Strategies to mitigate the adverse effect of drought stress on crop plants—influences of soil bacteria:A review
2021-07-16PriyankaCHANDRAAmoolyaWUNNAVAPoojaVERMAAmareshCHANDRAandRakeshKumarSHARMA
Priyanka CHANDRA,Amoolya WUNNAVA,Pooja VERMA,Amaresh CHANDRAand Rakesh Kumar SHARMA
1 Indian Council of Agricultural Research(ICAR),Central Soil Salinity Research Institute,Kar nal,Har yana 132001(India)
2 Depar tment of Biosciences,Manipal University Jaipur,Rajasthan,Jaipur 303007(India)
3 Indian Council of Agricultural Research(ICAR),Indian Institute of Sugarcane Research,Uttar Pradesh,Lucknow 226002(India)
(Received May 20,2020;revised June 9,2020)
ABSTRACT Drought stress affects plant growth and causes significant issues in meeting global demand for food crops and fodder.Drought can cause physiological,physicochemical,and morphological changes in plants,which negatively affects plant growth and productivity.To combat this under the increasing global threat of water shortage and rapid population expansion,it is crucial to develop strategies to meet global food demands.Plant growth-promoting rhizobacteria(PGPR)may provide a safe solution to enhancing crop yields through various mechanisms.These soil bacteria can provide drought tolerance to crop plants,allowing them to survive and thrive in water-scarce conditions.Productions of phytohormones,free radical-scavenging enzymes,and stress-combating enzymes that can increase tolerance to drought-induced stress are key features of plant-associated microbial communities.This review summarizes the beneficial properties of microbes that help plants tolerate water scarcity and highlights the bacterial mechanisms that enhance drought tolerance in plants.Key Words: abiotic stress,agricultural yield,drought tolerance,food crops,microbe,PGPR,rhizobactoria,water deficit
INTRODUCTION
Agricultural productivity is severely affected by climate change,particularly changes in weather patterns at regional or global scales.Increased anthropogenic activities,such as industrialization,urbanization,deforestation,agriculture,and changes in land-use patterns,lead to the emission of greenhouse gases,which increases the rate of climate change.The effects of climate change include increases in temperature,fluctuations in rainfall patterns,and higher atmospheric carbon dioxide(CO2)concentrations.According to the Intergovernmental Panel on Climate Change(IPCC)report published in 2018,an estimated 1.0°C increase in global temperature has occurred because of industrialization(IPCC,2018).It has also been proposed that the continuation of anthropogenic activities at the current rate could increase the global temperature by up to 1.5°C by the year 2052(IPCC,2018).Due to high temperature,drought,or soil moisture stress,has become one of the major abiotic stresses in arid and semi-arid ecosystems.However,drought can also be caused by low rainfall,salinity,unfavorable fluctuating temperature,and high light intensity.
Plant growth requires certain physiological and physiochemical conditions for optimum growth.Biotic and abiotic stresses such as pathogens,drought,and pH imbalance,can have adverse effects on plant growth,productivity,and yield.Among the abiotic stresses,drought stress is one of the most pressing concerns,especially as a growing population demands a well-functioning agricultural sector with high output(Vurukondaet al.,2016).Drought conditions generate stress in plants,impacting their biochemical and molecular properties,which can result in stunted growth and poor yield(Meenaet al.,2017).These changes mainly occur due to altered metabolic functions—for example,reduced light-harvesting properties because of reductions in photosynthetic pigments.
