自噬相关蛋白LC3-Ⅱ和p62在不明原因复发性流产患者绒毛组织中的表达和临床意义*
2021-07-19宋鹏书张燕华张奕梅莫梅珍
宋鹏书,张燕华,张奕梅,莫梅珍,何 升,张 莉
广西壮族自治区妇幼保健院妇产科,广西南宁 530003
复发性流产是指3次或3次以上发生在20周之内的妊娠失败,发生率高达3%~5%[1]。复发性流产病因复杂多样,可能与遗传因素、感染因素、免疫学因素、内分泌因素等有关,且约50%的复发性流产为不明原因复发性流产(URSA)[2]。
自噬是一种高度保守的细胞行为,其通过泛素蛋白酶系统维持细胞内稳定,是由双层膜性结构对细胞内成分进行包裹,之后通过与溶酶体结合从而将包裹的内容物进行分解、再利用的过程[3-4]。近年研究发现,自噬反应活性失调对妊娠期并发症的发生存在不可忽视的影响[5]。但有关复发性流产与自噬的相互关系报道较为少见。本研究探讨了自噬相关蛋白LC3-Ⅱ、p62在URSA患绒毛组织中的表达情况,分析URSA结局与绒毛组织发生自噬反应的相关性,旨在研究URSA的发生机制,为进一步寻找新的临床干预方法提供理论依据。
1 资料与方法
1.1一般资料 选取2020年3-7月在本院妇产科行清宫术URSA患者及行人工流产术的正常妊娠者分别作为URSA组和对照组,每组10例。两组患者诊断标准均参照第9版《妇产科学》。URSA组平均年龄(32±6)岁,平均孕周(10±2)周,纳入标准:(1)超声检查子宫发育正常;(2)夫妇染色体核型检查正常;(3)甲状腺功能检查正常;(4) 无血栓形成倾向;(5)自然流产次数≥3次;(6)排除免疫及感染因素影响。对照组平均年龄(30±4)岁,平均孕周(10±2)周,无不良孕产史,正常妊娠次数≥1次。本研究获本院伦理委员会批准,所有研究对象自愿签署知情同意书。
1.2主要试剂 多聚甲醛固定液购自Sigma公司;兔抗人LC3单抗隆抗体购自CST公司;兔抗人p62多克隆抗体购自SAB公司;兔抗人β-actin多克隆抗体购自北京博奥森公司;BCA蛋白定量试剂盒购自武汉博士德生物工程有限公司;山羊抗兔荧光二抗购自Abcam公司;显影液购自Bio-Rad公司。
1.3方法
