82份籼粳稻骨干亲本抗稻瘟病基因的分子检测
2021-07-20王晓玲吴婷唐书升李霞王智权肖宇龙余传源
王晓玲 吴婷 唐书升 李霞 王智权 肖宇龙 余传源


摘 要:北粳南移在江西省已推行11年,為进一步明确适合江西晚粳常规稻的稻瘟病抗性情况及粳渗籼骨干亲本的选择,本研究利用Pi37/Pi35/Pish、Pi5、Pi2、Pi9、Pia、Pi23、Pigm、Pi33/Pi42、Pi56、Pik/kg(t)/ks/kp/km/kh/43、Pikh共11个抗瘟基因的标记结合井冈山抗性谱的鉴定,对28份骨干粳稻和54份骨干籼稻进行分子标记与抗性分析。结果表明,粳稻中有1份携带其中的7个抗性基因,有5份携带6个抗性基因,有7份携带5个抗性基因,有12份携带4个抗性基因,有3份携带3个抗性基因,抗性频率最高的Pia为0.786,其次为Pikh和Pi9,频率均为0.643,Pigm和Pi37/Pish在这批粳稻材料中的抗性频率为0;籼稻中有8份携带其中的7个抗性基因,有10份携带6个抗性基因,有13份携带5个抗性基因,有17份携带4个抗性基因,有4份携带3个抗性基因,有2份携带2个抗性基因,抗性频率最高的Pi9为0.981,其次Pik/Pikh/Piks频率为0.833,Pigm、Pi37/Pish、Pikh、Pia、Pi5、Pi2、Pi23和Pi33/Pi42的频率分别为0.556、0.537、0.519、0.389、0.352、0.315、0.167和0.111,Pi56在这批籼稻材料中的抗性频率为0。明确这些材料抗瘟基因的分布,将为广谱持久抗性籼粳材料创制骨干亲本的选择与配组指明了方向。
关键词:水稻;稻瘟病;骨干亲本;分子标记
中图分类号:S511 文献标识码:A
Abstract: Japonica rice in Jiangxi, China has been planted for 11 years recently. In order to further elucidate the blast resistance of Jiangxi late japonica conventional rice, and select main parents for indica infiltrating japonica rice, 28 main japonica and 54 main indica parents were evaluated in Jinggangshan, Jiangxi with blast resistance gene markers Pi37/Pi35/Pish, Pi5, Pi2, Pi9, Pia, Pi23, Pigm, Pi33/Pi42, Pi56, piks/kg(t)/ks/KP/km/kh/43 and Pikh. Among the 28 japonica rice parents, one contained seven of the eleven resistance genes, five contained six resistance genes, seven contained five resistance genes, twelve carried four resistance genes, and three carried three resistance genes. The dis-tribution frequency of Pia was as high as 78.6%, followed by Pikh and Pi9 with frequency 0.643, but the resistance frequency of Pigm and Pi37/Pish in japonica varieties was 0. In 54 indica rice parents, eight carried seven resistance genes, ten carried six resistance genes, 13 carried five resistance genes, 17 carried four resistance genes, four contained three resistance genes, and two contained two resistance genes. The highest resistance frequency 0.981, was observed in Pi9, followed by Pik/Pikh/Piks, 0.833. The frequency of Pigm, Pi37/Pish, Pikh, Pia, Pi5, Pi2, Pi23 and Pi33/Pi42 was 0.556, 0.537, 0.519, 0.389, 0.352, 0.315, 0.167 and 0.111, respectively. The resistance frequency of Pi56 in indica rice was 0. Identifying the distribution of resistance genes in these materials would indicate the direction for creating indi-ca-japonica materials and selecting main parents for reconstituting broad-spectrum and persistent resistance genes.
