药用万寿菊多倍体的诱导与特征分析
2021-07-20符勇耀杨利平郑开敏徐文姬
符勇耀 杨利平 郑开敏 徐文姬

摘 要:为获得四倍体药用万寿菊材料,采用秋水仙素对药用万寿菊‘内蒙1号的种子进行多倍体诱导研究。以0.15%浓度的秋水仙素处理露白的万寿菊种子24 h,幼苗变异率较佳,可达到56%。幼苗定植于田间,初步筛选并标记形态变异植株,自花授粉后采收种子播种并观测子代形态指标,对形态变异明显的植株进行流式细胞仪检测,确定获得15株纯合四倍体。通过将四倍体与二倍体进行比较,四倍体植株茎粗、栅栏组织厚度、栅栏组织与海绵组织的比例和单朵花重量显著高于二倍体植株,四倍体植株总叶量、叶宽、叶长、叶长宽比、叶片气孔密度和单株花朵数量显著低于二倍体植株。获得的四倍体万寿菊植株单株花总重量比二倍体显著增加。
关键词:万寿菊;多倍体;育种;诱变
中图分类号:S567 文献标识码:A
Abstract: To obtain the material of polyploid medicinal marigold, colchicine was used for the polyploid induction with the seeds of medicinal marigold ‘Neimeng No. 1. The results showed that 0.15% of colchicine treated for 24 h was the best selection of the seedling induction of Tagetes erecta, reaching 56% of mutation rate. The seedlings were planted in the field and preliminary screening and marking of morphological variant plants were carried out. Seeds were harvested after pollination and seeded and morphological indicators of progenies were observed. Flow cytometry was used to detect the plants with obvious morphological variation and a total of fifteen tetraploid plants were identified. By contrast, the stem diameter, palisade tissue thickness, ratio of palisade tissue to spongy tissue and single flower weight of tetraploid plants were significantly