Several researchers have found and demonstrated the potential of soil microorganisms to combat drought stress(Naylor and Coleman-Derr,2018).Plant growth-promoting rhizobacteria(PGPR)may colonize the rhizosphere,where they confer properties beneficial to plant growth(Candidoet al.,2015).They may also elicit physical or chemical changes associated with plant defense.These PGPR may also enable induced systemic tolerance(IST),whereby the ability of the plants to withstand abiotic stresses is increased due to physical and chemical changes(Sarmaet al.,2012).Thus,PGPR help plants adapt to adverse environmental conditions and can mitigate drought stress by altering responses at the gene level.Many drought-tolerant PGPR strains have been identified as having potential to enhance plant physiological responses to water scarcity,thereby enhancing plant growth and survival under drought conditions(Niuet al.,2018).Inoculation of wheat plants with a consortium of PGPR and arbuscular mycorrhizal fungi resulted in a 41%increase in yield.Mutually beneficial relationships between soil microbiota and plants can result in an overall enhancement in plant growth and higher yields while also conferring drought tolerance in an environmentally friendly manner(Khanet al.,2018).This review summarizes the beneficial properties of microbes that help plants tolerate water scarcity and highlights the bacterial mechanisms that enhance drought tolerance in plants.
EFFECT OF DROUGHT STRESS ON PLANT
Water scarcity is one of the most prevalent abiotic stresses affecting crop plants.Drought stress arises when water availability in soil decreases.Additionally,water is also regularly lost through evapotranspiration due to high atmospheric temperature.Under these conditions,plants decrease leaf water potential and turgor pressure.This also leads to the closure of stomata,which in turn causes plant metabolism to slow down and the cessation of enzymatic reactions.Severe water shortage and lower photosynthetic rates ultimately lead to stunted plant growth and eventual death(Fahadet al.,2017).
Drought stress can lead to certain consequences in plant as follows.One is inhibition of cell enlargement and division.This affects normal biochemical and physiological functions such as ion uptake,photosynthesis,respiration,translocation,and nutrient metabolism.Along with reducing the elongation of cells at the initial stages of plant growth,this inhibition can also affect seed germination.Another is low turgor pressure,which results in dwarfism.A lack of sufficient water reduces leaf growth,thus reducing the photosynthetic area and negatively influencing the photosynthetic pigments(chlorophyllaandb).There is also oxidative damage in thylakoids through reactive oxygen species(ROS)production.This has an impact on plant chlorophyll and carotenoids and causes degradation of the core complexes of photosystems I(PSI)and II(PSII),thus inhibiting essential photochemical processes(Roach and Krieger-Liszkay,2014).
PLANT ADAPTATIONS TO DROUGHT STRESS
Plants employ various methods to promote tolerance to drought stress.Drought tolerance generally varies interand intra-specifically.To assess the degree of tolerance in plants,various drought-related indicators are used,such as morphological characteristics,water potential,tendency to naturalize osmotic pressure,and cell membrane stability.Plants generally survive under drought conditions through the following mechanisms:i)drought tolerance,which is defined as plants’potential to adjust to or tolerate droughtlike conditions;ii)drought escape,which is the ability of plants to conclude their life cycles before the inception of drought and escape from inhospitable conditions by regulating the somatic and reproductive cycles according to the availability of water;iii)drought avoidance,where plants can“avoid”drought-like conditions by retaining water in their cells through various adaptive modifications,achieved through molecular signals that control water uptake and loss,and certain signals result in the closing of stomata,thus reducing transpiration,or bifurcation of plant roots to boost the ability to absorb water from the soil;and iv)drought resistance,where cellular processes,including modification of the cell wall constitution through control of osmotic pressure,cellular elasticity,and protoplasmic resilience to maintain the physical cell structure and its activities,allow the plant to maintain normal cell function and morphology.
Plants employ various mechanisms to survive in waterscarce environments.These processes are assisted by other molecular changes to maintain cell homeostasis.Root cell division increases to augment water uptake.Changes in root architecture in such cases are mediated through chemical signaling,including increased production of phytohormones like auxin,cytokinins,gibberellic acid,and abscisic acid(ABA).Since the stomata close to reduce water loss through transpiration,CO2and nutrient uptake are also affected.Sometimes,plants modify pigments that aid in photosynthesis to maintain cellular functions.Additionally,the amino acids proline and glycine may aid in the maintenance of the structural integrity of the cellular membrane and counterbalancing the deleterious effects of ROS in plants(Hoqueet al.,2008).