Keywords: rice; blast; main parents; molecular markers
DOI: 10.3969/j.issn.1000-2561.2021.05.001
水稻是我国主要的粮食作物,稻瘟病(rice blast)是水稻的三大病害之一,一般可造成水稻减产10%~20%,严重时可导致绝收。稻瘟病菌生理小种的变异快,水稻品种的抗性一般仅能维持3~5年[1]。因此,可以说稻瘟病的抗性关系到水稻的高产、稳产甚至是粮食的安全,水稻稻瘟病的防治至关重要[2-4]。
通过常规杂交、回交将水稻的抗性基因转移到骨干水稻亲本中是最安全和生态友好型的手段[5],目前,水稻中鉴定了100多个抗稻瘟病基因(或等位基因),克隆了抗稻瘟病的基因有37个,鉴定了抗稻瘟病的QTLs有500多个[6],其中Pi9、Pigm、Pi50、pi21、Pi7、Pi57和Ptr等为广谱抗性基因[7]。早在1999年,Hittalmani等[8]就利用RFLP标记对Pi1、Piz-5和Pita进行了精细定位和辅助聚合育种,是稻瘟病较早的分子研究;2008年最早克隆了Pi-ta和Pib两个抗稻瘟病基因[9];而Pigm的鉴定、定位、克隆与应用更是研究广泛[10-15]。另外,水稻抗瘟基因的抗性复杂,一个位点可能有多个抗性等位基因,同一个基因又可能存在多个抗性单倍型[16-22]和不同的蛋白型[16],还会发生假基因化和其他基因的修饰[23],甚至稻瘟病抗性基因在表达上还有上位性互作效应[7],因此,在不同稻区抗瘟基因的应用频率不尽相同。
江西省“籼改粳”的发展思路是2009年提出来的,目前已过去11年,这期间江西粳稻推广生产已取得重大进展,创建了“早籼晚粳”高产高效的生产新模式[24],常规粳稻育种也初见成效,更适合江西生态环境的渗籼型粳稻正在努力培育中。在品质方面,整合籼稻的蓬松、清香和粳稻的軟滑、弹牙的口感是粳稻培育的一个切合点。主要的方向是引入籼稻的抗稻瘟病基因[25]、早生快发基因、较耐高温基因、穗型和粒型基因等在粳稻中的运用。因此,利用分子标记检测用于培育粳稻或渗籼型粳稻的籼粳骨干亲本抗稻瘟病的基因型对于今后粳稻抗稻瘟病育种至关重要。
1 材料与方法
1.1 材料
参试的28份粳稻材料来源于江西、云南、江苏和安徽等省份,参试的54份籼稻材料来源于江西、广东、浙江、云南和四川等省(区),这些材料全部是在江西省农业科学院水稻研究所试验田中试种后挑选出来的,经过株型、生育期、品质、产量及抗性等育种评价指标评价,适合江西稻作生态,所以选取作为育种骨干亲本。
1.2 抗性表型鉴定
1.2.1 穗颈瘟的鉴定 参照王军等[26]的方法鉴定穗颈瘟,采用井冈山自然条件诱病的四级鉴定方法。经2018—2019年鉴定,2年鉴定结果差异不大的取最大的指数计算,差异大于1的数据去除。0级:无病,为免疫;1级:1/4以下枝梗发病或穗颈部有斑点,表现抗病(有个别表型取0.5级);2级:1/4以上枝梗或主轴中部或颈部有病,对产量影响不大,表现中抗;3级:主轴中部或颈部发病,对产量有显著影响,表现感病;4级:穗颈发病造成白穗,表现高感。
1.2.2 叶瘟的鉴定 参照相关文献[17, 27-28]采用九级鉴定方法鉴定,经2018—2019年鉴定,2年鉴定结果差异不大的取最大的指数计算,差异大于2的数据去除。0级:无病斑,高抗;1级:叶片上产生针头状大小的褐点型病斑;2级:稍大病斑褐点;3级:小圆形稍长的灰色病斑,边缘褐色,病斑直径1~2 mm;4级:典型的纺锤形病斑,长1~2 cm,通常局限在两条主脉间,危害面积不超过叶面积的2%;5级:典型病斑,危害面积不超过叶面积的10%;6级:典型病斑,危害面积为叶面积的11%~25%;7级:典型病斑,危害面积在叶面积的26%~50%;8级:典型病斑,危害面积为叶面积的51%~75%;9级:典型病斑,危害面积为叶面积的76%至全叶枯死。0~3级为抗病,5~9级为感病。
