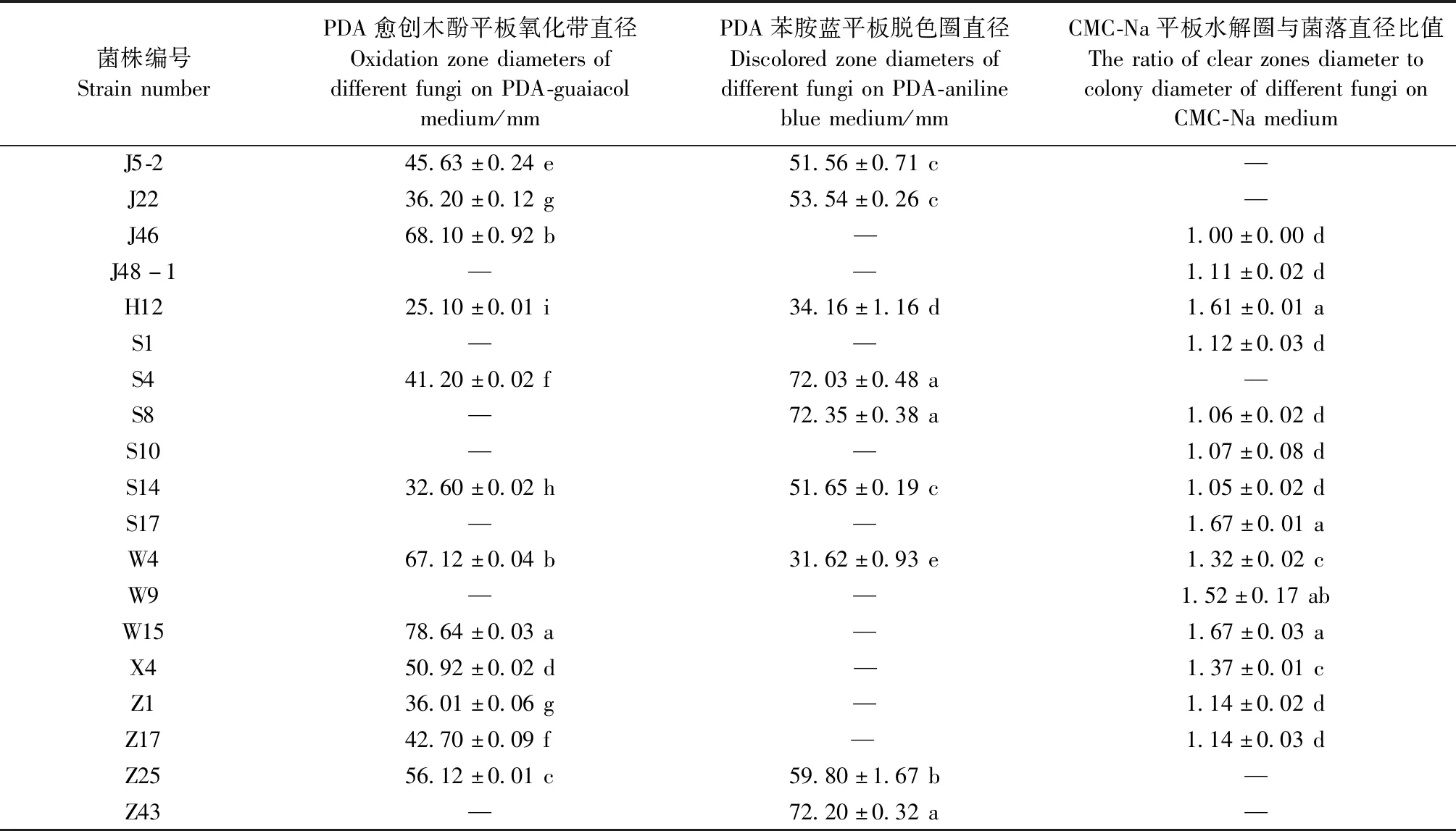
表1 不同木腐真菌在选择培养基上的变色反应①Tab.1 Color reaction of different wood rot fungi on the selected medium
2.1.2 具有抑制松材线虫生长繁殖能力的菌株筛选 将筛选的15个木腐真菌及硫磺菌、糙皮侧耳在PDA上培养成熟后,接入2 000条松材线虫,另用灰葡萄孢作为阴性对照菌株,共培养8天后,统计分析松材线虫在不同木腐真菌平板上的数量变化(图1)。菌株H12和对照菌株灰葡萄孢上线虫的繁殖量与其他菌株具有显著差异(df=17,F=13.743,P<0.05),松材线虫在菌株H12平板上数量增多约52倍,达每皿(103 967±28 395),在灰葡萄孢上增多约50倍,说明菌株H12能够促进松材线虫的大量繁殖,是室内培养松材线虫的潜力菌株,不适用于松材线虫病的疫木处理。在菌株S17、Z43、Z17、S8、S14、J22、W4上松材线虫数量增多,但显著小于对照菌株灰葡萄孢,其余菌株松材线虫数量均明显减少。其中在J5-2菌落上培养8天后,平均每皿仅分离出18±10条线虫,而在硫磺菌和糙皮侧耳菌落上则完全分离不到松材线虫。最终选择在平板培养上能够明显抑制松材线虫繁殖的9个菌株,即X4、Z1、W15、J46、S4、Z25、J5-2、硫磺菌、糙皮侧耳等进行下一步试验。

图1 不同木腐真菌培养8天后松材线虫的数量Fig.1 The number of B. xylophilus cultured by different wood rot fungi after 8 daysB. c:灰葡萄孢B. cinerea;L. s:硫磺菌L. sulphureus;P. o:糙皮侧耳P. ostreatus.