higher than those of diploid plants, and the total leaf weight, leaf width, leaf length-width ratio, leaf stomatal density and number of flowers per plant of tetraploid plants were significantly lower than those of diploid plants. The total flower weight per plant of tetraploid plants was significantly increased in contrast to that of diploid plants.
Keywords: Tagetes erecta; polyploid; breeding; induction
DOI: 10.3969/j.issn.1000-2561.2021.05.017
万寿菊(Tagetes erecta)为菊科万寿菊属一年生植物,原产墨西哥等美洲地区[1]。根据其用途可分为花用万寿菊和药用万寿菊两大类,其中药用万寿菊花瓣中的叶黄素含量很高,约占花瓣色素总质量的90%,因此万寿菊成为提取天然叶黄素的普遍植物材料[2]。药用万寿菊产业的终端产品叶黄素具有抗氧化、抗白内障、防止老年视黄斑退化、降低患癌率和防治冠心病等用途,在保健食品、化妆品、医药等行业领域普遍应用[3]。目前,辽宁、内蒙古和云南等省(区)是我国药用万寿菊的主要栽培地,万寿菊种植和叶黄素的提取加工已经成为地方脱贫致富的特色产业[4-7]。随着现代医药科技的发展,人们对叶黄素产品的需求逐渐增加,除了扩大种植面积来满足市场需求外,提高万寿菊鲜花单位面积产量和叶黄素含量成为当前研究的热点之一。目前,在提高栽培技术和杂交育种筛选高产品种提高万寿菊产量方面已有较多报道,如有学者筛选出适应种植地气候条件的高产品种和栽培技术,以提高万寿菊植株的花产量[8-9],也有育种者利用杂交优势培育出万寿菊花朵产量或色素含量均有提高的品种[10-12]。
通常多倍体植物的一些性状会比二倍体更加优良,比如植株高大、产量提高和适应性增强等,因此多倍体育种已经被廣泛应用于农业生产,并取得良好效果,如小麦(Triticum aestivum)、油菜(Brassica napus)和棉花(Gossypium spp.)等多倍体作物产量明显提高、适应性增强[13],药用植物四倍体金银花(Lonicera japonica)和板蓝根(Baphicacanthus cusia)有效成分含量比普通二倍体高出约40%~60%[14-15]。目前国内生产中所用的药用万寿菊品种多为二倍体(2n=2x=24),有关万寿菊多倍体的研究鲜有报道。刘金凤等[16]采用化学诱导万寿菊幼苗生长点成功获得变异株;He等[17]利用秋水仙素处理万寿菊雄性不育系(M525AB)并获得四倍体万寿菊材料,但多倍体后代的形态表现和农艺性状未见描述,栽培实践中未见多倍体万寿菊品种出现。农业企业主要以采摘万寿菊鲜花出售获得经济效益,增加万寿菊单株鲜花产量或提高叶黄素含量成为育种的主要目标之一。培育多倍体万寿菊材料,有可能获得鲜花产量增加的新种质,进而培育获得万寿菊多倍体新品种,对于药用万寿菊产业发展具有重要意义。