1.3 分子标记检测
1.3.1 DNA的提取 采用CTAB的方法提取DNA[1, 23, 5-6],2.0 mL的离心管中,加入2 cm的叶片,加入800 ?L的CTAB提取液,加入钢珠,60HZ研磨30 s,75 ℃水浴30 min,取出,冷却,加入等体积的氯仿,1200 r/min离心10 min,吸取700 ?L的上清液于1.5 mL的离心管中,加入等体积的无水乙醇,4 ℃放置2 h,1200 r/min离心10 min,去上清,倒置,干燥,加200 ?L去离子水,4 ℃保存。
1.3.2 PCR扩增 抗稻瘟病基因的连锁标记和克隆过基因的功能标记引物见表1,全部由生工生物工程(上海)股份有限公司合成,反应体系20 ?L,其中2×Mix 10 ?L,F/R引物各1 ?L,DNA模板1.5 ?L,加6.5 ?L水补齐。扩增程序:采用95 ℃预变性3 min;94 ℃ 30 s,55 ℃ 45 s,72 ℃ 1 min,共35个循环;72 ℃,5 min。
1.3.3 电泳检测 扩增产物采用6%的PAGE胶300 V电泳2 h检测,0.1%的硝酸银染色10 min,蒸馏水清洗2遍,1.5% NaOH(加0.025%四硼酸钠,0.5%甲醛)显色5~10 min,自来水清洗2遍。
1.4 数据处理
采用软件SAS 9.0和Excel 2010进行统计分析,相关性分析、遗传聚类均采用软件SAS 9.0的固定程序完成,采用非加权类平均法(UPGMA)进行聚类分析。
2 结果与分析
2.1 抗稻瘟病基因间的相关性分析
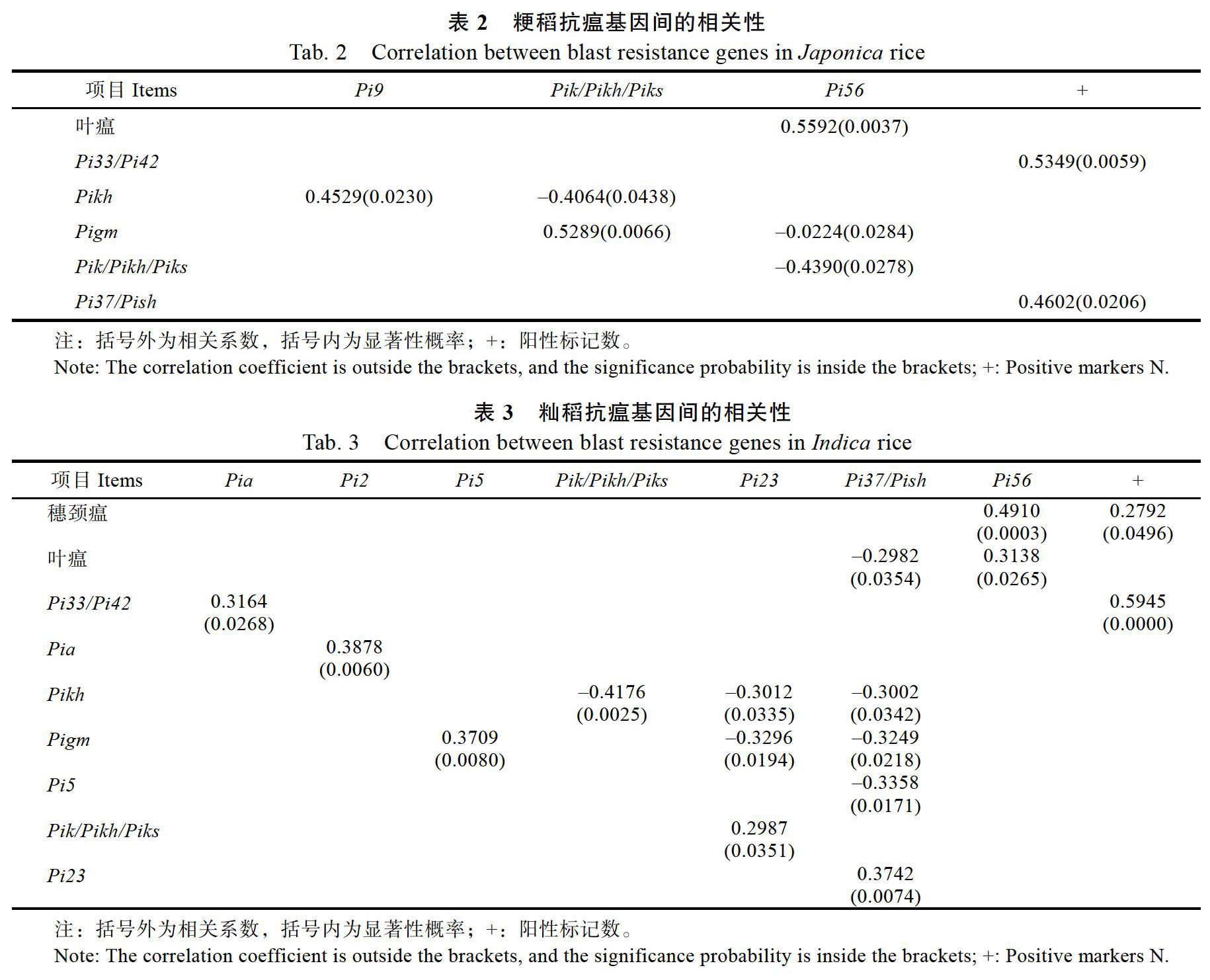
对28份粳稻和54份籼稻的稻瘟病抗性表现和11个抗稻瘟病基因位点之间的相关性分析显示,粳稻的抗穗颈瘟与抗叶瘟之间的相关系数微小,抗叶瘟与Pi56、Pi33/Pi42与阳性标记数、Pigm与Pik/Pikh/Piks之间的相关性呈极显著水平,而Pikh与Pik/Pikh/Piks、Pi56与Pigm、Pi56与Pik/Pikh/Piks之间的相关性呈负显著(表2)。由表3可看出,籼稻的抗穗颈瘟与抗叶瘟之间的相关系数为0.0883,但未达显著水平(P=0.05422),抗穗颈瘟与Pi56、Pi33/Pi42与阳性标记数、Pia与Pi2、Pigm与Pi5、Pi23与Pi37/Pish之间的相关性达到了极显著水平,Pikh与Pik/Pikh/Piks之间达到了极显著负相关。另外,Pi37/Pish与抗叶瘟、Pi23与Pikh、Pi23与Pikh和Pigm、Pi37/Pish与Pikh、Pi37/Pish与Pigm、Pi37/Pish与Pi5之间达到了显著负相关。
2.2 抗性基因间的遗传聚类分析
对28份粳稻材料的穗颈瘟指数、叶瘟指数和11个抗稻瘟位点的聚类分析显示,在遗传距离为0.75时划分为3类;在遗传距离为0.75~1.0之间时,有6份材料聚为一类,有3份材料未聚类(图1)。在54份籼稻材料的聚类分析中,在遗传距离为0.50时可划分为5大类,在遗传距离为0.75时划分为2类,有3份材料的抗性遗传距离大于0.75,4份材料未聚类(图2)。
2.3 分子标记检测结果
从表4可知,28份粳稻中,有1份材料携带其中11个抗性位点中的7个抗性基因位点,有5份材料携带6个抗性基因,有7份材料携带5个抗性基因,有12份材料携带4个抗性基因,有3份材料携带3个抗性基因。抗性频率数最高的Pia有22份,其次是Pi5有21份,Pikh和Pi9,频率数均为18份,Pigm和Pi37/pish在这批粳稻材料中的抗性频率数为0。除了RM24022和RM249分别有3和4个等位基因之外,其他标记在这批粳稻骨干亲本中都只有2个等位基因。