1.3.1绒毛组织留取 在人工流产术或清宫术术中妊娠组织清出后 3 min内,迅速于无菌状态下挑选绒毛组织,用无菌生理盐水冲洗干净,将其分为三份:两份分别置于已备制好的体积分数为4%的多聚甲醛液中固定 24 h,用于HE染色和免疫组化实验;另一份置于冻存管中,将其迅速放入液氮内,然后移至-80 ℃冰箱保存,用于目的蛋白表达量检测。
1.3.2绒毛组织病理学观察 取固定24 h的绒毛组织,组织块脱水,石蜡包埋,切成厚度4 μm薄片,放入37 ℃温水使切片伸展,贴至载玻片上,依次浸入二甲苯、乙醇脱蜡,蒸馏水冲洗,然后将载玻片放入苏木精溶液中染色,流水冲洗,放入37 ℃温水中进行返蓝,晾干,中性树胶封固保存,电子显微镜观察染色情况。
1.3.3Western blot检测绒毛组织中自噬相关蛋白LC3-Ⅱ、p62的表达 取-80 ℃冰箱保存的绒毛组织,称质量后剪碎放入EP管中,用电动匀浆机研磨碎,加入组织裂解液,放在冰盒上裂解40 min,4 ℃,12 000×g离心20 min,提取组织总蛋白,采用BCA法检测蛋白浓度。采用十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(SDS-PAGE)分离样品蛋白,转至PVDF膜,脱脂奶粉室温封闭30 min,TBST冲洗3次,每次5 min。加入LC3和p62单克隆一抗(1∶500)4 ℃摇晃孵育过夜,TBST冲洗3次,每次5 min,加入二抗室温孵育1 h,然后于显影液中孵育5 min,上机检测。采用Quantity One软件分析目的蛋白相对表达水平。
1.3.4免疫组化实验检测绒毛组织中自噬相关蛋白LC3、p62的表达 取固定24 h的绒毛组织,组织块脱水,石蜡包埋,切成厚度5 μm薄片,放入37 ℃温水表面便于切片伸展容易贴到载玻片上,再依次浸入二甲苯、乙醇脱蜡,高压修复,3%过氧化氢溶液中孵育。用PAP笔在组织切边画圈,然后滴加封闭液。一抗LC3和p62(1∶200)孵育过夜,二抗孵育20 min,二氨基联苯胺(DAB)显色液显色5 min。苏木精染色1 min,分化返蓝,梯度乙醇脱水,树胶封片,电子显微镜观察阳性面积大小。
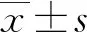
2 结 果