1 材料与方法
1.1 材料
以重庆市黔江区木莲山种养殖股份合作社基地提供的‘内蒙1号药用万寿菊二倍体(2n= 2x=24)种子为材料。
1.2 方法
1.2.1 多倍体诱导及栽培 2017年春精选颗粒饱满、无病虫害的万寿菊种子,清水浸泡催芽处理至稍露白,以0.05%、0.10%、0.15%、0.20%的秋水仙素溶液避光浸泡,震荡处理12、24、48、60、72 h,每组处理50粒种子,清水处理50粒种子为对照。重复3次。秋水仙素处理后的种子用清水冲洗3次后,点播在72穴的育苗盘,混合基质(V草炭∶V蛭石=3∶1),幼苗4片真叶时分为2组进行露底高畦定植,株行距為50 cm × 50 cm,定植后幼苗死亡及时替换。播种和定植之后常规水肥管理,株高30 cm时统一去除顶芽。
1.2.2 多倍体初步筛选 诱变的万寿菊种子长出胚芽后,观察胚轴和子叶的状况,与空白组进行比较,将下胚轴增粗,子叶增厚,或子叶畸形的植株视为变异苗。待植株长出1对真叶时,统计植株的变异率(即形态学上发生任何改变的幼苗)和存活率(存在已萌发却在一定时间无法生长的或缺失生长点的幼苗不计在内)。当幼苗出现2~3对真叶时,分处理变异组和空白组分组进行露地栽培。植株长势稳定后,将处理过的植株与空白组进行比较,处理组出现叶片加厚或叶色加深的植株,进行挂牌标记,初步确定为变异植株。2017年对初步筛选出的形态变异株万寿菊套袋自花授粉,获得子一代种子(F1)。
2018年春将万寿菊形态变异株子代种子和对照种子育苗、定植,方法同1.2.1。植株长势稳定后,再次将形态变异株子代植株与空白组进行比较,处理组出现叶片加厚或叶色加深的植株,进行挂牌标记,确定为变异植株,做多倍体鉴定材料。
1.2.3 多倍体鉴定 F1代形态变异植株高约40 cm时,选择形态(如叶片颜色、大小,茎干直径或花朵大小等)变异明显的植株,取其中部的叶片,用滤纸夹住并浸湿滤纸,放进封口袋中,注明植株编号,置于装有冰袋的保温箱中,寄送北京德世达科技有限公司进行流式细胞仪检测(检测仪器:德国Partec CyFlow Space试剂盒,Partec CyStain UV Precise P),二倍体为对照。得到染色体倍性测试报告和分析样本的测试图,确定四倍体植株。
1.2.4 万寿菊四倍体和二倍体形态特征比较 分别挑选10株四倍体和二倍体植株,待植株现蕾时分别测量植株总叶量、叶片面积和叶片长度、植株的茎粗、叶片横切栅栏组织与海绵组织的比例,单株花产量统计22株。测量时间为天气晴朗的早晨9:00—10:00。