下同。* 表示差异显著(P<0.05),虚线代表初始线虫量(2 000条)。* indicate significant difference among all treatments at 0.05 level,the dotted line shows the initial number of nematodes(2 000 pieces).The same below.
2.1.3 具备快速降解松木能力的木腐真菌筛选 将以上筛选出的9株木腐真菌进行松木试样侵染试验。由图2可知,在9个菌株处理中,有8个处理黑松木块的质量损失与对照相比具有显著差异(df=9,F=32.961,P<0.05),其中硫磺菌降解松木的能力最强,侵染黑松4个月后木材试样失重率达到22.82%,与其他处理相比差异显著;其次是菌株S4,造成木块质量损失21.68%;菌株J5-2和X4分别使木块质量减少了17.45%和15.43%;菌株W15、Z25、糙皮侧耳分别使木块质量减少了8.28%、6.76%和6.51%;菌株J46对木块降解较少,木块失重率为2.26%;而菌株Z1对木块失重影响最小,与对照组无显著差别。木材侵染试验表明,硫磺菌、S4、J5-2、X4降解木材能力较强,菌株W15、Z25、糙皮侧耳次之,菌株J46、Z1木材降解能力较弱。
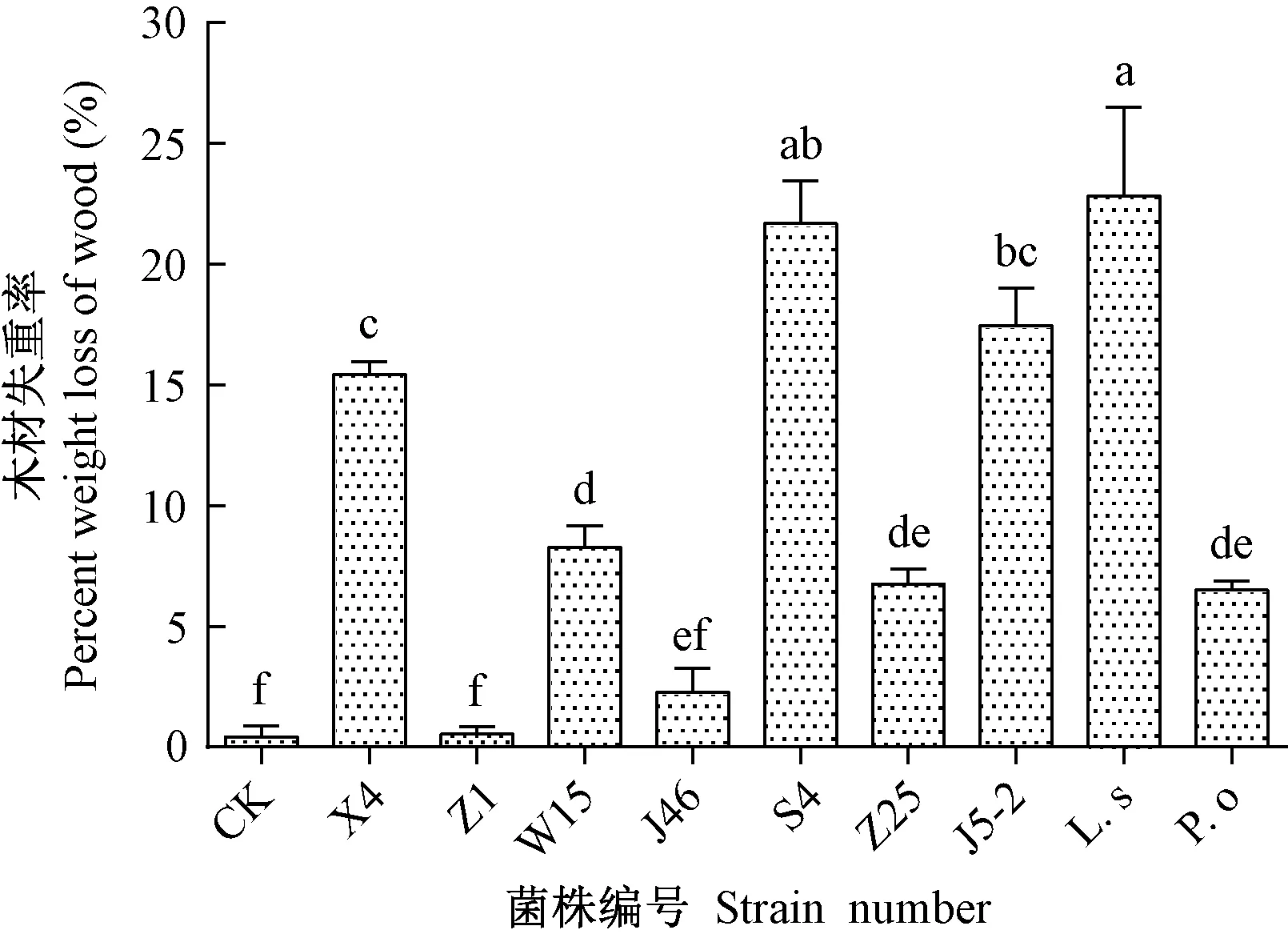
图2 木腐真菌接种后黑松木块失重率Fig.2 Weight loss of Pinus thunbergii wood block after inoculation with wood rot fungi不同小写字母表示差异显著(P<0.05)。Different lowercase letters indicate significant difference among all treatments at 0.05 level.
