荔枝LcMYB1异源表达促进矮牵牛和番茄花色苷的积累
2021-07-20杜丽娜陈春帆苏睿谭春艳赖彪
杜丽娜 陈春帆 苏睿 谭春艳 赖彪
摘 要:为分析荔枝LcMYB1的功能,以白色‘W115矮牵牛和‘Micro-Tom番茄为材料,利用农杆菌介导法将LcMYB1在矮牵牛和番茄中异源表达,观测转基因植株表型。结果表明:与‘W115相比,转基因矮牵牛的叶片和花瓣中积累了花色苷,且矮牵牛花色苷生物合成结构基因PhCHS和PhDFR、调控基因PhAN1的表达显著上调;与野生型‘Micro-Tom番茄相比,转基因番茄的叶片和花药中积累了花色苷,且相应组织花色苷生物合成结构基因SlDFR、调控基因SlAN1和SlJAF13的表达显著上调,虽然果实中没有积累花色苷,但SlAN1和SlJAF13表达也显著上调。LcMYB1在矮牵牛和番茄中异源表达时通过上调花色苷生物合成关键结构基因和调控基因bHLH的表达诱导花色苷积累。因此,荔枝LcMYB1是花色苷生物合成中的关键转录因子,具备异源转化利用的潜力。
关键词:LcMYB1;矮牵牛;番茄;花色苷
中图分类号:S667.1 文献标识码:A
Abstract: In order to better understand the function of LcMYB1 and provide theoretical support for further utilization, LcMYB1 was transformed into both petunia and tomato. White flower ‘W115 petunia and ‘Micro-Tom tomato were used as materials in LcMYB1 ectopic expressed assays. Anthocyanin contents and related gene expressions were ana-lyzed in transgenic plants. Ectopic expression of LcMYB1 in ‘W115 resulted in anthocyanin production in vegetative and floral tissues such as leaves and petals, probably by transcriptional activation of anthocyanin biosynthetic genes such as PhCHS and PhDFR and endogenous anthocyanin regulatory gene PhAN1. However, the transgenic tomato only produced anthocyanin in anthers and leaves but not in tomato