(1)茎粗和总叶量。利用游标卡尺,对植株主茎的基部向上第4节处进行茎的直径测量。记录植株基部丛生枝的数量,再挑选中等大小1枝丛生枝,记录其侧枝的对数和第1对侧枝、第4对侧枝和靠近顶芽的最后1对侧枝的叶量,以三者的平均值乘以侧枝的对数,再加上顶芽周围的叶量,可得到1枝丛生枝的平均叶量,经计算,即可得到植株总叶量。
(2)叶片形态及叶面积。采摘每株茎中部的新鲜叶片并清理干净,保持原状粘贴在透明胶带上,用叶面积仪(杭州托普有限公司YWJ-B160- 10)进行测量,记录单叶裂片的面积、长度、宽度和长宽比,每组叶片用叶面积仪重复测3次。
(3)气孔特征。分别选取四倍体和二倍体植株茎中部的叶片(朝南),获取单叶尖部裂片中部的上、下表皮制作表皮临时装片,观察气孔密度和气孔大小。
(4)叶片横切面栅栏组织与海绵组织的比例。取四倍体和二倍体植株茎中部单叶叶片(朝南),用清水将叶面清洗干净,摘下相同位置的单叶裂片制作徒手切片,显微镜下测量叶片横切面的栅栏组织与海绵组织的比例。
(5)单株花产量。万寿菊植株进入始花期开始对四倍体和二倍体植株每一株的花朵数量进行统计,每周记录1次,直到最后一朵花盛开。在同一植株花期的早、中、晚三个阶段各取1朵发育成熟的盛花,称重,取平均值。
2 结果与分析
2.1 多倍体诱导
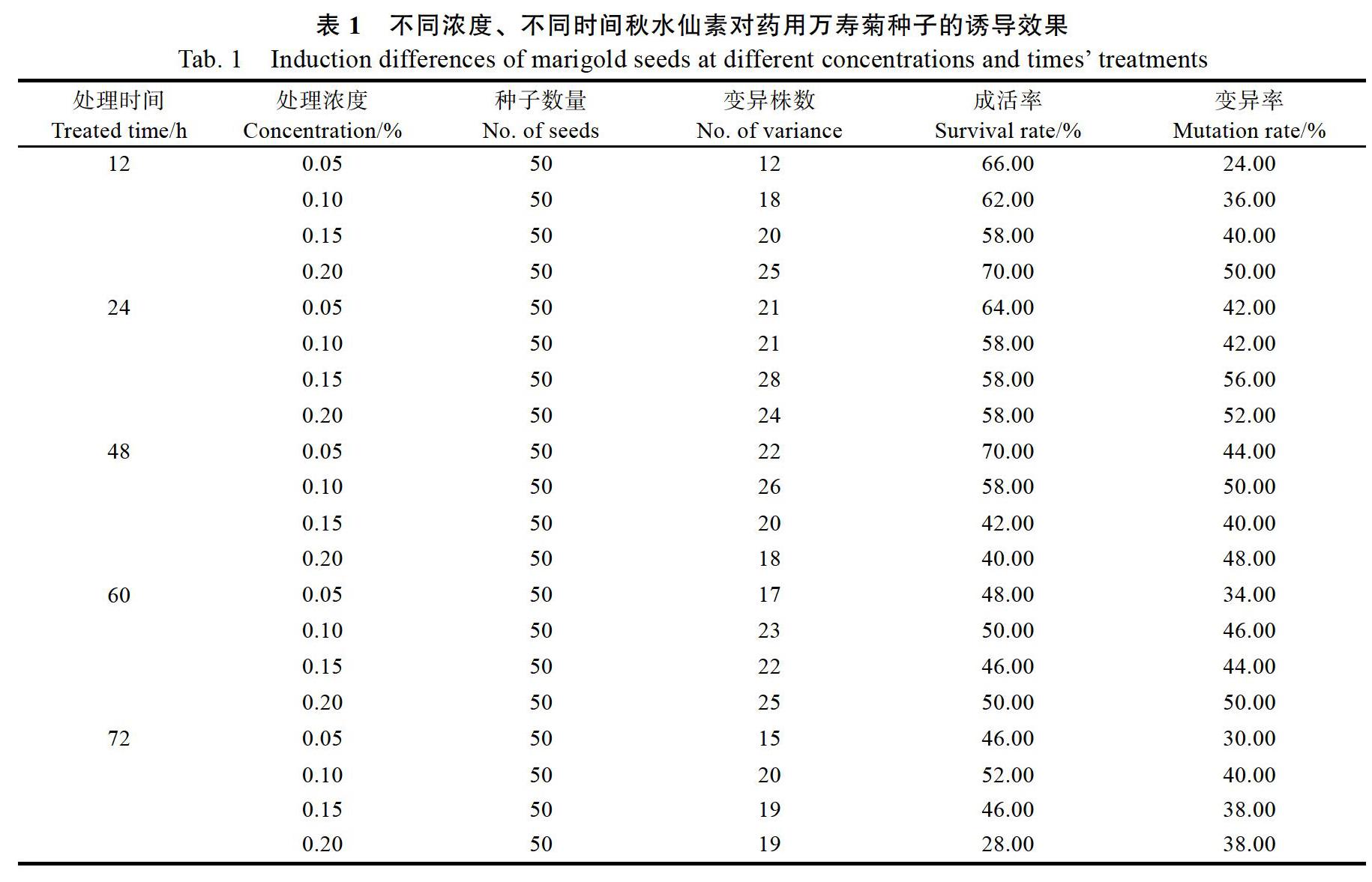
2.1.1 秋水仙素对万寿菊诱导的效果 万寿菊种子经秋水仙素处理后进行容器育苗,约12 d左右出苗,诱导过的幼苗出苗整齐度低于对照。播种后第25天统计幼苗存活率和变异率,变异苗出现多种特征,如两片子叶的边缘相连接、两片子叶一大一小、子叶变厚有棱、真叶畸形、下胚轴增粗、胚根短、生长点损坏、发芽时间晚和生长缓慢等(图1)。不同秋水仙素处理诱导万寿菊变异率存在一定的差异。当处理时间为24 h,秋水仙素浓度为0.15%时,其幼苗诱变率最高,为56%;当处理时间为12 h,秋水仙素浓度为0.05%时,诱导效果最差,诱变率为24%(表1)。随着处理时间的增加,不同浓度秋水仙素诱变率先增加后降低;在较短的处理时间内(12 h和24 h),处理浓度越高,变异率越高;在较长处理时间下(48 h,60 h和72 h),处理浓度增加,变异率增加不明显。随着处理时间的增加,各处理的幼苗存活率降低,在秋水仙素浓度为0.20%时,表现最明显(表1)。
2.1.2 多倍体田间初步筛选 2018年7月下旬田间栽培的万寿菊植株开始现蕾,植株生长基本稳定。观察发现变异植株与对照相比其叶片深绿色,叶片较厚无韧性,单叶上的裂片宽大,叶柄和茎秆较粗,总叶量偏低且植株生长缓慢。这些形态明显发生变异的植株,可初步确认为多倍体,进入下一步检查鉴定。
2.2 多倍体鉴定
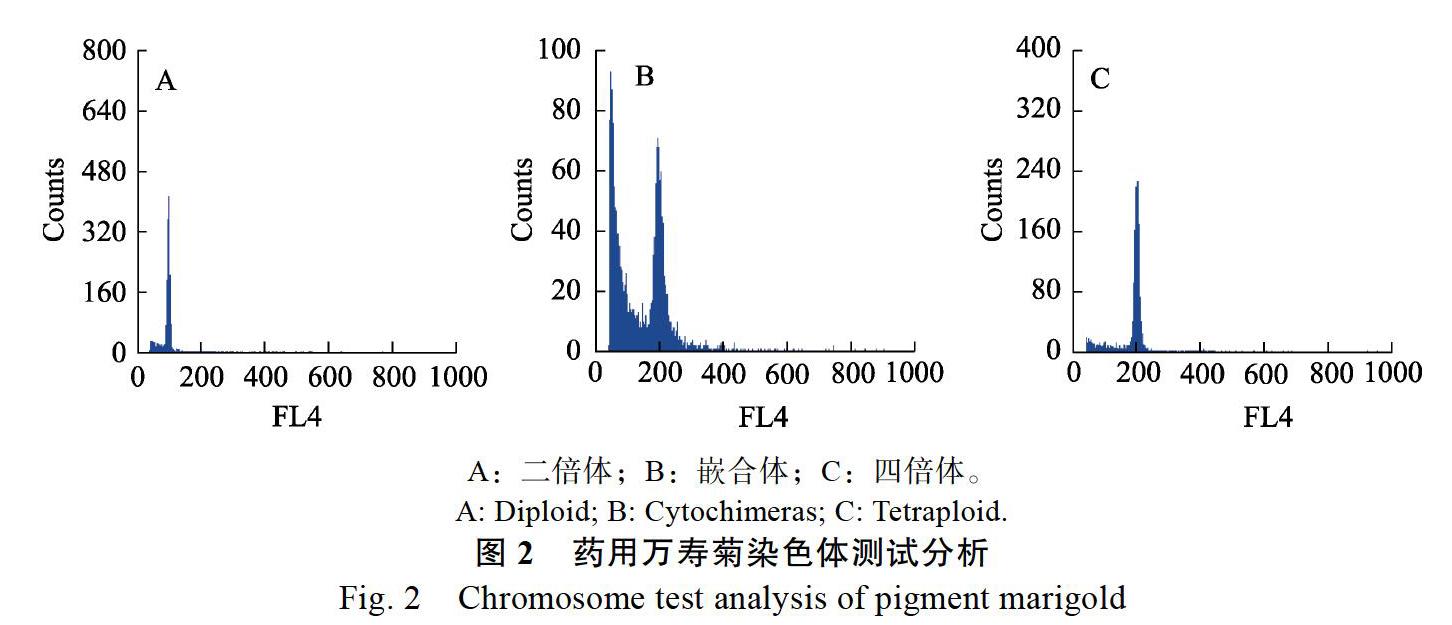