2.1两组绒毛组织的病理学结果 HE染色结果显示,对照组绒毛组织内部细胞形态饱满,细胞核呈梭子型,绒毛组织外部完整,URSA组绒毛组织内部细胞出现皱缩,颜色暗红,细胞之间出现断离,绒毛组织有破损。见图1。
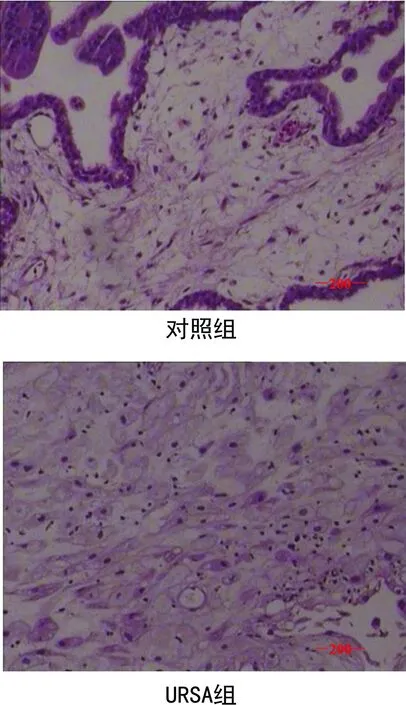
图1 两组流产绒毛组织HE染色结果(×200)
2.2Western blot检测自噬相关蛋白LC3-Ⅱ和p62在流产绒毛组织中的表达 Western blot检测结果显示,与对照组相比,URSA组的绒毛组织中LC3-Ⅱ 的蛋白表达量显著增加;同时,p62蛋白的表达量显著降低,差异有统计学意义(P<0.05)。见图2。

注:A为Western blot检测结果;B为蛋白定量分析;与对照组比较,aP<0.05。
2.3免疫组化实验分析自噬相关蛋白LC3和p62在绒毛组织中的表达 免疫组化实验结果显示,LC3和p62蛋白主要定位于细胞质中。与对照组相比,URSA组LC3蛋白表达量上升, p62蛋白表达量降低,结果与Western blot 检测结果一致。见图3。
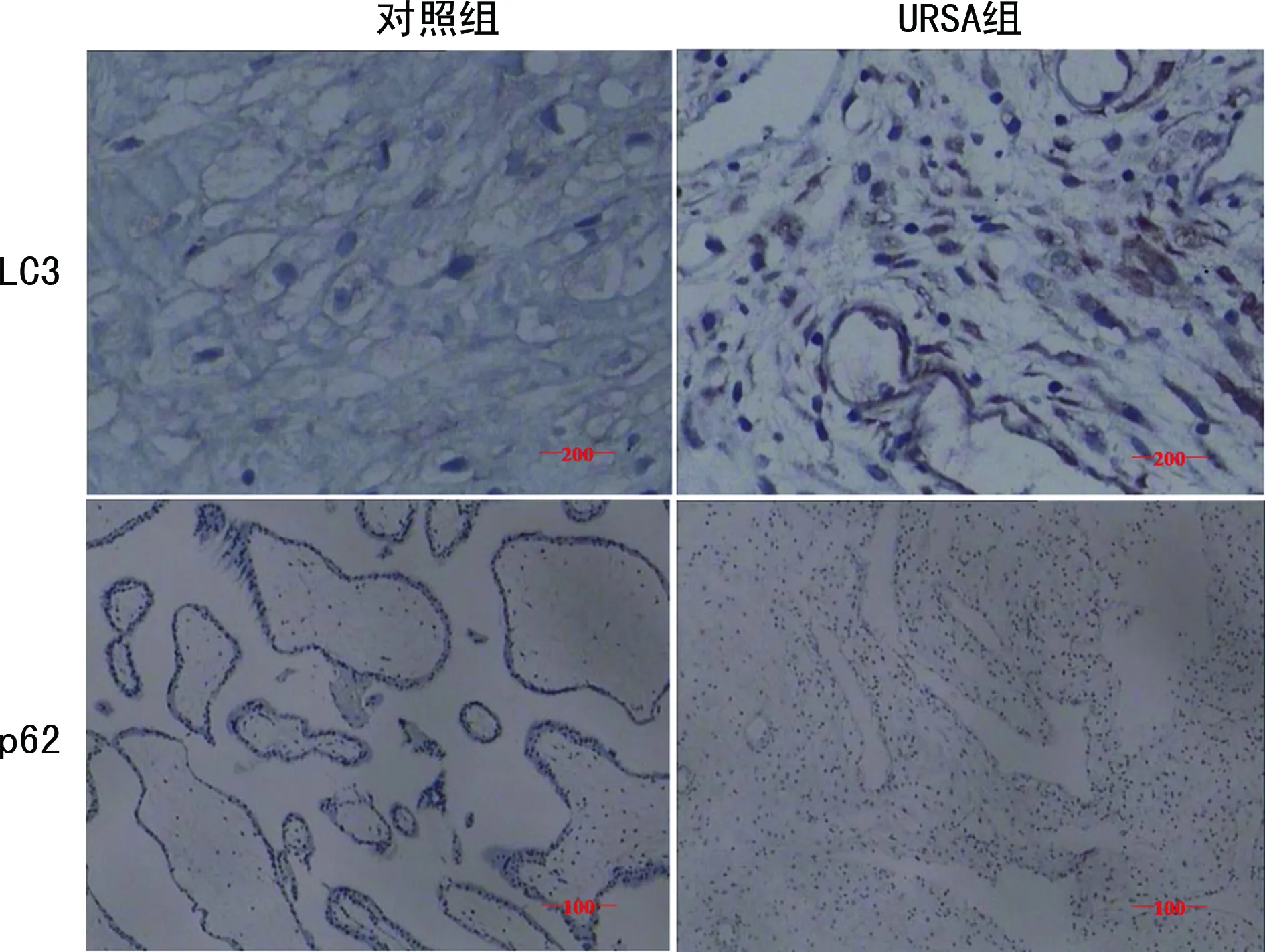
图3 LC3和p62蛋白在两组绒毛组织中的表达情况(×200)
3 讨 论
流产常发生在妊娠早期,其发生率随孕妇年龄的增长而升高。导致流产常见的因素包括免疫功能障碍、染色体异常、内分泌疾病及子宫畸形等。URSA是流产中的一种特殊类型,近年因其发病率呈上升趋势而受到广泛关注[6]。目前,URSA的发病机制尚不明确,且病因复杂,临床治疗尚处于探索阶段,因此成为妇产科领域的研究重点与难点。
卵母细胞受精、胚胎植入前后的发育,以及妊娠期并发症均涉及自噬的诱导,当滋养细胞早期处于缺氧和营养有限的环境下,自噬对其有重要的保护作用,利于其适应早期发育的微环境[7-8]。虽然自噬可以为细胞修复损伤的细胞器提供能量,但当其超过一定限度后,也可能破坏细胞结构,影响能量供应,引发自噬性细胞死亡[9-10]。近年研究显示,与正常妊娠孕妇胎盘绒毛组织相比,流产、子痫前期、胎儿生长受限、妊娠期糖尿病等病理妊娠的孕妇胎盘组织中均出现自噬过程受损[11-12]。此外,KHIDER等[13]研究发现,无论是胎儿早产还是足月分娩,均与胎盘自噬活性降低有关。分娩实际上是一个炎症持续过程,无论早产还是足月分娩,自噬活性的降低都会导致炎症因子清除不足,从而导致促炎细胞因子释放过多,最终引发不良分娩结局[14]。
