藏东南嘎朗国家湿地公园华山松种内与种间竞争强度研究
2021-08-05姚慧芳
姚慧芳,卢 杰,王 超,陈 康
(1.西藏农牧学院 高原生态研究所,西藏 林芝 860000;2.西藏高原森林生态教育部重点实验室,西藏 林芝 860000;3.西藏林芝高山森林生态系统国家野外科学观测研究站,西藏 林芝 860000)
竞争是2个及以上的有机体在可利用资源、环境和空间不足的情况下所产生的相互关系[1-2],林木竞争是指在同一生长环境中的林木,因阳光、水分等生长发育所必须的条件而产生争夺共有资源的情况,这也是植物群落内部个体关系的主要形式之一,竞争会导致一部分林木因资源缺乏而生长缓慢甚至死亡[3-5]。研究发现,竞争是森林群落中树木之间的一个基本过程,它对于种群存活、种群动态及群落演替具有重要作用[6]。一些林分中,竞争已经成为植被生长和生产力的决定因素之一[7]。在众多关于计算树种间竞争强度的指标中,最为典型和常用的是F.Hegyi于1974年提出的与距离有关的Hegyi单木竞争指数模型[8-9]。
华山松(Pinusarmandii)属松科(Pinaceae)松属(Pinus),因产于陕西华山而得名。华山松适生海拔范围 1 000~3 300 m,广泛分布在云、贵、川、青、台、藏等地,在西藏主要分布在雅鲁藏布江下游。华山松具有很高的经济、生态和药用价值,松木材质轻软,耐水、耐腐,且易于加工,工业上松香、松油也是重要原料;在干旱半干旱地区,可以起到涵养水源,保持水土,防止风沙等作用;华山松的种子球果都可用来入药,对于治疗咳嗽、咳痰以及润肺通便具有重要作用[10]。本文以藏东南华山松为研究对象,采用Hegyi单木竞争模型定量分析其种内与种间竞争关系,明确在群落中种内和种间竞争的比重,进而分析藏东南华山松生长和发展动态,以期为该区华山松的后续研究提供参考。
1 研究区概况
藏东南是指西藏的东南部,其地形、地貌上主要由高山深谷和阶地构成,由于印度洋的暖湿气流沿着雅鲁藏布江大峡谷浩荡北上,因此该区域的降水量较多,气候温和,形成诸多森林植被类型[11],这里也是青藏高原海拔最低的地区。研究区地处藏东南林芝市波密县嘎朗国家湿地公园内(29°90′68.03″N,95°60′61.77″E),园区主要由嘎朗湖构成,嘎朗湖是地壳运动的结果,属自然形成,湖区面积2km2。园区内以针叶林为典型,也是华山松分布较为集中的地方。