PRESENT STRATEGIES TO MITIGATE THE ADVERSE EFFECT OF DROUGHT STRESS
Various strategies are being adopted to meet global demands for food and other plant derivatives under adverse conditions.Plant breeders employ practices such as breeding drought-tolerant varieties or incorporating genetic engineering techniques.
Breeding of drought-tolerant varieties
Conventional plant breeding practices employ methods of propagating superior wild-type varieties of plants that can survive under harsh drought conditions.However,genetic variation in the prevailing germplasm is not a reliable solution as it requires extensive crossing and screening of the potential genotypes(i.e.,those exhibiting favorable characteristics),which can be laborious and time consuming(Kumaret al.,2014).
Genetic approaches for the improvement of drought tolerance in plant
Certain genes and their transcripts,which can have multiple functions,are modified when they undergo droughtinduced stress to improve adaptation to these conditions,resulting in the manifestation of drought-responsive genes(Miret al.,2012).Breeding of genetically superior plants requires focusing on these drought-responsive genes,which code for the production of proteins that are involved in absorption and transportation of water and ions and are responsible for transcriptional regulation,signaling cascades,and structural protection of the cell and its membrane(Khanet al.,2016).To increase stress tolerance,scientists are targeting the genes that code for antioxidant production,growth regulators,and compatible solutes(Royet al.,2011).Expression of the genes that encode for the enzymes choline monooxygenase and beta aldehyde dehydrogenase in higher plants elevates glycine betaine(GB)levels and allows the plants to flourish,even under drought stress(Giri,2011).
The enzyme superoxide dismutase(SOD)is produced by plants during metabolic damage caused by unfavorable environmental conditions.Genes associated with the production of SOD have been incorporated into crops like alfalfa,potato,and rice to produce drought-tolerant transgenic varieties(Perlet al.,1993;McKersieet al.,1996).By incorporating genes encoding ascorbate peroxidase and mono dehydro-ascorbate reductase,researchers have developed transgenic tobacco that is able to produce optimal yields under drought stress(Eltayebet al.,2007).Late embryogenesis abundant(LEA)proteins enable plants to preserve their structural and physiological processes,such as their ionic balance,that are disturbed during water scarcity.These LEA protein-encoding genes may be used to enhance plant growth under drought stress(Browneet al.,2002).
Generation of drought stress tolerance in plant through agronomical practices
Application of growth regulators and osmoprotectants at regular intervals has been shown to have a positive influence on drought stress tolerance in crops.A method called“seed priming”involves the hydration of seeds in a controlled manner,while avoiding the emergence of the radicle,to encourage pre-germinative metabolic activity(Khanet al.,2018).Rice seedlings primed using 5%polyethylene glycol(PEG)and sodium chloride result in better yield under drought stress(Goswamiet al.,2013).Similarly,exogenous application of growth regulators(e.g.,jasmonates,gibberellic acid,and salicylic acid)and osmoprotectants(e.g.,GB)on the foliage of certain plants confers beneficial responses such as better stomatal conductance and transpiration rates as well as photosynthetic and antioxidant activity.These applications can also enhance GB and proline accumulation under water scarcity(Ashraf and Foolad,2007;Rivas-San Vicente and Plasencia,2011).Production of ABA analogs that target ABA receptors in plant cells has also been shown to improve drought-tolerance in plants(Sahet al.,2016).Additionally,silicon usage can improve water absorption,root growth,net photosynthetic rate,stomatal conductance,and plant defense(Habibi and Hajiboland,2013).
EFFECT OF DROUGHT STRESS ON SOIL MICROBIOMES
Soil-inhabiting microorganisms play a vital role in maintaining soil health and plant nutrient uptake(Jiaet al.,2020).These microbes form complex relationships with plant roots and are also affected by biotic and abiotic stresses(Candidoet al.,2013).The life cycles and overall health of plants appear to be dependent on their association with soil microorganisms(whether symbiotic or non-symbiotic)under unfavorable environmental conditions.Water scarcity not only affects plant growth,but also has a strong effect on the soil and its microbiomes.Soil bio-physicochemical properties may change over time,which may alter the water-holding capacity of the soil.This change ultimately influences microbial colonization and plant-microbe interactions(Silva and Lambers,2020).Nonetheless,soil microbiomes may regulate and influence plant growth and yield through various metabolic mechanisms by forming ecological communities(Hartmannet al.,2017).This is achieved through the modification of plant processes and morphologyviaplant-microbe associations,which enhance the tolerance of plants to drought conditions,promoting their survive(Qiaoet al.,2017;Tthet al.,2017).