综合室内筛选结果,选取具有抑制松材线虫繁殖且具备较强松木降解能力的菌株X4、W15、S4、Z25、J5-2、硫磺菌、糙皮侧耳进行野外接种试验,进一步探究利用木腐菌株进行田间疫木除害的能力。
2.2 利用木腐真菌进行田间疫木处理的除害效果
2.2.1 接种木腐真菌120天后疫木内松材线虫数量的变化 将不同木腐真菌通过液体菌丝就地处理疫木4个月后,对比初始疫木线虫量,8组疫木内松材线虫数量均减少。其中对照组减少39%,可能因松木树皮脱落、水分流失等多种环境因素导致线虫种群数量下降。由图3可知,7个施菌处理组均可使松材线虫数量降低65%以上,与对照组相比差异显著(df=7,F=4.579,P<0.05),其中菌株J5-2使疫木木段内线虫数量减少最多,减少率达79%,其次是硫磺菌和糙皮侧耳,线虫减少率分别为73%、72%;菌株X4、S4、Z25、W15使线虫数量减少67%~70%。不同菌株处理4个月后疫木内线虫数量减少率均大于对照组,各菌株间差异不显著(P>0.05),不同处理组需取蛹室周围木样进一步探究其除害效果。
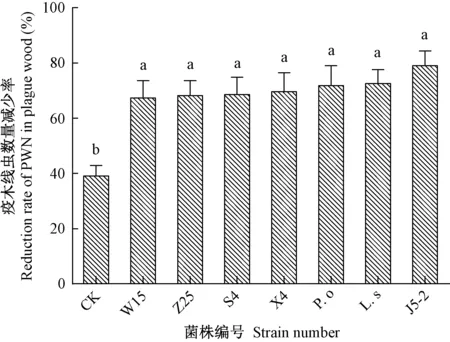
图3 木腐真菌接种120天后疫木内松材线虫减少率Fig.3 Reduction rate of PWN in plague wood after inoculated with wood rot fungi for 120 days不同小写字母表示差异显著(P<0.05)。Different lowercase letters indicate significant difference among all treatments at 0.05 level.