54份籼稻抗稻瘟病基因检测结果显示(表5),有8份材料携带其中的7个抗性基因,有10份材料携带6个抗性基因,有13份材料携带5个抗性基因,有17份材料携带4个抗性基因,有4份材料携带3个抗性基因,有2份材料携带2个抗性基因,抗性频率数最高的Pi9为53份,其次Pik/Pikh/Piks为45份,Pi56在这批籼稻材料中的抗性频率数为0。RM72有5个等位基因,Pigm有4个等位基因,Pi2、RM249、RM24022有3个等位基因之外,其他标记只有2个等位基因。
2.4 抗性基因的频率分布
抗性基因在这2类材料中的频率分布差异较大,在28份骨干粳稻中,Pia的频率分布最大为0.786,其次为Pi5、Pikh、Pi9、Pi33/Pi42、Pi23,抗性频率分别为0.750、0.643、0.643、0.607、0.571,Pigm、Pi37/pish的抗性频率均为0。在54份骨干籼稻中,Pi9的抗性频率分布最大为0.981,其次为Pik/Pikh/Piks、Pigm、Pi37/Pish、Pikh,抗性频率分别为0.833、0.556、0.537、0.519,pi56的抗性频率为0(图3)。
3 讨论
3.1 抗性强弱需要主效基因和正向互作
虽然大部分的研究认为,聚合多个抗性基因比单个抗性基因的抗性增强[8, 28-29],但也不是绝对的,由于基因间的互作等原因[7],聚合材料的抗性不完全等同于單个基因抗性的简单叠加[13],甚至有些单个抗病基因的抗病能力在几年后会逐步减弱,甚至丧失[30-31],就算持续抗性20多年的谷梅4号、地谷等王牌品种也是含有多个主效抗病基因,才能持续多年达到高抗的水平[32]。本研究中BP39携带5个抗性位点,穗颈瘟是4级,而1级抗性的武育粳33、鹅湖香粳2和奉新晚粳却只带有4个抗性位点;携带5个抗性位点的R927、GB6,穗颈瘟也是4级,携带6个抗性位点的浙江长粒粳的抗性是3级,而携带3个抗性位点的麻壳糯、黄壳糯的抗性是1级,携带4个抗性位点的马坝油毡抗性是0.5级。抗性位点之间还存在负相关性,与王军等[33]的研究结果一致,说明抗性级别和主效抗性基因及抗性位点之间的互作相关。当然,也有一些抗性位点表现出了正加性效应,使得像粳226、粤油丝苗、粤禾丝苗、粤晶丝苗2、粤王丝苗等带有7个抗性位点表现出了1级强抗。
3.2 抗性基因的应用与当地的病原生理小种相关
水稻不同品种对稻瘟病的抗性具有致病型专化性,抗性基因所发挥的作用也不近相同[34],有些抗性基因(如Pigm)对水稻的产量性状还存在一定影响[12],不同稻区、不同年份的稻瘟病菌往往表现出很大的抗性差异。江苏粳稻中,Pi-ta和Pi-b基因可以大大提高粳稻穗颈瘟的抗性[26],Pi-b的分布频率要高于Pi-ta[9],而Pi-ta的效果又比Pi-b好[28];浙江粳稻中,Pib、Pi25、Pi40和Pi41基因的分布最广、频率最高[35],其次是Pia,Pi1/Pi3/Pi5和Pit抗性基因的频率较小[36];云南抗病材料中,Pi9和Pi20基因的抗谱最宽,其次是Pita和Pizt[17],而Pi-z对云南育种抗性改良的效果最好[37];安徽抗稻瘟病育种中,Pi9基因具有最高的应用价值[38],整个黄淮区粳稻抗稻瘟病育种中,Pi-ta、Pi-b、Pi54和Pikm的抗性基因应用广泛[39];新疆广谱抗性的基因是Pi2、Pigm、Pil、Pikh/Pi54和Pi63,生理小种特异性抗性的基因是Pia、Pid2和Pid3/Pi25[40];河南稻瘟病育种中,Pi9和Piz-t基因的利用频率最广[31];宁夏抗稻瘟病基因Pikh的分布频率最高;黑龙江抗性育种中,单个Pi2/Pizt、Pita和Pi5的贡献较大,而Pita和Pi5 聚合时抗性改良最大[41];广东省常规水稻抗稻瘟病育种的两大抗源是28占和青六矮1号[42];江西早稻主栽杂交稻组合中抗性基因频率最高的是Pi-z和Pi-d3[3]。