PGPR ROLE IN MITIGATING DROUGHT STRESS
Rhizospheric microorganisms are beneficial to plants as they are involved in nutrient fixation and other processes that promote physiological responses to biotic and abiotic stresses.These microbes may enhance the tolerance and resistance of plants to pests,pathogens,drought,salinity,and pH imbalances(Goswami and Deka,2020).Plantmicrobe associations may vary as they depend on the type of PGPR,i.e.,extracellular,root colonizing,or endophytic(Gray and Smith,2005).Rhizospheric microorganisms also aid atmospheric nitrogen fixation and uptake of other microand macronutrients such as phosphorous,zinc,and potassium.These microbes may also induce phytohormone and siderophore production,promote signal transduction,and modify immune responses for optimal growth.Table I shows some of the soil bacteria identified in recent years that are known to alleviate drought stress in plant.
Microbes use a variety of mechanisms to survive under adverse conditions,and their association with plants may provide numerous advantages to the host plants growing under stressed conditions(Table I).During drought conditions,PGPR can promote drought tolerance in many crops,including chickpea,maize,mung bean,pea,pepper,tomato,and wheat.Certain drought-tolerant strains of PGPR may extend this property to the plants when they are in an association with them.During drought conditions,stress-induced plant hormones like ethylene are up-regulated in plants,requiring the precursor aminocyclopropane-1-carboxylate(ACC).On the other hand,some PGPR may produce ACC deaminase,which cleaves ACC to alleviate stress(Etesami and Maheshwari,2018).Soil bacteria may also provide additional benefits during plant growth under two simultaneous stress conditions,for example high metal and salinity levels along with drought(Brunoet al.,2020).

TABLE IDifferent mechanisms used by soil bacteria identified in recent years for drought stress alleviation and plant growth promotion
Under drought conditions,AcinetobacterandPseudomonasare able to increase shoot growth,leaf biomass,and overall photosynthetic activity in grape plants(Rolliet al.,2015).The parameters or extent of plant growth may vary depending on the specific plants and microbes,as demonstrated by the effects ofAzospirillum brasilense,Bacillus pumilus,and arbuscular mycorrhizal fungus(AMF)on three types of leguminous trees in desert environments(Bashanet al.,2012).The PGPR may also live as endophytes in different parts of plants,such as the leaves and tubers,by entering the vascular system through the endodermis and root cortex and can positively influence plant growth under stressed conditions(Kushwahaet al.,2020).The colonization frequency of these PGPR represents their effectiveness in adapting to specific ecological niches.It has been shown that endophytes living in desert date palms can successfully cross-colonize in the roots of other palm species as well asArabidopsis(Liet al.,2017).Roots of the desert date palm are able to selectively modify the endophytic community to encourage plant growth under drought conditions.This seems to be a crucial requirement in maintaining the plant and its life cycle in oasis ecosystems(Santoyoet al.,2016).