细胞自噬是多种自噬相关基因和关键蛋白参与的动态过程,可通过这些分子的表达水平来评估自噬活性。在自噬循环过程中,LC3分子经酶切反应后形成胞质型LC3-Ⅰ,后与磷脂酰乙醇胺反应转化为脂质型LC3-Ⅱ,该亚型附着在自噬体上直至与溶酶体融合形成自噬溶酶体[15-16]。LC3-Ⅱ是监测自噬水平稳定可靠的特异性标志物,其含量在一定程度上反映了自噬体的数量。p62也是自噬反应发生过程中的重要衔接分子,其水平变化与自噬强弱呈负相关,常与 LC3共同用于判断自噬反应活性[17]。本研究中,Western blot和免疫组化实验结果均显示,与正常妊娠孕妇相比,URSA患者的绒毛组织中自噬相关蛋白LC3-Ⅱ上调,p62蛋白下调,差异有统计学意义(P<0.05),提示在URSA患者绒毛组织中自噬水平增强。推测自噬与URSA发生的关系可能为:(1)滋养层细胞受到环境强烈刺激时,细胞中的自噬水平过度会直接导致自噬性细胞死亡;(2)过度自噬可通过自噬溶酶体途径降解细胞中生存素等抗凋亡因子,导致细胞凋亡,从而引发URSA。自噬是一把“双刃剑”。在妊娠早期滋养层细胞处于相对缺氧的环境中,缺氧环境可以诱导自噬的发生,细胞内的自噬体通过收集受损、老化的细胞器,产生新的能量以维持细胞正常增殖分化。如果滋养层细胞受的刺激过度,同样会使细胞自噬反应激烈,使细胞中的细胞器大量受损从而使细胞死亡,最终导致复发性流产。谢冰等[18]研究发现,Beclin-1 mRNA及蛋白、LC3-Ⅱ/LC3-Ⅰ蛋白表达水平在稽留流产的绒毛及蜕膜组织中均上调,提示细胞自噬参与了稽留流产的发生,其主要原因是绒毛及蜕膜组织发生炎性反应。王婧瑶[19]研究发现,IGF2在胚胎停止发育绒毛组织中表达降低,mTOR通路调节自噬的作用缺失,导致自噬活动紊乱,进而使胚胎停止发育。本研究结果提示,自噬相关蛋白在URSA患者绒毛组织中高度表达也有可能是妊娠早期的绒毛组织发生炎性反应,绒毛组织细胞通过自噬过度激活以应对炎性反应,最终使得细胞发生自噬性死亡。
本研究为URSA发病机制的阐明提供了一定理论依据,可考虑将自噬相关蛋白LC3-Ⅱ、p62作为URSA早期诊断的标志物。但复发性流产的致病原因复杂,还需进一步深入研究。