乔木层除华山松之外还伴生有高山松(Pinusdensata)、林芝云杉(Picealikiangensisvar.linzhiensis)、川滇高山栎(Quercusaquifolioides)等,灌木层主要有卫矛(Euonymusalatus)、三桠乌药(Linderaobtusiloba)、小檗属(Berberis)、白簕(Acanthopanaxtrifoliatus)等,草本植物主要有卵萼花锚(Haleniaelliptica)、草玉梅(Anemonerivularis)、唐松草(Thalictrumaquilegifoliumvar.Sibiricum)、天南星属(Arisaema)等。
2 研究方法
2.1 数据采集
2020年9月初,在全面踏查的基础上,根据华山松的分布和具体生境,在波密县嘎朗国家湿地公园内设置样地5个,每个样地的面积为30 m×30 m。对5个样地的总体情况进行调查并记录:1)样地的海拔为 2 731 m,坡度为18°,坡向为北358°,郁闭度达0.90,盖度为95%;2)调查样地内灌木、草本的种类;3)参照刘万生等[12]、高浩杰等[13]方法,对样地内胸径≥5 cm的乔木进行每木检尺,包括树种、位置、胸径、树高等;4)竞争范围参照李帅锋等[14]、黄小波等[15]和骆文建等[16]方法,根据林隙半径、上层树的影响范围和样地的树高及冠幅确定竞争木距离范围,在样地调查中,发现林窗半径为8 m左右,因此将竞争范围确定为8m。
2.2 对象木和竞争木的选取
选取对象木时要考虑边缘效应的影响,尽量不选取样地边缘的树作为对象木,根据毛磊等[17]方法,在样地边缘建立缓冲区,在该缓冲区内不选取对象木,沿着样地周围建立8 m的缓冲区。鉴别竞争木通常运用固定半径法,首先选择对象木作为原点,以r=8为半径画1个圆,竞争木就是与对象木距离小于r的树木。
2.3 径级结构
起测胸径为5 cm,并以5 cm为径阶距,按照胸径大小的范围,划分为9个径级(DBH)较为合理[2]:第1径级(5 cm≤DBH<10 cm),第2径级(10 cm≤DBH<15 cm),第3径级(15 cm≤DBH<20 cm),依次类推,划分至第8径级(40 cm≤DBH<45 cm),第9径级(DBH≥45 cm),统计各径级对象木和竞争木的数量、百分比、平均胸径和平均树高。
2.4 竞争指数的确定