Nutrient availability
As shown in Fig.1,PGPR may increase the uptake of soil nutrients by plants through fixation,mineralization,and solubilization.Plants are unable to synthesize nitrogen for amino acid and protein production;therefore,they completely rely on soil microorganisms for biological nitrogen fixation.These microbes(e.g.,Azospirillum,Azotobacter(free-living nitrogen-fixing bacteria),Rhizobiumsp.,Biernackisp.,andKlebsiella pneumoniae(symbiotic microbes))may form a symbiotic or non-symbiotic relationship with the plant(Gopalakrishnanet al.,2015).Application of nitrogenfixing PGPR in agricultural fields can restore healthy growth and metabolism,maintain appropriate nitrogen content in the rhizosphere,and improve soil quality and nodule formation.Co-inoculation of bacteria can also have a positive influence on plant growth.Paenibacillus polymyxaandRhizobiumtropicican increase nodule formation,nitrogen content,and dry biomass of the shoots compared to un-inoculated plants under drought conditions(Figueiredoet al.,2008).Bradyrhizobiumsp.enhances plant growth by increasing nitrogen and phosphorus uptake and regulation in lupin under drought conditions(Egamberdievaet al.,2017).Similarly,Bacillussp.ESA 402 can support sorghum growth by positively influencing the photosynthesis rate,the recovery of photosynthesis,and nitrogen accumulation in the shoots(Santanaet al.,2020).
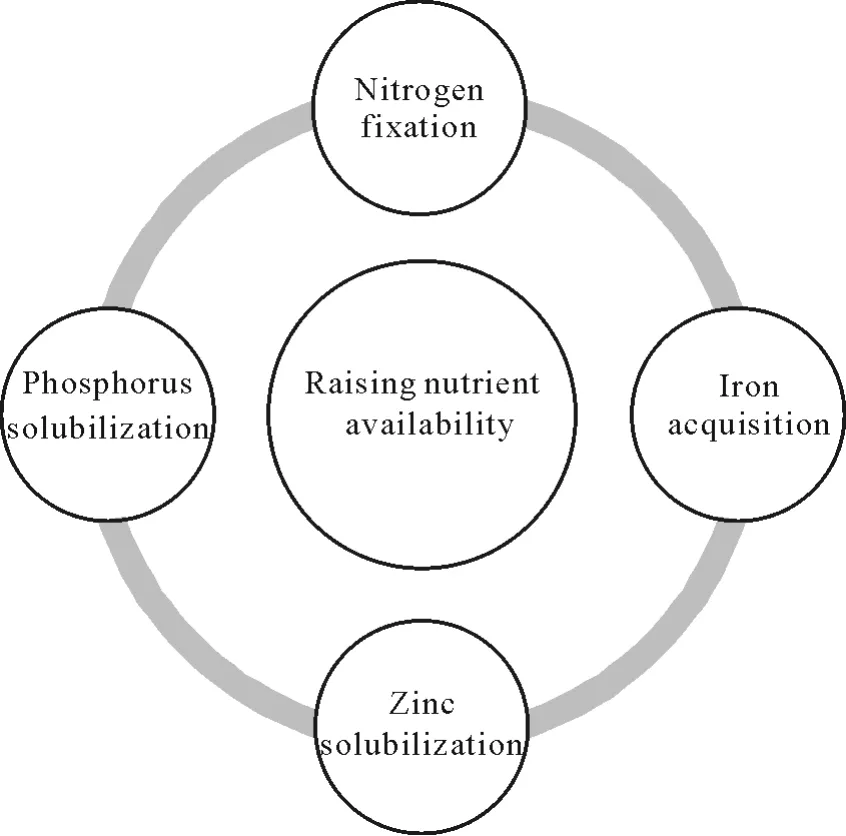
Fig.1 Plant growth-promoting rhizobacteria(PGPR)raising nutrient availability to plants under drought stress.
Nutrient bioavailability for plant uptake is severely affected by drought conditions(Naylor and Coleman-Derr,2018).In such scenarios,PGPR may fix the nutrients by solubilizing complex phosphates.Such nutrient-solubilizing PGPR includeBacillus subtilis,Bacillus megaterium,Pseudomonas fluorescens,andPseudomonas putida.By secreting ironbinding proteins and siderophores,PGPR provide a channel for easy absorption of nutrients during drought stress.Phyllobacteriumproduces siderophores that can be used by strawberry plants to sequester iron,resulting in improved growth and yield and higher-quality fruit under drought stress(Flores-Flixet al.,2015).