fruits and petals. The results suggested that LcMYB1 could enhance anthocyanin production in vegetative and floral tissues of both petunia and tomato by activating anthocyanin biosynthesis and regulatory genes. LcMYB1 is an important transcriptional regulatory factor in anthocyanin biosynthesis of plants and is potentially used in ectopic transformation.
Keywords: LcMYB1; petunia; tomato; anthocyanin
DOI: 10.3969/j.issn.1000-2561.2021.05.013
色澤是花卉和果蔬的重要品质组成之一。花色苷是植物组织或器官呈现出粉色、红色、紫色甚至黑色的重要次生代谢成分之一。花色苷除了能给蔬菜、花卉、水果带来鲜艳的颜色,还具有对生物和非生物胁迫给植物带来伤害的保护作用,比如:紫外线照射、冷胁迫、干旱胁迫等[1-3]。大量研究表明,花色苷合成代谢的调控主要在转录水平上[4]。转录因子R2R3-MYB、bHLH(basic Hlix-Loop-Hlix)和WD40相互作用形成MBW蛋白复合体,通过调控花色苷合成代谢的结构基因表达来调控植物花色苷的生物合成[5]。
苹果(Malus domestica)中有2个MYB转录因子调控果实花色苷的生物合成,其中MdMYB1主要调控光依赖的果皮着色[6],而MdMYB10主要调控果肉的花色苷积累[7]。此外,在梨(Pyrus pyrifolia)[8]、山竹(Garcinia mangostana)[9]、杨梅(Myrica rubra)[10]等水果中也克隆到了调控果实花色苷生物合成的关键MYB类转录因子。这些花色苷生物合成的关键调控基因的发现为果实的遗传改良提供重要的基因资源。
荔枝(Litchi chinensis)是我国南方重要的热带亚热带果树,种质资源丰富,果实色泽类型多样。荔枝果皮的红色是花色苷积累的结果,前期利用同源序列方法克隆了荔枝LcMYB1,并在烟草中超表达LcMYB1诱导了叶片和花瓣花色苷的积累[11]。本文将该基因在‘W115矮牵牛和‘Micro-tom番茄中超表达,进一步研究LcMYB1的功能,为该基因作为基因工程育种的候选基因提供理论依据。
1 材料与方法
1.1 材料
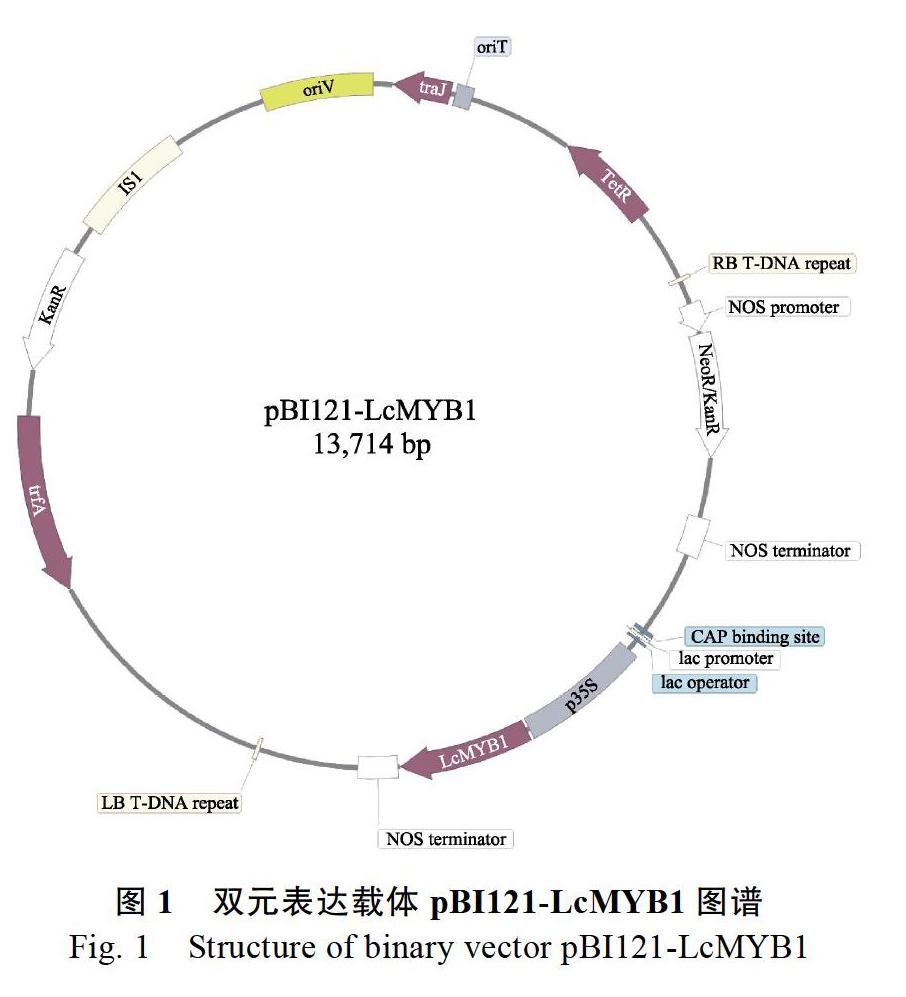