本研究中,江西井冈山粳稻抗性频率最高的是Pia,籼稻抗性频率最高的是Pi9,籼粳抗性共同相关的位点是Pi56,且穗瘟与叶瘟的抗性反应明显不一致。说明抗穗颈瘟和抗叶瘟能力可能与发病时期的感应不同,或者抗性机制可能不同,不同类型的材料抗病基因之间存在着非常复杂的互作关系,这与前人的结论是一致的[37, 43-44]。抗病基因数量在这批粳稻中对穗颈瘟和叶瘟抗性无规律可循,在这批籼稻中一般抗病基因数量越多,抗性较强,说明粳稻抗性基因之间的互作关系要远复杂于籼稻,在抗性育种中对寻求主效抗性基因和正效应基因之间的互作至关重要。江西粳稻抗性育种需要Pia的主效基因及pi23等基因的互作,籼稻抗性育种需要Pi9的主效基因及Pi2和Pik/Pikh/Piks等的互作。
3.3 水稻抗稻瘟病的新基因与新思路
水稻基因组中带NBS-LRR抗性蛋白家族基因有500多个[6, 17],现在克隆的基因只占小部分。王军等[2]2014年从江苏省粳稻品种中发现了Pi-kh位点的等位基因;李彬等[4]在第11号染色体长臂末端RM4112和RM2064之间发现了抗稻瘟病新基因Pi-ja;朱亚军等[18]将发现的2个新的抗性位点命名为Pi-7-1(t)和Pi-7-2(t);Tsuyoshi等[19]在第6号染色体上Pid3位点发现一个新的复等位基因Pid3-I1;肖湘谊等[45]在湖南地方品种湘资3150中于第11号染色体标记RM224和RM5926间鉴定了广谱、持久抗稻瘟病新基因Pi47;周镕等[27]在云南地方具广谱持久抗稻瘟的粳稻品种子预44中共检测出13个抗稻瘟QTLs,其中卓晓轩等[46]在其第4号染色体长臂上鉴定了新的单显性抗稻瘟病基因Pi-zy4(t),李金璐等[47]也在其中鉴定了1个新的抗稻瘟病候选基因,说明水稻抗稻瘟病基因的挖掘还大有可为。本研究中,第5号染色体上与pi23基因连锁的RM249标记检测到4个等位基因,其中28份粳稻中特异等位基因数有9个,占比10.98%,54份籼稻中特异等位基因数有25个,占比46.30%,且特异等位基因对应的抗性表型不一,有高抗也有高感,说明在第5号染色体上pi23基因附近可能存在新的抗稻瘟基因,这个基因在籼稻中的频率更高。连锁标记在一批材料中扩增的等位基因数过多,
说明材料的遗传多样性丰富,但可能影响基因型的统计,我们对于非抗性的等位基因都计为非抗,后期酌情更换标记,或开发新的更好用的标记。
从抗性育种角度来选择亲本,类群内材料的基因重组有利于水平抗性的加强,内群外材料的基因重组有利于垂直抗性(广谱抗性)的加强,因此抗性材料的聚类研究为通过基因互补创建广谱、高抗材料亲本的选择指明了方向。另外,Sánchez-Sanuy等[48]报道,在病原菌侵染过程中,铁在病原菌进入的M. oryzae附着胞附近积累,从而抵御病菌的入侵,提高抗性。日本用Fluidigm SNP基因分型平台,对水稻品种特异性抗瘟基因建立了一个基于快速DNA分型的完善的SNP标记系统[20],以及利用报道的24个稻瘟病无毒基因调控水稻的抗性[1],这些都为提高水稻稻瘟病的抗性与分子改良开辟了新思路。
4 结论