Root development
Plant growth-promoting rhizobacteria may influence root development and architecture and lateral root formation,thus allowing the plants to withstand drought stress(Vacheronet al.,2013;Ngumbi and Kloepper,2016).Rhizobacteria influence the growth and differentiation of root cells and amplify root hair density by increasing the number of cortical cells around the radial axis,thus indirectly providing a larger surface area for the root hair-forming epidermal cells(Cohenet al.,2008;Salomonet al.,2014).Rhizobacteria are also able to produce volatile organic compounds(VOCs)that induce division in lateral root founder cells,which further differentiate into the lateral leaf primordia.Studies have suggested that the root alterations caused by PGPR increase water and nutrient absorption by increasing the overall surface area of the root(Timmusket al.,2014).
Plant hormones,such as auxin and cytokinin,produced by PGPR may also promote plant growth and development(Arkhipovaet al.,2007).The soil bacteriumB.subtilisproduces cytokinin and can enhance the growth of lettuce by increasing the amount of this phytohormone in the plant cells.Pseudomonas simiae(aided by various bacterial quorumsensing molecules)can enhance auxin production in the meristem ofArabidopsis,leading to an increase in cell division rate by inducing the expression of an auxin-responsive gene(Verbon and Liberman,2016).This auxin-responsive gene in turn enhances the lateral root growth rate without inhibiting the growth of primary roots(Ortiz-Castroet al.,2011).Auxin signaling is also affected by the bacterial compound indole,which is utilized in various processes such as biofilm formation and virulence.The application of indole may induce lateral root primordium development(Sunet al.,2020).
Bacterial quorum-sensing molecules such as diketopiperazines(DKPs)andN-acyl-homoserine lactones(AHLs)influence root development in auxin-dependent and auxinindependent manners,respectively.Diketopiperazines trigger auxin-inducible gene functions,possibly by binding to the auxin receptor itself.At high concentrations,AHLs induce lateral root growth while inhibiting primary root growth by altering the functional response of cytokinin.Shortchain AHLs at lower concentrations increase primary root elongation by positively influencing meristematic cell divisions involving calmodulin and G-protein signaling.Another molecule,pyocyanin(PCN)modulated by quorum sensing,can regulate root phenotypes independent of both auxin and cytokinin,likely by modifying the levels of ROS in the primary root tipviamanipulation of ethylene levels(Ortiz-Castroet al.,2014).
Bacillus megateriumcan manipulate root morphology by producing the volatile compound dimethyl disulfide.This process involves a unique mechanism through which plant root development is influenced independently of either auxin or ethylene(Meldauet al.,2013).Alcaligenes faecalishas a positive effect on drought-affected maize plants by increasing water and ultimately nutrient uptake and root length by up to 10%compared to the plants not inoculated with the PGPR,thus imparting drought tolerance in the crop(Naseem and Bano,2014).Another study on maize plants reported thatBurkholderia phytofirmansandEnterobactersp.can promote tolerance in drought-affected plants.Plants inoculated with these PGPR showed up to 70%increases in root biomass caused by improved water uptake(Naveedet al.,2014).Similarly,Bacillus thuringiensiscan alleviate drought stress by inducing root hair and lateral root growth in wheat plants(Timmusket al.,2014).
Phytohormone production
Phytohormones are responsible for various physiological processes in plants.Rhizobacteria can produce different plant hormones efficiently,which can support plants,allowing them to thrive when water is scarce(Fig.2).The indole-3-acetic acid(IAA)-producing bacteriaAzospirillum,Bradyrhizobium,andPseudomonascan promote root proliferation(Enebe and Babalola,2018).Cytokinins synthesized by PGPR are involved in physiological processes such as apical dominance,cell division,chloroplast differentiation,leaf senescence,and nutrient mobilization(Vejanet al.,2016).The cytokinin-producing bacteriumB.subtilisincreases plant growth under drought stress conditions(Arkhipovaet al.,2007).Bacillus subtilisalong with several other PGPR(Azospirillum lipoferum,A.brasilense,Acetobacter diazotrophicus,Herbaspirillum seropedicae,Bacillus cereus,B.subtilis,Bacillus licheniformis,Burkholderiasp.,andP.fluorescens)also produce gibberellins or diterpenes,hormones that are involved in fruit and flower growth,seed germination,and floral induction(Bottiniet al.,2004).