对经初步筛选得到的形态变异植株进行标记,通过与二倍体植株对比,选取叶片厚度显著增加、叶色深绿色或生长紧凑的植株进行流式细胞仪鉴定,将二倍体植株叶片作为参照,由样本的荧光强度X-Mean判断倍性,将参照二倍体峰设在100,则四倍体峰在200,单倍体在50。在筛选的150株明显变异的植株中,编号36、67、78、81、82、88、89、90、92、95、98、100、101、104、105、106、108、116、152、115、117、148的植株在相对荧光强度值50~200的位置出现2个峰(图2B),表明这22株为嵌合体;编号12、59、62、66、71、80、91、96、97、102、110、118、120、121、114的植株在相对荧光强度值200的位置出现1个峰(图2C),表明这15株为纯合四倍体;其余植株(除编号64)在相对荧光强度值100的位置出现1个峰(图2A),表明其为二倍体。
2.3 四倍体和二倍体植株形态特征
2.3.1 茎粗、叶量和叶面积 四倍体和二倍体植株在叶量、茎粗等性状上有明显的差异。四倍体植株的茎干变粗,总叶量减少,其中四倍体植株茎粗比二倍体植株增加0.18 cm,总叶量仅为二倍体的28.55%(表2,图3A,图3B)。四倍体万寿菊与二倍体相比,单片叶面积未发生显著变化;叶片的长度、宽度及长宽比与二倍体万寿菊植株存在显著差异(表3)。
2.3.2 叶片微形态 万寿菊四倍体与二倍体植株的叶表皮气孔密度、气孔长度、气孔宽度和气孔面积均具有显著差异,而四倍体植株叶表皮气孔长宽比未发生显著变化。四倍体植株气孔密度明显小于二倍体,其中叶上表皮气孔密度是二倍体的47.22%,叶下表皮气孔密度是二倍体植株的60.80%,其上表皮气孔长度和宽度比二倍体植株分别增加了50.34%和51.12%(图4,表4,表5)。
栏组织增加,植物光合能力增强。四倍体植株叶片栅栏组织厚度、栅栏组织与海绵组织的比例与二倍体植株具有显著差异,四倍体万寿菊植株栅栏组织比二倍体植株增厚了58.86%,四倍体植株海绵组织厚度与二倍体植株相比差异不显著(图5,表6)。
2.3.3 单株鲜花产量 鲜花产量是万寿菊育种目标之一,在其他栽培条件相同的情况下花朵大小、叶黄素含量和单株产量显得最为重要。分析发现四倍体万寿菊植株平均单朵花的花径明显大于二倍体(图3 C,图3 D),单朵花的重量显著高于二倍体,其平均每株花朵的数量显著低于二倍体植株(表7)。四倍体平均单株花产量为772.25 g,二倍体平均单株花产量为705.22 g,四倍体单株花产量比二倍体显著增加。四倍体植株与对照的花期没有显著差异。
3 讨论
‘内蒙1号万寿菊种子经过0.15%秋水仙素诱导24 h效果最好,幼苗存活率为58%,诱变率达56 %。幼苗和田间成年植株形态初步筛选出形态明显发生变异的植株,但初选时当代变异株比较多,并且植物形态变化上具有一定的连续性;同时也发现当代变异植株通常生长缓慢,可能受到秋水仙素处理的影响,不利于选择多倍化表型。