实验所用的‘Micro-Tom番茄、‘W115矮牵牛(MYB转录因子PhAN2和PhAN4双突变体)和根癌农杆菌菌株AGL0由阿姆斯特丹大学Francesca M Quattrocchio和Ronald Koes研究小组馈赠。pBI121-LcMYB1(图1)质粒由本实验室保存。
1.2 ‘W115矮牵牛的遗传转化
取温室生长状态良好的‘W115植株叶片,表面消毒(70%酒精清洗30 s,10%次氯酸清洗10 min,然后用无菌水清洗4~5次),用无菌手术刀在超净工作台内将准备好的叶片切成1 cm× 1 cm的小方块,放入准备好的OD600为0.1的农杆菌中侵染10 min。将侵染后的外植体取出,用灭菌滤纸吸收多余的农杆菌后放入共培养培养基(1/2 MS+2.0 mg/L 6-BA+0.1 mg/L NAA+1.0 mg/L玉米素+2.0%蔗糖+1%葡萄糖)中。于25 ℃共培养2 d后将外植体转移到筛选培养基(1/2 MS+ 2.0 mg/L 6-BA+0.1 mg/L NAA+1.0 mg/L玉米素+ 2%蔗糖+1%葡萄糖+250 mg/L头孢霉素+100 mg/L卡那霉素)中,待出芽后将芽切下转移到生根培养基(1/2 MS+2%蔗糖+1%葡萄糖+250 mg/L头孢霉素+100 mg/L卡那霉素)中培养生根。
1.3 ‘Micro-Tom番茄的遗传转化
取适量的‘Micro-Tom种子进行表面消毒(70%酒精清洗2 min,10%次氯酸清洗10 min,然后用无菌水清洗4~5次),将消毒的种子播种于MS培养基中,待种子发芽长出叶子备用(大约10 d左右)。将一片子叶切成2段后转移到准备好的农杆菌中侵染10 min,将侵染后的外植体取出,用灭菌滤纸吸收多余的农杆菌后放入共培养培养基(1/2 MS+0.1 mg/L IAA+0.4 mg/L玉米素+2%葡萄糖+1.0 mmol/L MES)中。于25 ℃共培養2 d后将外植体转移到筛选培养基(1/2 MS+0.1 mg/L IAA+0.4 mg/L玉米素+2%葡萄糖+1.0 mmol/L MES+250 mg/L头孢霉素+100 mg/L卡那霉素)中,待出芽后将芽切下转移到生根培养基(1/2 MS+2%葡萄糖+1.0 mmol/L MES+250 mg/L头孢霉素+100 mg/L卡那霉素)中培养生根。
1.4 花色苷含量测定
参照Wrolstad等[12]的方法,采用pH示差法测定矮牵牛和番茄各组织器官的花色苷含量,具体步骤为:分别取各组织0.1 g样品于3 mL浸提液(甲醇∶水∶浓盐酸=85∶12∶3)中,室温避光充分浸提(约5~6 h)。Buffer 1:0.2 mol/L KCl-0.2 mol/L HCl(25∶67),pH=1;Buffer 2:1 mol/L NaAc-0.4 mol/L HCl(100∶150),pH 5。取0.5 mL浸提液分别加入2支试管中,然后分别加入2 mL的Buffer 1和Buffer 2,用紫外分光光度计分别测定530 nm吸光值。