本研究中,籼粳稻的抗穗颈瘟和抗叶瘟之间的相关性都不显著,可能是发病时期的感应不同,也说明穗颈瘟和叶瘟的抗性机制可能不同;粳稻的抗叶瘟与RM24022(Pi56连锁)相关系数达到极显著,籼稻的抗叶瘟与RM24022相关系数达到显著,且籼稻抗穗颈瘟与RM24022相关系数达到了极显著,与阳性标记数的相关系数达到了显著,说明这个抗瘟标记位点在籼粳群体材料中表现了较强的群体抗性。10份强抗(抗穗颈瘟和叶瘟)粳稻均带有Pia基因,都不带Pigm基因和Pi37/Pish;5份抗性较弱的粳稻均带有Pi33、Pi42基因,不带Pi2、Pigm、Pik/Pikh和Pi37/Pish基因,说明粳稻中抗性最强的基因是Pia,Pia基因可能是粳稻强抗性所必须的,但还需要pi23等基因的相互作用。相同类型的材料带有相似的抗性标记,如广东优质稻大都带有Pi9、Pi37/Pish和Pik/Pikh/Piks,而不带Pi56;粳型广亲和恢复系大都带有Pi9,都不带Pi2、Pikh、Pi23和Pi56,江西引进这些粳稻材料时需要注意Pi2基因的引入;赣州3份原生地大禾谷资源的抗性基因主要也是pi9;说明籼粳抗性强的共同标记是Pi9和Pikh;籼稻第5号染色体Pi23的附近可能有新的抗性基因。28份粳稻中,带抗性基因位点多的材料,抗性基本较强(BP39例外),也有部分带抗性基因位点少的材料,抗性也较强,说明要注重主效抗性基因位点的应用;云南陆稻粳稻的抗性都比较强。54份籼稻中,带抗性基因位点少的材料,抗性基本较弱,带抗性基因位点多的材料,抗性基本较强。但也有例外,浙江长粒粳籼带11个抗性位点中的7个抗性基因位点,抗性却很弱。所有31份穗颈瘟抗性较强的都带有Pi9和Pik/Pikh/Piks,带有Pigm的只有13份,带Pi2的有16份。但4级抗性中也大部份带有Pi9,却都不带Pi2和Pikh,说明籼稻的群体强抗性中,Pi9可能是必需要的基因,但还需要像Pi2、Pik/Pikh/ Piks等基因的协同作用。
参考文献
[1] 曹 妮, 陈 渊, 季芝娟, 等. 水稻抗稻瘟病分子机制研究进展[J]. 中国水稻科学, 2019, 33(6): 489-498.
[2] 王 军, 杨 杰, 朱金燕, 等. 稻瘟病抗病基因Pi-kh功能标記的开发及江苏粳稻品种中Pi-kh的变异[J]. 中国水稻科学, 2014, 28(2): 141-147.
[3] 兰 波, 杨迎青, 陈 建, 等. 江西水稻主栽品种的稻瘟病抗性基因分子标记检测与分析[J]. 分子植物育种, 2019, 17(8): 2559-2567.
[4] 李 彬, 邓元宝, 颜学海, 等. 一个粳稻来源抗稻瘟病基因的鉴定、遗传分析和基因定位[J]. 作物学报, 2014, 40(1): 54-62.
[5] Alok Das, Soubam D, Singh P K, et al. A novel blast resis-tance gene, Pi54rh cloned from wild species of rice, Oryzar hizomatis confers broad spectrum resistance to Magnaporthe oryzae[J]. Functional and Integrative Genomics, 2012, 12(2): 215-228.
[6] Li W T, Chern M S, Yin J J, et al. Recent advances in broad-spectrum resistance to the rice blast disease[J]. Current Opinion in Plant Biology, 2019, 50: 114-120.
[7] 杨德卫, 王 莫, 韩利波, 等. 水稻稻瘟病抗性基因的克隆、育种利用及稻瘟菌无毒基因研究进展[J]. 植物学报, 2019, 54(2): 265-276.