2.2.2 疫木接菌处理对天牛蛹室松材线虫数量的影响 2020年5月初,即试验地天牛羽化初期,木腐真菌处理后蛹室线虫数量变化率如图4。由于天牛蛹期疫木内扩散性松材线虫具有向蛹室聚集的效应,未施菌对照组蛹室线虫数量与初始量相比增加44%,而7种木腐真菌接种5个月后造成疫木内蛹室周围线虫数量不同程度的消减。其中,菌株J5-2处理后疫木蛹室线虫数量减少72%,其次为硫磺菌和菌株S4,蛹室周围线虫减少率分别达69%、68%。菌株W15、Z25、X4处理后蛹室线虫数量分别降低37%、36%、32%,糙皮侧耳使蛹室线虫数量下降22%。
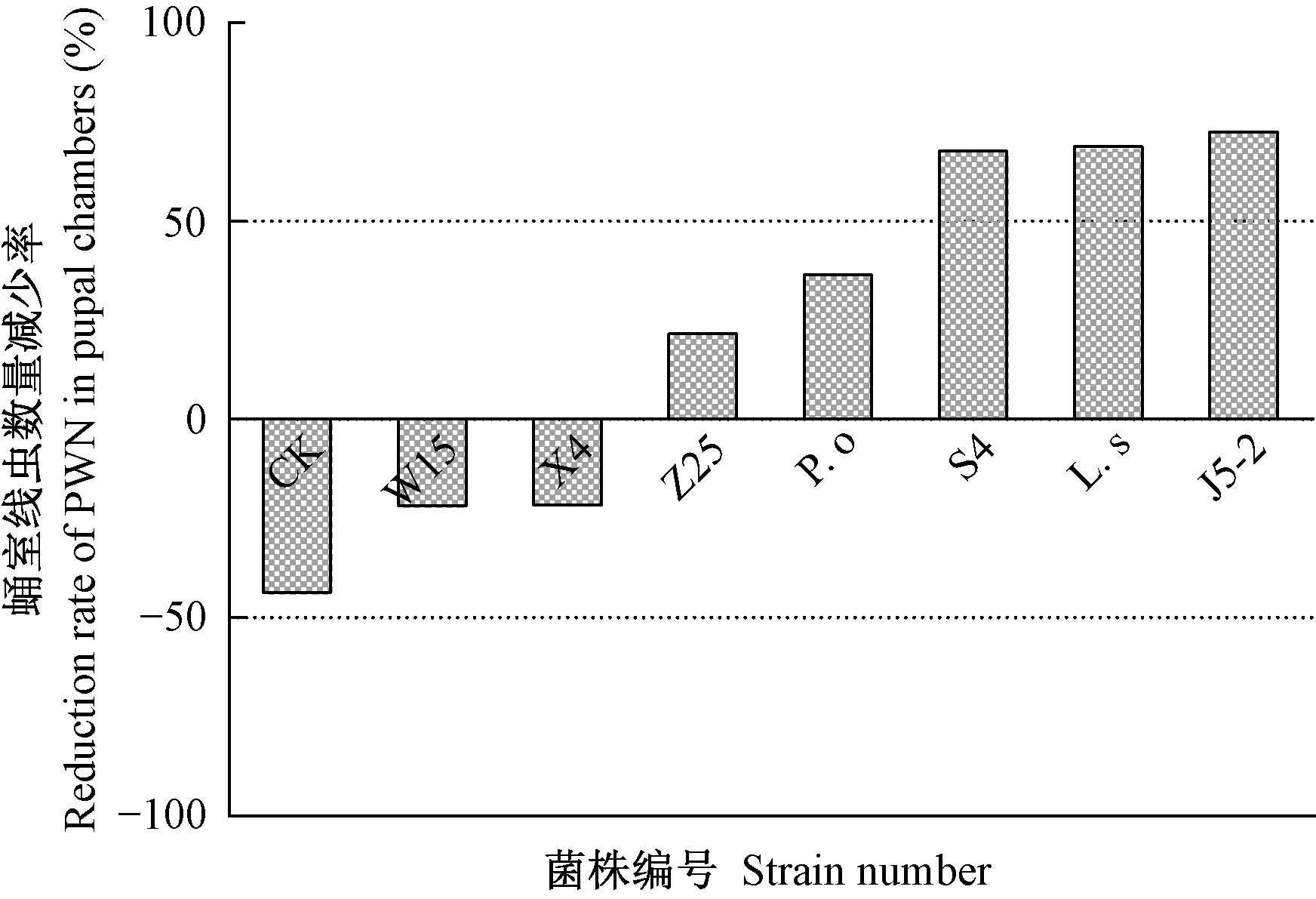
图4 不同木腐真菌处理后疫木内蛹室周围松材线虫数量变化Fig.4 Change in the number of PWN around pupal chamber after inoculating the plague wood with different wood rot fungiL. s:硫磺菌L.sulphureus;P. o:糙皮侧耳P.ostreatus.