根据经验公式计算结果,花色苷含量(mg/g)= ;其中5为稀释倍数,3为提取液体积(mL),445.2为矢车菊素-3-葡萄糖苷的相对分子质量,29600为矢车菊素-3-葡萄糖苷的摩尔比吸收系数,0.1表示样品质量(g)。
1.5 RNA的提取与cDNA合成
使用Magen公司的Hipure Plant RNA Mini Kit试剂盒(具体方法参照说明书)进行矮牵牛和番茄各组织器官样品RNA的提取。使用DNase I(TaKaRa)消化RNA中残留的DNA(具体方法参照说明书)。RNA的浓度测定和纯度分析:取1 L RNA溶液稀释至100 L,置于紫外核酸蛋白检测仪上分别测定260、280和320 nm吸光值并计算OD260/OD280值。当OD260/OD280值介于1.80~2.00之间时,RNA样品用于下一步试验。总RNA样品浓度按下列公式计算:RNA(ng/L)=OD260× 40 ng/L×100。为进一步评价总RNA的完整性,取2~3 L总RNA进行1.2%(W/V)的琼脂糖凝胶电泳检测。以总RNA作为模板合成第一链cDNA,反转录采用TaKaRa的SuperScript One- Step RT-PCR System试剂盒,引物为Oligo(dT)15(Ta?KaRa)。cDNA的合成参照反转录酶说明书进行。
1.6 荧光定量PCR
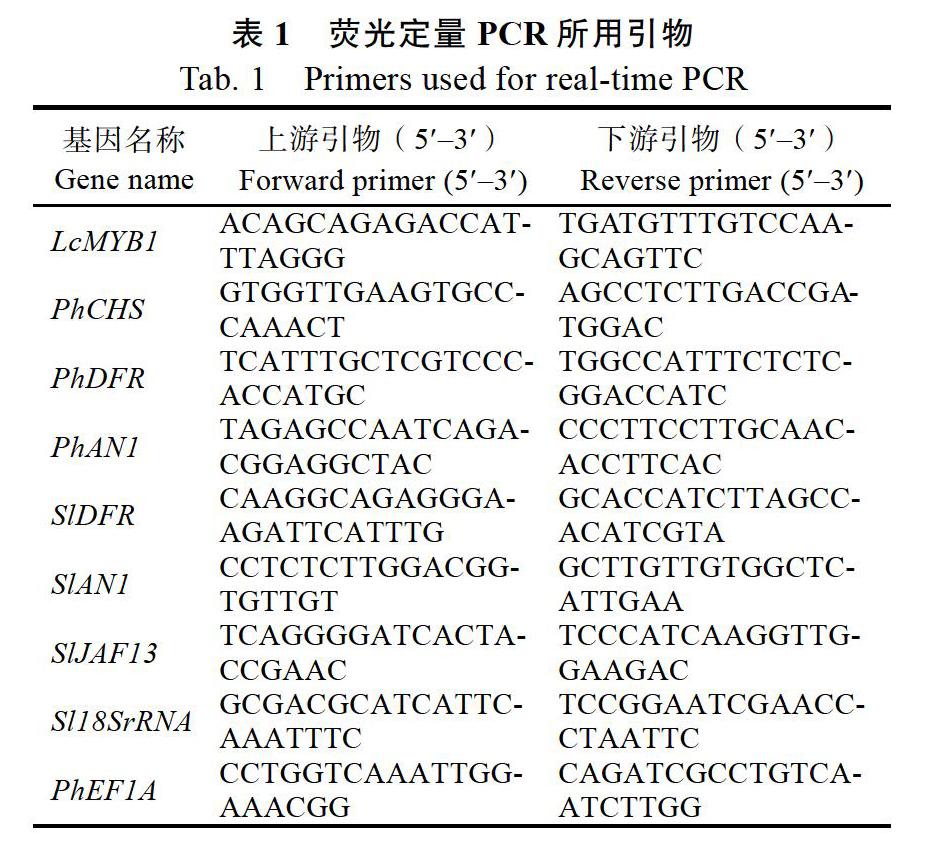
Real-time PCR反应在ABI QuantStudio 3 Real-time PCR System上进行,引物见表1。反应条件如下:95 ℃ 15 s;56 ℃ 15 s;72 ℃ 35 s;40个循环。熔解曲线分析参考Wei等[13]的方法。运用2?ΔΔCT法进行数据处理[14],计算出花色苷生物合成相关基因在不同样品中的表达水平。以上所有实验均设3次重复,每次实验以ddH2O为阴性对照。矮牵牛和番茄的内参基因分别为PhEF1Α和Sl18SrRNA。
2 结果与分析
2.1 转LcMYB1基因矮牵牛表型观测
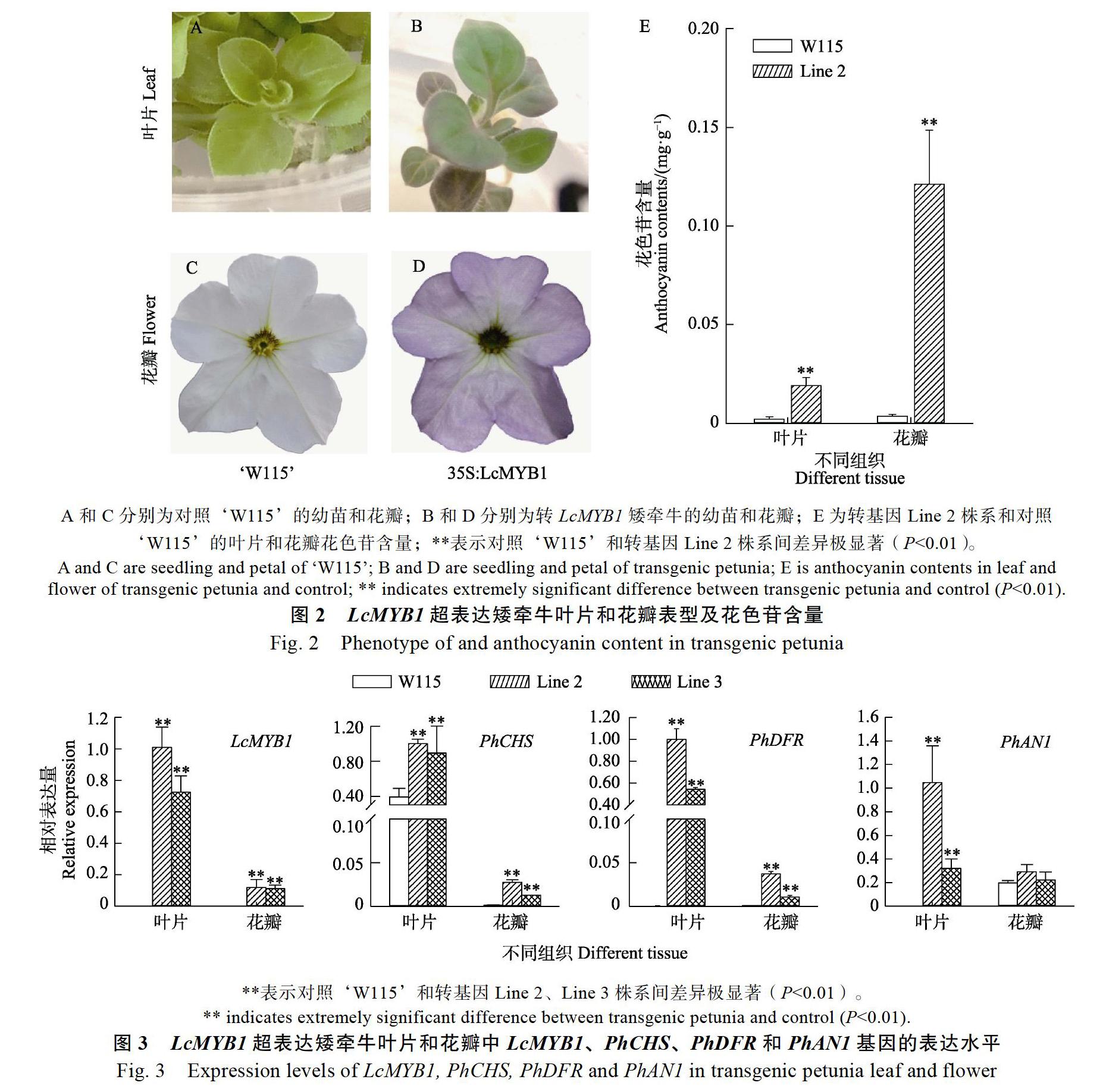
研究中使用的白色‘W115矮牵牛是花色苷合成调控的关键MYB转录因子PhAN2和PhAN4的双突变体,可避免矮牵牛自身MYB转录因子对颜色形成的干扰。将重组质粒pBI121-LcMYB1转化农杆菌AGL0中后,利用农杆菌介导的遗传转化方法,将LcMYB1转入白色‘W115矮牵牛中。与对照相比,转基因矮牵牛的叶片变红,花瓣变为浅紫色(图2A~图2D)。LcMYB1超表达的叶子和花瓣均显著积累花色苷,叶片中花色苷含量为0.02 mg/g,花瓣中花色苷含量为0.12 mg/g(图2E)。说明LcMYB1异源表达可以诱导矮牵牛叶片和花瓣花色苷积累,一定程度上弥补了PhAN2和PhAN4的缺失。