为此,蕾期对变异株套袋并自花授粉后采收种子播种并观测子代形态指标,子代中出现150株形态变异株,流式细胞仪检测确定筛选的个体中有15株为纯合四倍体,并且这15株的茎、叶的形态和微形态符合多倍体的基本特征,其中四倍体植株茎粗、栅栏组织厚度、栅栏组织与海绵组织的比例和单朵花重量显著高于二倍体,四倍体植株总叶量、叶宽、叶长、叶长宽比、气孔密度和单株花朵数量显著低于二倍体植株。确定的四倍体不仅茎干粗壮抗倒伏,而且单株鲜花产量显著高于二倍体,基本达到了育种目标。此外,子代中也出现嵌合体,占14.7%,其表型主要是介于四倍体和对照组之间,与前人研究中也发现个别植物嵌合体自交可以获得嵌合体后代相似[18-19]。二倍体植株的比率占到了75.3%,说明有可能当代变异植株的染色体数目并没有发生加倍,大部分形态变异是由于在自交过程中,染色体发生重组引起的变化。有研究者用秋水仙素浸泡处理花用万寿菊雄性不育系(M525AB)种子,发现浓度0.05%,处理3 h和6 h存活率为41%~50%,变异率为89%[17]。本试验中使用秋水仙素的浓度提高了3倍,处理时间增加4~8倍,而存活率相当,诱变率略低。两者之间差异很大,产生差异的原因可能是万寿菊不同品种基因型导致。
单倍体育种技术只需2个世代便可获得纯合的DH系,有效缩短育种周期。通常获得单倍体的途径有利用高效诱导系为父本对目标作物授粉[20-21],也有利用Co?射线辐射授粉获得作物单倍体[22]。本研究在确定获得15株纯合四倍体的同时,也发现变异株子代群体中有7株形态明显矮小、细弱的个体,流式细胞仪检测确定其中的1株(编号64)为单倍体个体。秋水仙素处理万寿菊子代中产生单倍体植株现象未见报道,产生单倍体可能是种子来源的二倍体亲本产生。
染色体计数法鉴定植物倍性是一种较为常用且精准的方法,但制作染色体压片的实验步骤较为繁琐复杂且耗时长,需具备熟练的操作技术,不适合大量样品的检测[23]。采用流式细胞仪鉴定法,样品制备方便、灵敏度高、分析快速、植物损伤性小,是鉴定大量试验样本倍性的最快速有效的方法[24],如用流式细胞仪鉴定非洲菊大孢子再生植株群体的倍性,其准确率可达95%,利用流式细胞仪鉴定诱变后的薄皮甜瓜,再结合气孔特征辅助鉴定,可以快速、准确地确定变异株的倍性[25-26]。采用形态学筛选和流式细胞仪鉴定组合方法,可有效弥补待检测群体较大和万寿菊染色体小,压片计数操作困难的缺点[27],使四倍体万寿菊的鉴定和筛选变得准确高效。
参考文献
[1] 中国科学院中国植物志编辑委员会. 中国植物志(第75卷)菊科二 [M]. 北京: 科学出版社, 1979: 389.
[2] 汪殿蓓, 陈芬芬. 万寿菊叶黄素开发利用研究进展[J]. 北方园艺, 2007(1): 44-46.
[3] 林登贵, 曾 丽, 王 鹏, 等. 万寿菊研究现状及发展趋势[J]. 上海农业学报, 2014, 30(6): 145-149.
[4] 潘润平, 吴凯龙, 赵海荣. 对万寿菊花卉业的思考[J]. 内蒙古农业科技, 2015, 43(2): 86.