根据对象木与其对应竞争木的相对坐标计算出对象木与其对应竞争木之间的距离Lij[18],公式如下:
(1)
式中:xi,yi为对象木i的坐标值;xj,yj为竞争木j的坐标值。
采用与距离有关的Hegyi竞争指数模型计算种内与种间竞争指数大小,计算公式为:
(2)
式中:CI为竞争指数;Di为对象木胸径;Dj为竞争木胸径;Lij为竞争木和对象木之间的距离;m为竞争木的株数。CI值越大,表示对象木受到的竞争压力越大。
幂函数回归模型:
CI=AD-B
(3)
式中:CI为竞争强度指数;D为个体胸径;A和B为模型参数。
竞争分为种内和种间竞争,种内竞争是华山松与相同树种之间的竞争,种间竞争是对象木与群落内的其他树种之间的竞争,因此竞争木有两类,种间和种内的竞争强度之和为对象木的总竞争强度。
2.5 数据处理
采用Excel进行统计和计算,在SPSS 19.0中分析对象木胸径与竞争指数的关系,回归分析得到最优模型,并绘制相关图表。
3 结果与分析
3.1 对象木与竞争木
在嘎朗国家湿地公园共调查对象木39株,最小胸径为5.1 cm,最大胸径为65.29 cm。将所调查的对象木和竞争木按径级分组统计(表1)。在5 cm≤DBH<10 cm径级时,对象木株数最多,占总数量的35.90%;其次为10 cm≤DBH<15 cm,25 cm≤DBH<30 cm,35 cm≤DBH<40 cm和DBH≥45 cm径级的对象木株数较多,占总数的10.26%,对象木小径级的数量最多,总体上随着径级的增加株数在减少。
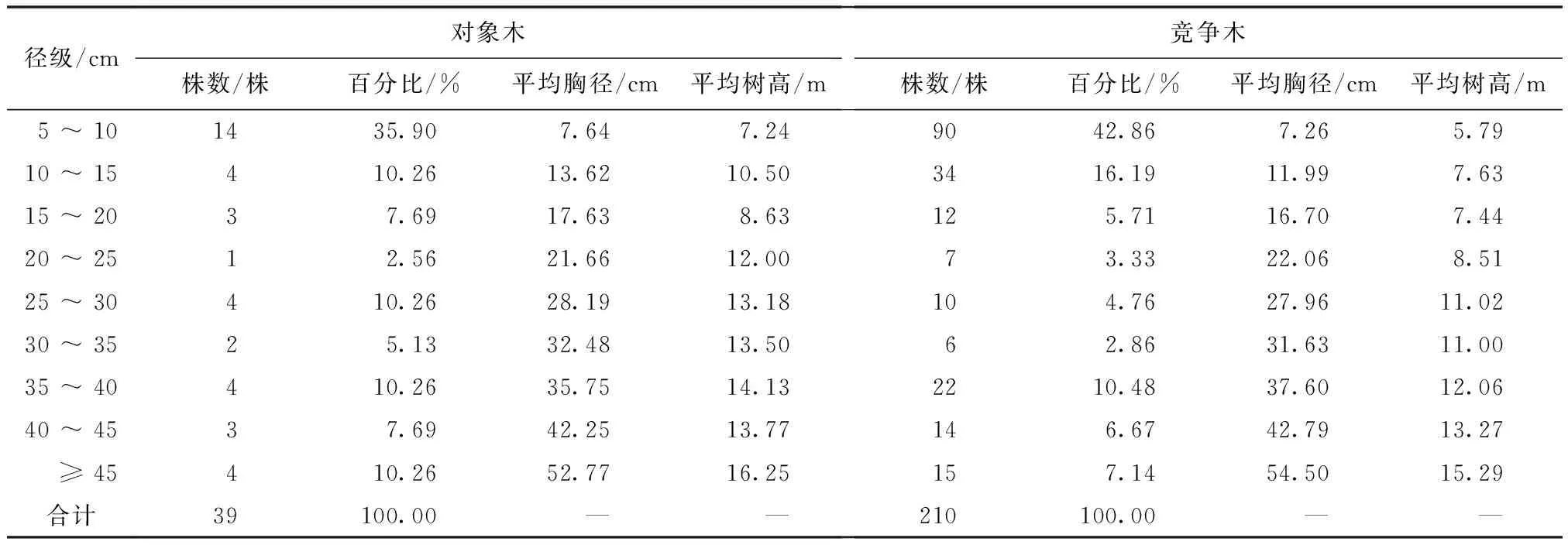
表1 对象木与竞争木的概况
竞争木共4种210株,种间竞争木包括高山松、川滇高山栎和林芝云杉,最小胸径为5.1 cm,最大胸径可达85.99 cm。在5 cm≤DBH<10 cm径级的竞争木数量最多,占总数量的42.86%;其次为10 cm≤DBH<15 cm径级的竞争木株数较多,占总数量的16.19%;30 cm≤DBH<35 cm径级的竞争木数量最少,仅占总数的2.86%。竞争木的株数大致随径级的增大先减后增,其中大径级的竞争木大部分是高山栎。
3.2 华山松种群树高与胸径的关系