2.2 转LcMYB1基因矮牵牛花色苷生物合成相关基因的表达
为进一步分析LcMYB诱导矮牵牛花色苷的积累机理,利用荧光定量PCR检测了LcMYB1、花色苷生物合成关键结构基因PhCHS、PhDFR和bHLH调控基因PhAN1在对照和转基因植株中的表达情况。结果表明,LcMYB1在对照植株花瓣和叶片中均无表达,在转基因的矮牵牛花瓣和叶片中均表达,且在叶片中的表达量更高,说明LcMYB1已成功转入到‘W115中(图3)。PhCHS在转基因矮牵牛叶片中的表达水平比对照增加1倍,在花瓣中的表达明显被激活。PhDFR在对照的叶片和花瓣中均无表达,而转基因植株中其表达显著被激活,且在葉片中的表达高于花瓣。在对照中,PhAN1在叶片中无表达,在花瓣中有一定的表达,在转基因植株叶片中PhAN1的表达显著被激活,花瓣中PhAN1有一定提高,但不显著。
因此,LcMYB1可能通过上调矮牵牛PhCHS、PhDFR和PhAN1的表达诱导花色苷的积累。
2.3 转LcMYB1基因番茄表型观察
利用农杆菌介导的遗传转化方法,将LcMYB1转入‘Micro-Tom番茄中。与对照相比,转基因番茄的叶片变红,花药也由黄色变成了红色(图4A~图4D)。通过测定分析花色苷含量发现,对照植株除叶片有少量花色苷积累外,果实、花药、花瓣均无花色苷积累,而转基因植株的叶片和花药中积累了花色苷,含量分别为0.24 mg/g和0.10 mg/g,其花瓣和果实中均未检测到花色苷(图4E)。
2.4 转LcMYB1基因番茄花色苷生物合成相关基因的表达
同样,利用荧光定量PCR检测了转基因番茄中LcMYB1、SlDFR和花色苷生物合成关键bHLH调控基因SlAN1和SlJAF13的表达情况。由图5分析发现,LcMYB1在转基因番茄的叶片和花药中表达较高,果实中表达相对较低,而在对照植株的各个器官均无表达,说明LcMYB1已成功转入到‘Micro-Tom番茄中。与在矮牵牛中LcMYB1的表达模式类似,LcMYB1在各个组织中的表达有较大差异。SlDFR和SlAN1的表达模式相似,SlDFR和SlAN1在对照植株的叶片、果实和花药中均有较低或无表达,而在转基因植株的叶片和花药中其表达显著被激活,在果实中的表达量均较低。SlDFR和SlAN1的基因表达水平与番茄的花色苷含量一致。SlJAF13在对照植株叶片、果实和花药中均有一定表达,在转基因植株中该基因在各组织的表达水平均提高了约1倍。
3 讨论
本研究组前期研究发现荔枝LcMYB1在各个组织器官中的表达水平与花色苷含量呈正相关,并通过烟草异源表达确定LcMYB1是调控荔枝中花色苷生物合成的关键转录因子[11]。本研究分别在白色‘W115矮牵牛和‘Micro-Tom番茄中超表达LcMYB1,结果显示,转基因矮牵牛可以在叶片和花瓣中积累花色苷,转基因番茄的叶片和花药积累了大量花色苷。‘W115为矮牵牛花色苷生物合成关键MYB调控基因PhAN2和PhAN4双突变植株,花瓣为白色[15],因此,利用‘W115研究LcMYB1的功能可以避免内源MYB基因的影响。此外,荔枝LcMYB1无论是通过瞬时表达还是稳定遗传转化均可诱导烟草花色苷积累[11],这表明荔枝LcMYB1在烟草、番茄和矮牵牛中均有较强的诱导花色苷生物合成的能力。