[5] 丛 崇. 赴辽宁省彰武县考察色素万寿菊发展情况的报告[J]. 现代农业, 2019(5): 37.
[6] 汪殿蓓, 陈芬芬. 万壽菊叶黄素开发利用研究进展[J]. 北方园艺, 2007(1): 44-46.
[7] 周保华, 李宗林. 永胜县万寿菊种植技术及经济效益分析[J]. 现代农村科技, 2018(2): 17.
[8] 曾 丽, 周叶林, 陈光甫, 等. 万寿菊提取叶黄素专用品种筛选及配套技术研究[J]. 上海交通大学学报(农业科学版), 2002, 20(2): 145-149, 165.
[9] 丁学才, 赵艳霞. 万寿菊高产栽培技术[J]. 北方园艺, 2007(5): 183-184.
[10] 王贵余. 万寿菊保护地杂交制种技术[J]. 中国种业, 2003(10): 59-60.
[11] 张永强, 刘锦荣, 高文学. 色素万寿菊杂交育种后代的性状表现[J]. 中国种业, 2008(7): 47-48.
[12] 李福荣, 张继冲, 续九如, 等. 万寿菊×孔雀草杂交育种及杂种不育性的研究[J]. 内蒙古农业大学学报(自然科学版), 2005, 26(2): 51-54.
[13] 李霖锋, 刘 宝. 植物多倍化与多倍体基因组进化研究进展[J]. 中国科学: 生命科学, 2019, 49(4): 327-337.
[14] 杨映红, 张丽君, 徐小强. ‘九丰一号金银花在天水地区的表现及栽培技术[J]. 农业科技通讯, 2009(12): 168-169.
[15] 张秋红, 闫 滨, 王晓燕. 板蓝根及其多倍体的研究概况[J]. 山东医药工业, 2002, 21(1): 22-24
[16] 刘金凤, 张行杰. 万寿菊多倍体诱导的研究[J]. 中国科技博览, 2014(4): 235.
[17] He Y H, Sun Y L, Zheng R R, et al. Induction of tetraploid male sterile Tagetes erecta by colchicine treatment and its application for interspecifific hybridization[J]. Horticultural Plant Journal, 2016, 2(5): 284-292.
[18] 李天菲. 典型植物嵌合体——鸡冠花嵌合体的遗传学研究[D]. 武汉: 湖北大学, 2002.
[19] Lindsay G C, Hopping M E, Binding H, et al. Graft chimeras and somatic hybrids for new cultivars[J]. New Zealand Journal of Botany, 2011, 33(1): 79-92.
[20] 成 锴, 苏晓慧, 栗建枝, 等. 不同加倍技术加倍玉米单倍体的研究[J]. 玉米科学, 2019, 27(4): 42-46.
[21] 秦 燕, 赵永康, 周华安, 等. 不同来源的糯玉米单倍体诱导研究[J]. 种子, 2019, 38(5): 117-119, 123.
[22] 付文苑, 唐 兵, 邓 英, 等. 辐射花粉授粉诱导黄瓜单倍体及染色体加倍[J]. 分子植物育种, 2019, 17(21): 7150- 7155.
[23] 王荣邦, 张秀海, 吴忠义, 等. 菊花染色体倍性鉴定研究[J]. 安徽农业科学, 2010, 38(23): 12778-12780, 12789.
[24] 万正林, 周艳霞, 武 鹏, 等. 同源四倍体黑皮冬瓜的诱导与倍性鉴定[J]. 华南农业大学学报, 2018, 39(3): 31-38.
[25] 王 康, 何林池, 魏小云, 等. 利用二甲戊乐灵创制薄皮甜瓜同源四倍体[J]. 西北农业学报, 2015, 24(4): 114-119.
[26] 王丽花, 杨秀梅, 吴学尉, 等. 非洲菊大孢子再生植株倍性的快速鉴定方法[J]. 西北农业学报, 2013, 22(1): 155-161.
[27] 赵 慶, 唐道城. 万寿菊根尖染色体观测方法的优化[J]. 北方园艺, 2013(20): 98-100.
责任编辑:沈德发