林分中树高和胸径是2个非常重要的因子,利用所测的胸径和树高值,自变量为胸径,因变量是树高,由此绘制散点图(图1)。通过散点分布趋势,作回归分析,发现其较好地服从幂函数关系(y=2.9853x0.4249,R2=0.6939,P<0.0001):华山松的幼树数量最多,随着径级的增加,树高生长较快,最高可达18 m,其年龄结构属于增长型种群。
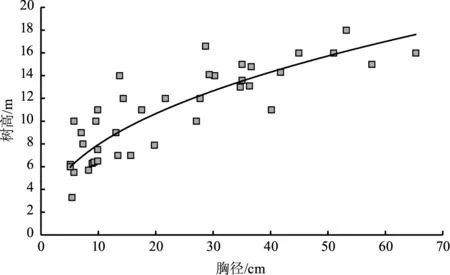
图1 研究区华山松胸径与树高的关系
3.3 华山松的种内与种间竞争
华山松受到的种内竞争强度为37.748,占总竞争强度的54.68%;种间竞争强度为31.284,占总竞争强度的45.32%,种内竞争强度大于种间,说明华山松的竞争主要来自于种内(表2)。华山松的竞争强度大小与径级大小及植株数量关系密切,中小径级的个体及数量较多的个体所受到的竞争强度较大,随着径级的增大,平均竞争指数呈明显减少趋势。在华山松的整个生长阶段中,在径级5~15 cm和30~35 cm时,华山松的种内和种间的竞争强度都较大,分别占总竞争压力的82.25%和92.69%,种内和种间的竞争强度也随着对象木径级的增加大致减小,这与华山松的实际情况也较符合。华山松个体胸径随着其自身的生长发育不断增大,树高也不断增加,由此其核心竞争能力也在不断增强,尤其生长发育到成熟阶段,冠幅较大,矗立于主林层,对地上和地下的可利用资源和空间占有绝对优势,而竞争木对华山松的竞争强度逐渐减弱,并趋于稳定。
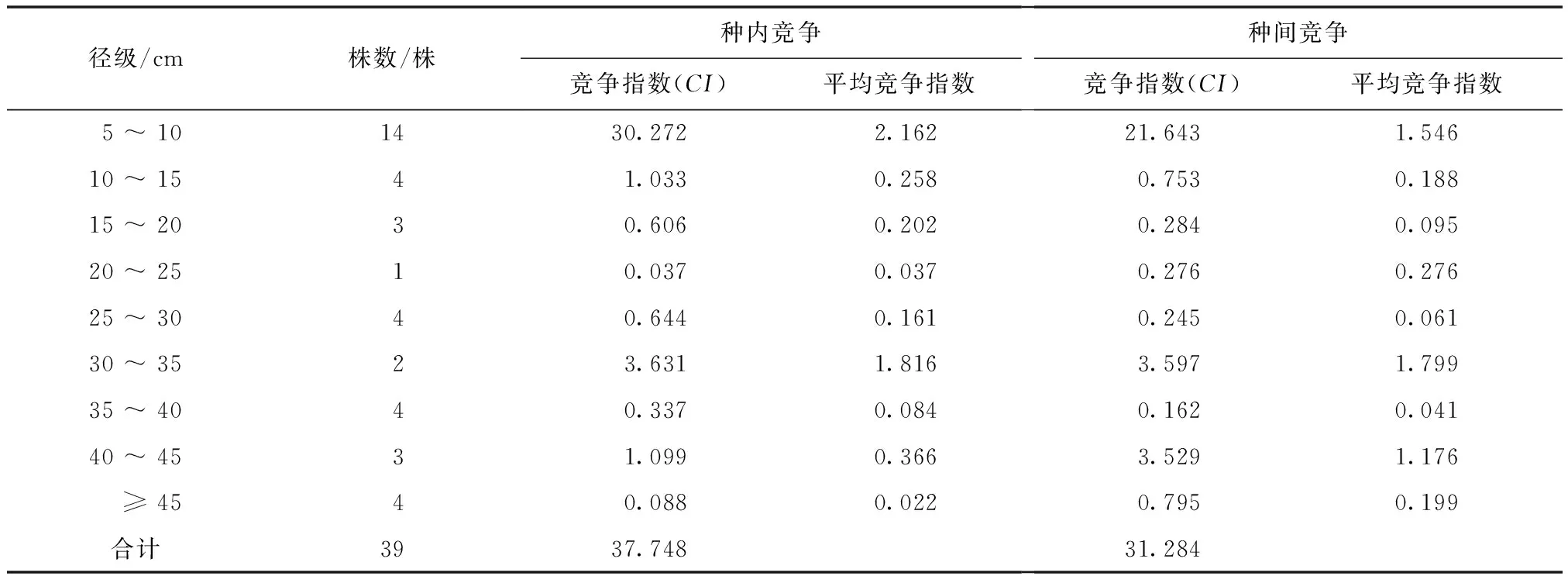
表2 华山松种内与种间竞争强度
除竞争木的大小和数量影响华山松的种间竞争程度外,竞争木的种类不同其竞争程度也存在较大差异(表3)。在调查的3种伴生种中,竞争力最大的是川滇高山栎,竞争指数为15.259,原因可能是川滇高山栎为阔叶树种,其冠幅较大,很大程度上影响了对象木对资源的利用;其次为林芝云杉,竞争指数为8.962;竞争最小的为高山松,竞争指数为7.063,仅占总竞争指数的22.58%。华山松的主要伴生种间的竞争顺序依次为川滇高山栎>林芝云杉>高山松。

表3 华山松竞争木的种类组成及竞争强度
3.4 华山松胸径与竞争指数的关系及其预测