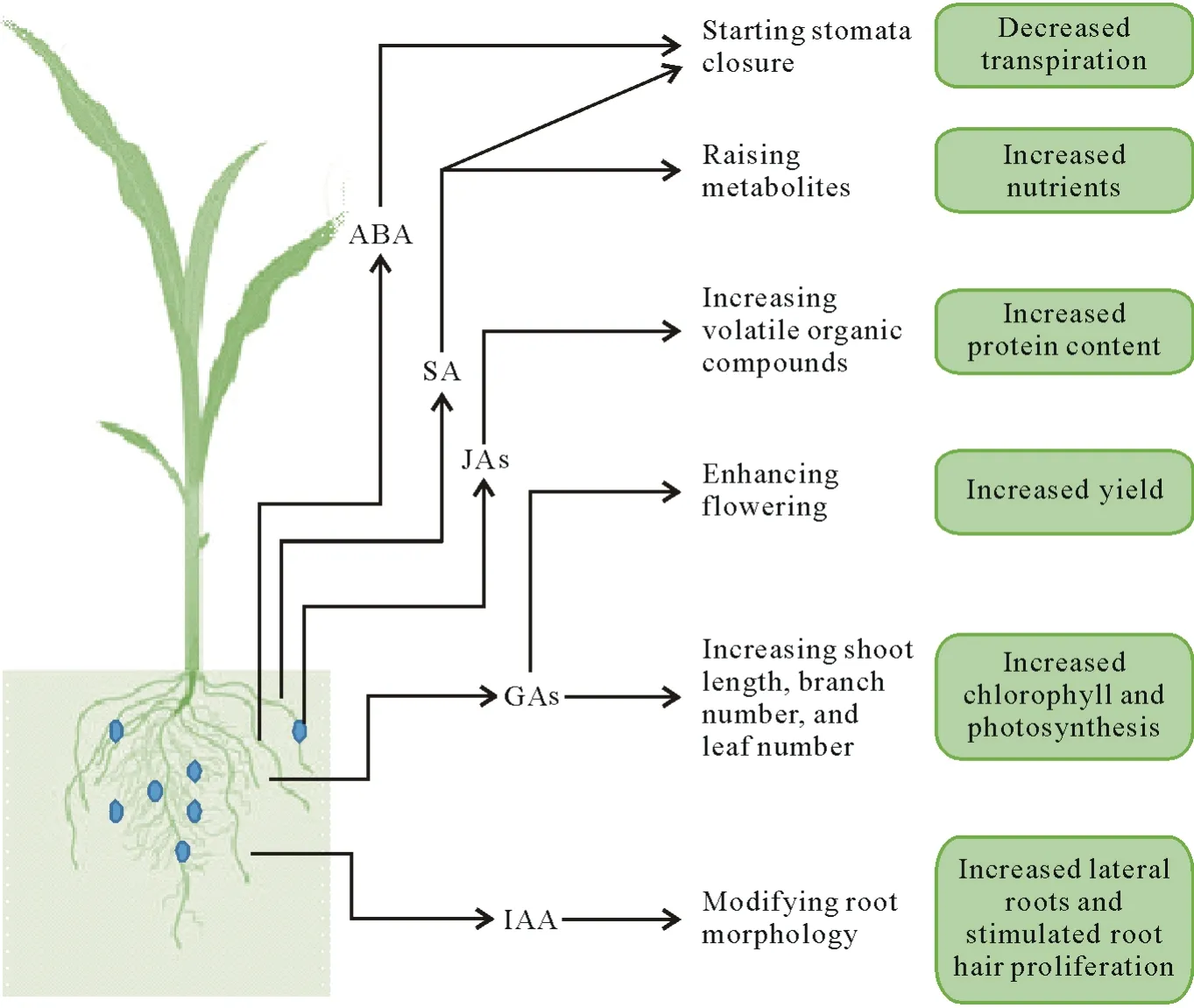
Fig.2 Schematic representation of different plant growth-regulating hormones produced by plant growth-promoting rhizobacteria and their functions in supporting plants under drought stress.ABA=abscisic acid;SA=salicylic acid;JAs=jasmonates;GAs=gibberellins;IAA=indole-3-acetic acid.