MYB转录因子调控花色苷生物合成的机理是MYB转录因子结合到花色苷生物合成途径中的结构基因的启动子上,调节这些结构基因的表达[4]。在植物积累花色苷的过程中,MYB转录因子和结构基因的表达模式和花色苷的积累量一致。在杨梅果实发育过程中,MrF3H、MrF3H、MrDFR1、MrUFGT和MrMYB1的表达水平与杨梅果实的花色苷含量正相关[10]。在苹果不同着色程度的果皮中,MdANS、MdUFGT和MdMYB1的表达水平也与果实的着色程度高度一致[6]。与对照相比,矮牵牛花色苷生物合成关键调控基因PhAN1、结构基因PhCHS和PhDFR在转基因植株中均表达上调,而转基因番茄的叶片、果实和花药中花色苷生物合成的关键调控基因SlAN1、SlJAF13、结构基因SlDFR的表达上调。综合前期研究发现,LcMYB1转录因子可以在不同植物中(包括烟草、矮牵牛和番茄)通过调控花色苷生物合成的结构基因和调控基因来促进或诱导花色苷的积累。
近期研究发现,番茄SlMYB75可以诱导‘Micro-Tom番茄叶片、花药和果实积累大量花色苷,与SlMYB75相比,LcMYB1可以诱导番茄叶片和花药的花色苷生物合成,但并不能诱导番茄果实积累花色苷,推测可能是由于番茄果实中抑制花色苷生物合成的R3-MYB转录因子SlMYBATV的表达与LcMYB1竞争bHLH,从而抑制花色苷的合成,导致无法启动果实中花色苷生物合成的关键结构基因[16-17]。前人将调控玉米花色苷生物合成的Lc(MYB)和C1(bHLH)同时转化到番茄中,无法诱导番茄果实花色苷的合成[17]。而在番茄中同时超表达来自金鱼草的Del(bHLH)和Ros1(MYB),则可以诱导番茄果实合成大量的花色苷[18]。这表明,不同植物来源的MYB转录因子在同一植物异源表达时会出现不同表型。
综上所述,本研究证实了在矮牵牛和番茄中LcMYB1通过上调花色苷生物合成关键结构基因和bHLH调控基因的表达来调控花色苷生物合成的功能,同时也发现LcMYB1转录因子在不同植物上异源表达的表型有一定的差异,不能诱导植物的所有器官积累花色苷,这些结果可为利用基因工程育种在基因选择上提供参考。
参考文献
[1] Ubi B E, Honda C, Bessho H, et al. Expression analysis of anthocyanin biosynthetic genes in apple skin: Effect of UV-B and temperature[J]. Plant Science, 2005, 170(3): 571-578.