植株的竞争能力受到植株个体冠幅、胸径的大小、生长的阶段等多种因素的制约,其中对林木影响最大的因子是植株个体胸径的大小[14]。为研究藏东南华山松个体所受的竞争指数大小与自身生长的相关关系,以对象木胸径为自变量,采用线性、指数、对数、幂函数等多种数学公式对竞争指数与对象木胸径间的关系进行回归拟合。研究发现,对于华山松胸径与整个林分(图2)、华山松胸径与伴生树种(图3)、华山松胸径与种内(图4)竞争指数的关系,在所采用的数学模型中,幂函数的显著性水平均达到极显著,拟合优势度也最大,故采用幂函数回归模型。
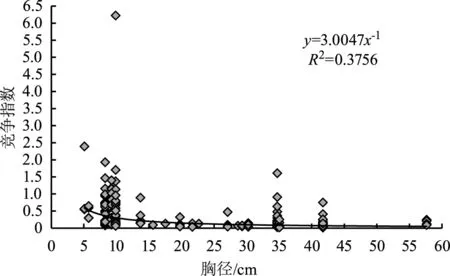
图2 华山松胸径与整个林分的竞争指数关系
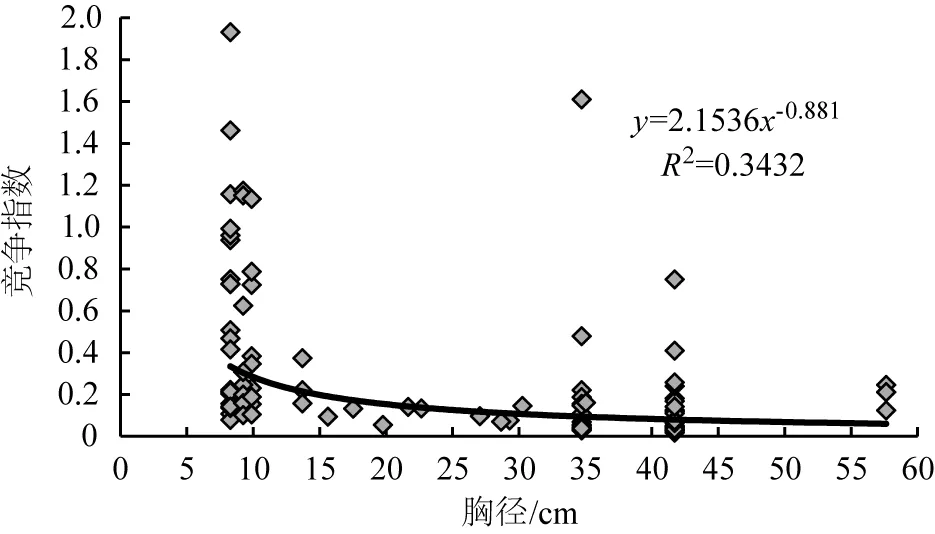
图3 华山松胸径与伴生树种的竞争指数关系
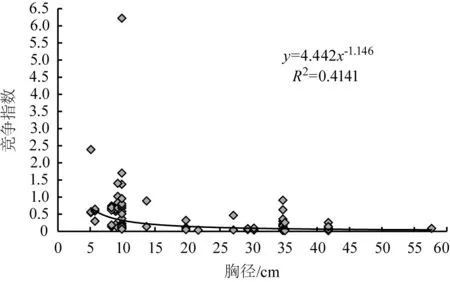
图4 华山松胸径与种内的竞争指数关系
模型通过显著性检验,结果表明均达极显著水平(表4)。对39株对象木胸径与整个林分、伴生树种、种内的竞争指数关系进行分析,发现模型的幂值均为负值,即对象木的个体越大,受竞争木的竞争作用就越弱,竞争指数逐渐减小,但胸径在35~45 cm之间的竞争指数骤然增加,这可能是因为35~45 cm径级对象木对应的竞争木数量较大,具体原因还有待进一步研究。
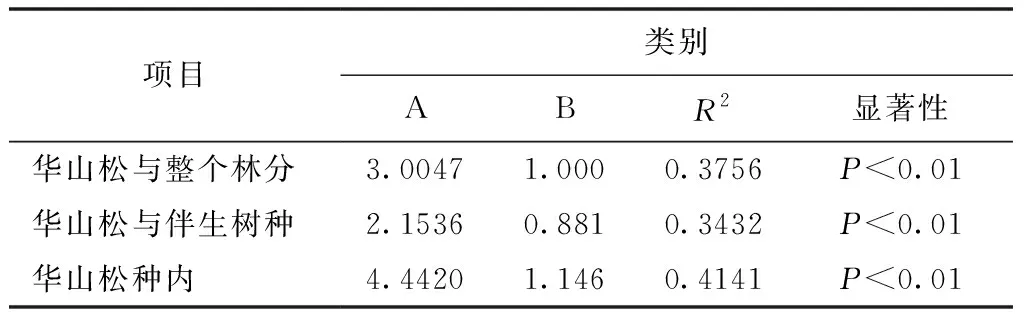
表4 竞争强度与对象木的胸径模型参数
通过对华山松个体胸径与其竞争强度的模型预测表明(表5),随着华山松个体胸径的不断增大,与之对应的种内和种间的竞争指数不断下降。当胸径超过25 cm时,下降的幅度趋缓,幂函数的回归模型对于预测华山松的竞争强度较合理。
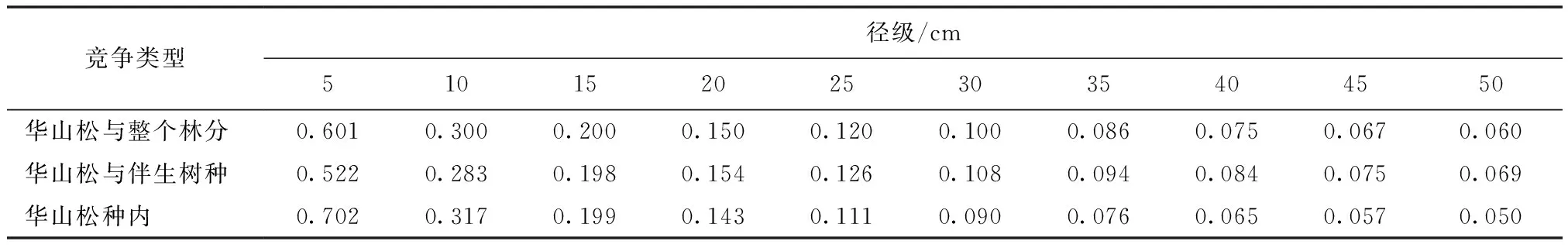
表5 华山松种内种间竞争强度与对象木胸径的模型预测
4 讨论与结论
4.1 讨论
4.1.1华山松种内与种间竞争强度
1) 林木之间的竞争主要表现在其争夺可利用资源和空间时所产生的相互关系[19]。Hegyi竞争指数模型是研究植物种内与种间竞争关系最常用的模型之一。虽能够反应出个体对资源的竞争程度,并且能够更好地将其数量化[20],但它是一个静态分析过程,具有局限性,在今后的研究中需要结合动态数据,例如气候和空间分布等。