2.2.3 疫木接菌处理对羽化后的松墨天牛携带线虫量的影响 试验地5—6月为天牛羽化高峰期,在8组网罩中共捕捉214只松墨天牛成虫,对天牛身体上的松材线虫携带率及携带数量进行检测。由表2可知,在未接菌的对照组疫木中羽化的松墨天牛,其携带松材线虫的数量范围为120~23 400条,平均每只天牛携带线虫数量为(4 396±1 143)条,显著高于施菌处理组(df=7,F=7.013,P<0.05)。菌株J5-2处理下疫木内天牛平均携带线虫数量以及天牛最高携带数量均最少,平均每只天牛携带(15±4)条,最高仅携带60条线虫。其次是硫磺菌及菌株S4,平均携带线虫量分别为每只(55±6)条、64±19条,3个菌株最高携带线虫量均低于200条。7株木腐真菌处理疫木均能使疫木木段内羽化天牛携带线虫量显著下降,且与疫木及蛹室周围线虫减少量统计结果一致,说明室内筛选的不同木腐真菌处理可以消减疫木内松材线虫,进而减少天牛携带线虫数量。
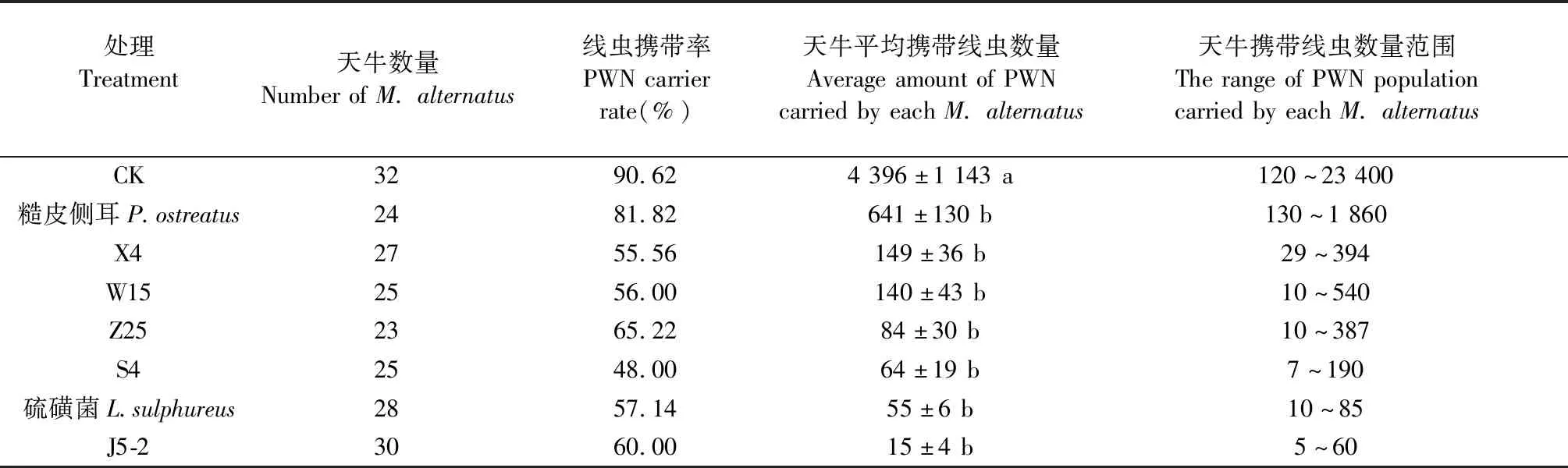
表2 不同真菌处理后疫木中羽化后的天牛上携带松材线虫情况①Tab.2 The number of PWN carried by M. alternatus emergenced in plague wood after treated with different wood rot fungi
不同处理组与对照组天牛平均携带线虫数量分布存在差异。由图5可知,对照组和糙皮侧耳处理组中存在3个极端异常值,除此之外对照组与各施菌处理组的平均数普遍高于其中位数,说明各组中均存在少数天牛携带线虫数量较高的情况。根据箱型图上下限分布可知,对照组天牛成虫平均携带线虫数量分布离散程度最高,中位数集中在900条。施菌处理组中除糙皮侧耳外,其余6个菌株天牛成虫平均携带线虫数量上限值均低于600条,中位数低于300条。综合分析,处理组中菌株S4、硫磺菌、菌株J5-2天牛平均携带线虫数量最低,数据在均值上下集中分布,稳定性高。
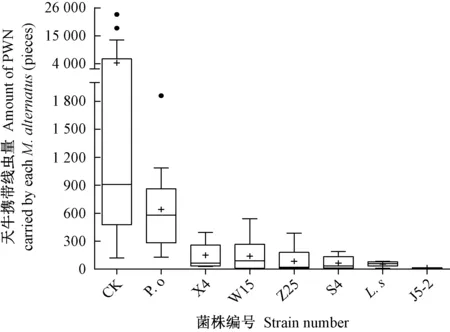
图5 不同木腐真菌处理后天牛平均携带线虫数量Fig.5 The average number of PWN carried by M. alternatus after inoculated with wood rot fungi “·”表示数据异常值;“+”表示数据平均值。“·”indicate outliers,“+”indicates average value.
2.3 优良木腐真菌的初步鉴定