[2] Castellarin S D, Pfeiffer A, Sivilotti P, et al. Transcriptional regulation of anthocyanin biosynthesis in ripening fruits of grapevine under seasonal water deficit[J]. Plant, Cell & En-viron, 2007, 30(11): 1381-1399.
[3] Zhang Y, Zheng S, Liu Z, et al. Both HY5 and HYH are necessary regulators for low temperature-induced anthocyanin accumulation in Arabidopsis seedlings[J]. Journal of Plant Physiology, 2011, 168(4): 367-374.
[4] Allan A C, Hellens R P, Laing W A. MYB transcription factors that colour our fruit[J]. Trends in Plant Science, 2008, 13(3): 99-102.
[5] Xu W, Dubos C, Lepiniec L. Transcriptional control of flavonoid biosynthesis by MYB-bHLH-WDR complexes[J]. Trends in Plant Science, 2015, 20(3): 176-185.
[6] Takos A M, Jaffe f W, Jacob S R, et al. Light-induced ex-pression of a MYB gene regulates anthocyanin biosynthesis in red apples[J]. Plant Physiology, 2006, 142(3): 1216-1232.
[7] Espley R V, Hellens R P, Putterill J, et al. Red colouration in apple fruit is due to the activity of the MYB transcription factor, MdMYB10[J]. Plant Journal, 2007, 49(3): 414-427.
[8] Feng S, Wang Y, Yang S, et al. Anthocyanin biosynthesis in pears is regulated by a R2R3-MYB transcription factor Py-MYB10[J]. Planta, 2010, 232(1): 245-255.
[9] Palapol Y, Ketsa S, Lin-Wang K, et al. A MYB transcription factor regulates anthocyanin biosynthesis in mangosteen (Garcinia mangostana L.) fruit during ripening[J]. Planta, 2009, 229(6): 1323-1334.
[10] Niu S S, Xu C J, Zhang W S, et al. Coordinated regulation of anthocyanin biosynthesis in Chinese bayberry (Myrica rubra) fruit by a R2R3 MYB transcription factor[J]. Planta, 2010, 231(4): 887-899.
[11] Lai B, Li X J, Hu B, et al. LcMYB1 is a key determinant of differential anthocyanin accumulation among genotypes, tissues, developmental phases and ABA and light stimuli in Litchi chinensis[J]. PLoS One, 2014, 9(1): e86293.
[12] Wrolstad R E, Culbertson J D, Cornwell C J, et al. Detection of adulteration in blackberry juice concentrates and wines[J]. Journal - Association of Official Analytical Chem-ists, 1982, 65(6): 1417-1423.
[13] Wei Y Z, Hu F C, Hu G B, et al. Differential expression of anthocyanin biosynthetic genes in relation to anthocyanin accumulation in the pericarp of Litchi chinensis Sonn[J]. PLoS One, 2011, 6(4): e19455.
[14] Schmittgen T D, Livak K J. Analyzing real-time PCR data by the comparative C(T) method[J]. Nature Protocols, 2008, 3(6): 1101-1108.
[15] Quattrocchio F, Wing J, Van Der Wouder K, et al. Mo-lecular analysis of the anthocyanin2 gene of petunia and its role in the evolution of flower color[J]. Plant Cell, 1999, 11(8): 1433-1444.
[16] Jian W, Cao H, Yuan S, et al. SlMYB75, an MYB-type
transcription factor, promotes anthocyanin accumulation and enhances volatile aroma production in tomato fruits[J]. Horticulture Research, 2019, 6(1): 22.
[17] Sun C, Deng L, Du M, et al. A transcriptional network pro-motes anthocyanin biosynthesis in tomato flesh[J]. Molecular Plant, 2020, 13(1): 42-58.
[18] Bovy A, De Vos R, Kemper M, et al. High-flavonol toma-toes resulting from the heterologous expression of the maize transcription factor genes LC and C1[J]. Plant Cell, 2002, 14(10): 2509-2526.
[19] Butelli E, Titta L, Giorgio M, et al. Enrichment of tomato fruit with health-promoting anthocyanins by expression of select transcription factors[J]. Nature Biotechnology, 2008, 26(11): 1301-1308.
責任编辑:谢龙莲