2) 在藏东南华山松天然群落中,种内竞争强度大于种间,即自疏作用大于他疏作用。种内竞争主要是因为相同物种的资源利用的普遍重叠。在研究影响林木竞争能力的因素中,生态习性与生态位越接近的物种,对资源的竞争越激烈,是因为其占据相同或相似的生态宽度;在一个植物生态系统中,占主导地位的物种,其竞争能力也最强。这一现象与华南五针松(Pinuskwangtungensis)[2]、长白落叶松(Larixolgensis)[7]、云南松(Pinusyunnanensis)[21]、兴安落叶松(Larixgmelinii)[22]等植物的竞争状况较为相似,都是以种内竞争为主,但并不代表其可以和伴生种稳定共存,需进一步研究植物之间的共存机制。
3) 华山松的种间竞争因树种不同而受到的竞争强度也不同。在3种伴生种中,与华山松竞争强度较大的树种是川滇高山栎(15.259),由于其是阔叶树种,冠幅大,在可利用资源方面占较大优势,因此最大程度地影响了华山松对象木对于资源的利用。林芝云杉(8.962)和高山松(7.063)作为针叶树种其竞争强度相对较弱。
4.1.2华山松个体胸径与竞争强度
1) 华山松对象木个体胸径与整个林分、伴生树种及种内的竞争强度关系都符合幂函数模型(y=3.0047x-1,y=2.1536x-0.881,y=4.442x-1.146),幂值均为负数,说明随着华山松胸径的增加,3种竞争强度均总体减小。原因是华山松林中小径级个体数量占大多数,冠层被遮挡,影响个体的生长发育,所受的竞争压力大;大径级华山松个体在种群中处于上层,竞争能力增强,所受的竞争压力变小。但胸径在35~45 cm之间的华山松竞争指数骤然增加,这可能是由于该径级华山松的竞争木数量较大,需进一步研究具体原因。
2) 预测华山松每一径阶距胸径的竞争强度表明:华山松胸径从5~50 cm的过程中,其对应的3种竞争强度均呈下降趋势;当胸径大于25 cm时,趋势变缓。这是由于华山松个体不断发育,到达成熟林阶段时处于主林层,成为优势种,能够更好地利用资源和空间,因此竞争木对其影响减弱,竞争强度趋于稳定。
4.2 结论
小径级华山松数量最多,种群属于增长型,为防止种群内部出现断层现象,因加强保护中龄林个体;华山松天然林群落以种内竞争为主,且其3种伴生树种的竞争强度顺序依次为川滇高山栎>林芝云杉>高山松;华山松胸径与整个林分、伴生树种、种内竞争指数均呈负相关关系,随着胸径的增加,其竞争指数逐渐减小。本研究将有助于认识藏东南华山松种内与种间复杂的竞争过程,了解华山松生长动态。在未来的经营管理中,需充分考虑林木的竞争关系,采用合理措施,促进藏东南华山松种群的天然更新。