结合室内和田间试验结果,将效果最优菌株J5-2进行基因组测序,登陆NCBI数据库,将测序拼接后的菌株J5-2序列进行Blast比对,结果显示菌株J5-2与Ceriporia相似度最高,为100%,初步鉴定菌株J5-2为多孔菌科(Polyporaceae)蜡质菌属(Ceriporia)一种。
3 讨论
本研究通过室内试验筛选出7株能够有效降解松木并且抑制松材线虫繁殖的木腐真菌,根据松材线虫病流行规律,选择病原及媒介天牛均处于病死木树体中的最佳除害时期,即11月至翌年4月,人工接种优良木腐真菌,使其在野外快速定殖并占据疫木内微生物群落生态位,根据抑减疫木体内松材线虫的数量和松墨天牛成虫的线虫携带量,进一步验证室内筛选菌株的田间除害效果,为筛选松材线虫病疫木处理潜力菌株提供了新的方法思路。
在平板抑虫试验中,松材线虫在硫磺菌、糙皮侧耳等菌株上完全不能存活,这与陈瑶等(2008a)、张建平等(2003)、向红琼等(2000)的试验结果一致。蒙海琴等(2020)发现在茯苓的菌落上也完全分离不到松材线虫。邓习金等(2014)和陈瑶等(2008b)研究发现硫磺菌菌株对疫木伐桩具有较强的分解力和定殖力,并对伐桩中线虫有抑杀作用。在本研究中,硫磺菌侵染松木试样4个月后失重率达22.82%,显著高于其他试验菌株,且田间试验结果表明,硫磺菌处理后疫木内蛹室线虫数量减少69%,羽化天牛平均携带线虫数量少于100条,验证了硫磺菌的良好田间应用潜力。在蒙海琴等(2020)的报道中,茯苓液体菌丝和固体菌剂接种150天后,分别可使疫木内松材线虫含量减少74.51%和65.78%。在野外接种1个月后,笔者发现硫磺菌接种点附近产生大量橘黄色孢子,菌丝体生长没有J5-2菌株旺盛,硫磺菌在野外疫木环境中菌丝延伸能力没有菌株J5-2强,从而导致田间效果不及菌株J5-2好。一些木腐真菌在室内平板上虽具有抑杀线虫的效果,但田间效果不理想,如糙皮侧耳,其对松木的降解率仅为6.51%,田间试验使蛹室线虫数量减少22%。可能由于针叶树大多具萜类及脂类等物质,具有天然耐腐性,对木腐真菌的生长起强烈的抑制作用(Lahretal.,2013),这也是许多阔叶树上常见木腐真菌无法成功定殖在松属植物上的原因,故前期菌株筛选工作可以从野外腐朽松木上分离能够在松树上定殖的木腐真菌,以达到预期的接种试验效果。疫木中真菌的种类可以影响树体中松材线虫数量及天牛携带线虫的数量(Maeharaetal.,2000;2002;郑雅楠等,2014),当适合于线虫繁殖的真菌普遍存在时,会导致树体中线虫密度增加,更多的线虫在蛹室周围聚集,最终天牛携带的线虫数量增加(Maeharaetal.,2005),而当拮抗真菌占优势时,蛹室周围线虫密度下降,导致天牛携带的线虫密度下降(Maeharaetal.,1997)。