[8] Hittalmani S,Parco A,Mew T V, et al. Fine mapping and DNA marker-assisted pyramiding of the three major genes for blast resistance in rice[J]. Theoretical and Applied Ge-netics, 2000, 100(7): 1121-1128.
[9] 何 重, 陈 涛, 张亚东, 等. 江苏部分粳稻品种和品系中稻瘟病抗性基因 Pi-ta和Pi-b的基因型分析[J]. 江苏农业学报, 2014, 30(5): 921-927.
[10] Deng Y, Zhu X, Shen Y, et al. Genetic characterization and fine mapping of the blast resistance locus Pigm(t) tightly linked to Pi2 and Pi9 in a broad-spectrum resistant Chinese variety[J]. Theoretical and Applied Genetics, 2006, 113(4): 705-713.
[11] 张礼霞, 王林友, 范宏环, 等. 利用Pigm基因改良粳稻保持系的稻瘟病抗性研究[J]. 核农学报, 2017, 31(3): 424-431.
[12] 曾生元, 李 闯, 杜灿灿, 等. Pigm特异性选择标记的开发及其在粳稻穗颈瘟抗性育种中的利用[J]. 中国水稻科学, 2018, 32(5): 453-461.
[13] 张柱坚, 陈子强, 顾建强, 等. 稻瘟病抗性基因Pi-d2、Pi-d3和Pigm不同敲除突变体的抗性评价[J]. 福建农业学报, 2018, 33(12): 1231-1236.
[14] 王芳权, 陈智慧, 许 扬, 等. 水稻广谱抗稻瘟病基因PigmR功能标记的开发及应用[J]. 中国农业科学, 2019, 52(6): 955-967.
[15] 陈 涛, 孙旭超, 张善磊, 等. 稻瘟病广谱抗性基因Pigm特异性分子标记的开发和应用[J]. 中国水稻科学, 2020, 34(1): 28-36.
[16] 马继琼, 杨 奕, 孙一丁, 等. 云南地方水稻品种抗稻瘟病基因Pita功能区段序列变异分析[J]. 中国水稻科学, 2016, 30(3): 265-272.
[17] 邹 茜, 董丽英, 袁平荣, 等. 广谱持久抗病基因Pi40在云南高原粳稻的应用研究[J]. 西南农业学报, 2016, 29(7): 1493-1498.
[18] 朱亚军, 孙 强, 王金明, 等. 粳稻品种吉粳809的稻瘟病抗性基因分析[J]. 作物学报, 2016, 42(11): 1638-1646.
[19] Inukai T, Nagashima S, Kato M. Pid3-I1 is a race-specific partial-resistance allele at the Pid3 blast resistance locus in rice[J]. Theoretical and Applied Genetics, 2019, 132(2): 395-404.
[20] Kitazawa N, Shomura A, Mizubayashi T, et al. Rapid DNA-genotyping system targeting ten loci for resistance to blast disease in rice[J]. Breeding Science, 2019, 69(1): 68-83.
[21] 刘畅媛, 孙一丁, 马继琼, 等. 中国部分水稻品种Bsr-d1启动子稻瘟病持久抗性位点序列分析[J]. 分子植物育種, 2020, 18(13): 4331-4337.
[22] 马继琼, 孙一丁, 杨 奕, 等. 稻瘟病抗性基因Pita在中日韩粳稻中的分布[J]. 分子植物育种, 2020, 18(2): 459-465.
[23] 李 书, 李 权, 樊琳琳, 等. 高原粳稻子预44中三个稻瘟病抗性基因的假基因化分子标记鉴定[J]. 分子植物育种, 2014, 12(2): 219-225.
[24] 余丽琴, 尹建华. 江西省晚稻生产“籼改粳”的战略思考[J]. 江西农业学报, 2016, 28(11): 20-23.
[25] 何秀英, 王 玲, 吴伟怀, 等. 水稻稻瘟病抗性基因的定位、克隆及育种应用研究进展[J]. 中国农学通报, 2014, 30(6): 1-12.
[26] 王 军, 杨 杰, 杨金欢, 等. Pi-ta、Pi-b基因在江苏粳稻穗颈瘟抗性育种中的价值分析[J]. 华北农学报, 2012, 27(6): 141-145.
[27] 周 镕, 王 波, 杨 睿, 等. 粳稻子预44中稻瘟病数量抗性位点分析[J]. 植物学报, 2015, 50(6): 691-698.
[28] 陈 涛, 张亚东, 朱 镇, 等. Pi-b和Pi-ta基因在江苏省粳稻中的分布以及与穗颈瘟抗性的关系[J]. 江苏农业学报, 2016, 32(1): 1-8.