本研究证实接种不利于线虫繁殖的木腐真菌并使其在疫木中大量定殖,能够降低蛹室线虫密度,从而使天牛携带线虫数量明显下降。研究表明,松墨天牛携带松材线虫的密度决定病害的严重程度(Futai,2010)。当天牛携带10 000多条线虫时,它们可以在5天内将平均1 500条线虫传播到健康树木的嫩枝上,而携带1 000~9 999条中等数量线虫的天牛在传播曲线的峰值平均传播370条线虫。当天牛携带不到1 000条线虫,即使聚集到一棵树上也无法致病(Togashi,1985;2014)。在早期的研究中,Hashimoto(1974)发现接种300条以上松材线虫会导致8年生松树发病,说明若天牛有效传播300条线虫到树体即可致病。本研究除糙皮侧耳菌株外,其余6个木腐真菌处理后的疫木中松墨天牛携带线虫量范围均低于1 000条,菌株J5-2、硫磺菌、S4处理组天牛携带线虫数量均低于200条,说明3种木腐真菌具有疫木除害处理的潜力,施菌处理阻碍了疫木内松材线虫向媒介天牛聚集,减少了天牛携带线虫量,从而降低松材线虫向健康木上的传播效率和可能性。由于真菌的生长受环境因素影响大,同时媒介天牛传播松材线虫具有累加效应,通常一棵树会有几只或几十只天牛取食或产卵,实际防病效果还应该在林间应用后进行评估。
本研究中首次发现蜡质菌属Ceriporia真菌对松材线虫繁殖有抑制作用,是处理松材线虫病疫木的潜力菌株。对于优良菌株在疫木中抑制线虫的具体机制,其代谢产物是否影响线虫与天牛间的化学信号的传导以及不同木腐真菌混合接种的除害效果等一系列问题需要开展进一步的探究。此外,由于松材线虫病疫区不断向北迁移,木腐菌的实际应用需要考虑到施用菌株的低温适应性。本研究林间试验期间11月至翌年4月所在地平均温度为11~13.3 ℃,需要筛选更多耐低温的试验菌株以保证在北部低温地区也能够具备良好的除疫效果。蒙海勤等(2020)的研究证实,液体摇培菌丝接种具备保湿性好、菌丝量大、成本低等特点,优于固体菌丝(袋)接种。但前期准备液体菌丝工作量较大,今后可以针对具备林间应用潜力的菌株进行培养基优化及菌剂开发,提高生物量,扩大林间接种量,促进菌丝向蛹室的延展,从而达到更好的除害效果。
4 结论
本研究中菌株J5-2(Ceriporiasp.)、硫磺菌和糙皮侧耳等木腐菌对松材线虫的抑制作用明显,硫磺菌和菌株S4降解木材的能力强,菌株J5-2、硫磺菌、S4的林间效果最好,可使疫木中线虫数量减低65%以上。利用木腐真菌是松材线虫病疫木就地处理的有效途径,值得继续深入研究。