[29] Kanyange L, Kamau J, Ombori O, et al. Genotyping for blast (Pyric?Laria oryzae) resistance genes in F2 pop?Lation of supa aromatic rice (Oryza sativa L.)[J/OL]. International Journal of Genomics, 2019: 5246820 (2019-11-15) [2020-05-08]. https://www.hindawi.com/journals/ijg/2019/ 5246820/.
[30] 宛柏杰, 劉 凯, 赵绍路, 等. 水稻抗稻瘟病基因Pi-ta、Pi-b、Pigm和Pi54在骨干亲本中的分布以及对穗颈瘟抗性的作用[J]. 西南农业学报, 2020, 33(1): 1-6.
[31] 王生轩, 李俊周, 谢 瑛, 等. 河南粳稻抗稻瘟病基因Pi9、Pita和Piz-t的分子检测[J]. 分子植物育种, 2017, 15(3): 951-955.
[32] 李 刚, 袁彩勇, 曹奎荣, 等. 544份水稻种质稻瘟病抗性鉴定及抗性基因的分布研究[J]. 中国农业大学学报, 2018, 23(5): 22-28.
[33] 王 军, 宫丹妮, 杨 杰, 等. 江苏省粳稻品种抗稻瘟病基因型与穗颈瘟抗性分析[J]. 江苏农业学报, 2016, 32(2): 250-256.
[34] 岂长燕, 许兴涛, 马 建, 等. 抗稻瘟病基因Pib、Pita、Pi5、Pi25和Pi54在我国水稻微核心种质中的分布[J]. 植物遗传资源学报, 2019, 20(5): 1240-1246, 1254.
[35] 翟荣荣, 叶胜海, 朱国富, 等. 浙江省12个常规晚粳稻品种抗稻瘟病基因的分子检测[J]. 分子植物育种, 2020, 18(11): 3626-3633.
[36] 何海燕, 柴荣耀, 邱海萍, 等. 五个抗稻瘟病基因在浙江省水稻品种中的分布和抗性评价[J]. 浙江农业学报, 2019, 31(6): 922-929.
[37] 张学堂, 廖新华, 朱振华, 等. Pi-z、Pi-ta2等基因在云南粳稻稻瘟病抗性育种中的应用[J]. 中国农业科技导报, 2010, 12(1): 100-105.
[38] 陈晴晴, 夏加发, 张海珊, 等. 106份水稻材料稻瘟病抗性鉴定及Pi9基因的分子检测[J]. 安徽农业科学, 2020, 48(7): 154-156.
[39] 陈 峰, 徐建第, 姜明松, 等. 黄淮区粳稻抗稻瘟病基因Pi-ta、Pi-b、Pi54、Pikm的分子检测[J]. 生物技术进展, 2018, 8(1): 46-54.
[40] 张燕红, 贾春平, 文孝荣, 等. 新疆水稻主要育成品种13个稻瘟病的抗性基因分布[J]. 新疆农业科学, 2017, 54(9): 1595-1605.
[41] 相亚超, 王丽丽, 徐 凡, 等. 抗稻瘟病基因在黑龙江水稻资源中的分布[J]. 分子植物育种, 2018, 16(23): 7705-7717.
[42] 钟春燕, 孟 醒, 王茂辉, 等. 广东省常规水稻品种稻瘟病抗性研究与分析[J]. 广东农业科学, 2020, 47(2): 102-109.
[43] 董丽英, 李国生, 赵秀兰, 等. 云南粳稻品种(系)对稻瘟病的抗性鉴定与评价[J]. 西南农业学报, 2018, 31(12): 2458-2465.
[44] 余 玲, 戴正元, 吴云雨, 等. 利用分子标记辅助选择提高江苏粳稻稻瘟病抗性[J]. 扬州大学学报(农业与生命科学版), 2014, 35(4): 57-62.
[45] 肖湘谊, 史学涛, 盛浩闻, 等. 水稻抗稻瘟病基因Pi47的精细定位和候选基因分析[J]. 作物学报, 2018, 44(7): 977-987.
[46] 卓晓轩, 樊琳琳, 安星宇, 等. 云南地方品种子预44中一个新的抗稻瘟病基因的定位[J]. 中国水稻科学, 2019, 33(1): 12-19.
[47] 李金璐, 张 慧, 焦泽宇, 等. 水稻子预44和江南香糯基因组比较鉴定稻瘟病抗性相关基因[J]. 中国水稻科学, 2020, 34(1): 8-16.
[48] Sánchez-Sanuy F, Peris-Peris C, Tomiyama S, et al. Osa-miR7695 enhances transcriptional priming in defense responses against the rice blast fungus[J]. BMC Plant Biolo-gy, 2019, 19(1): 563.
责任编辑:黄东杰