Abscisic acid is a stress-signaling hormone that regulates stomatal aperture closure.Studies have reported that inoculation of plants with certain PGPR results in increased levels of ABA and contributes to drought-tolerant properties.Abscisic acid production byA.lipoferumin maize can enhance cell turgidity and relative water content.Reductions in the rate of water loss by 4%and 10%have been observed in grape leaf tissues when inoculated withB.licheniformisandP.fluorescens,respectively(Salomonet al.,2014).
Paenibacillus polymyxaandR.tropicican modify stomatal conductance and help maintain hormone homeostasis,whileAzospirillumsustains cell turgor by adjusting the osmotic pressure in wheat plants(Creuset al.,2004).
Aminocyclopropane-1-carboxylate deaminase production
Ethylene is a plant hormone that promotes fruit ripening along with several other physiological responses in plant life cycles.Under stressed conditions,it is produced in excess,resulting in a cascade of detrimental effects that retard plant growth(e.g.,obstruction of seed germination and root development).As the precursor for ethylene,ACC is found in excessive amounts in plants under drought stress conditions.In such cases,various rhizobacteria can hinder the action of ACC by producing ACC deaminase,which breaks down the ACC molecule intoα-ketobutyrate and ammonium.This decreases the levels of ethylene in the plant,thus protecting it from stress-induced growth inhibition(Chandraetal.,2019).Variovorax paradoxuscan decrease ethylene levels in plant leaves and slow ethylene synthesis in the wild typeArabidopsis thaliana,an ethylene-overproducing plant.Agrobacterium,Achromobacter,Azospirillum,Burkholderia,Enterobacter,Pseudomonas,Rhizobium,andRalstoniahave all demonstrated ACC deaminase production under drought stress(Bressonet al.,2014).By inoculating legume plants with these microorganisms,increases in nodule formation have been observed.Bacillus amyloliquefaciensandAgrobacterium fabrumcan promote considerably higher yields in wheat,whether inoculated individually or combined.These bacteria produce ACC deaminase that catabolizes the ethylene produced in response to drought stress(Zafar-Ul-Hyeet al.,2019).
Plant growth-promoting rhizobacteria may regulate the amount of ethylene in plants by monopolizing the expression of the hormone-producing genesacs(encoding ACC synthase)andaco(encoding ACC oxidase).Burkholderia phytofirmanssignificantly enhances the expression of bothacsandacoinA.thaliana(Poupinet al.,2016),whilePhyllobacterium brassicacearummore specifically targets the expression ofacs7andacs11in the same plant.Phyllobacterium brassicacearuminoculation improves the elongation of root hairs;however,this effect is only partially dependent on the ethylene-signaling pathway(Bressonet al.,2014).
Exopolysaccharide(EPS)production
Soil bacteria may improve soil health and positively influence plant drought stress tolerance by mediating physiochemical and hydrological alterations in the rhizosphere.Microbial biofilms are made up of specific arrangements and attachments of bacterial cells through EPS.This EPS is made up of homo-and heteropolysaccharides on the cell surfaces as a capsule or slime.Bacteria that produce EPS confer beneficial properties on drought stress-affected plants by producing a rhizosheath around the roots,preventing desiccation.These rhizosheaths also maintain soil moisture content even in dry sandy or desert soil(Naseemet al.,2018).The presence of EPS-producingB.subtilisfacilitates water availability for plants by reducing evaporation.The EPS may improve soil water retention properties and hydraulic conductivity as it has high water-holding capacity,which may modify soil matrix structures and connectivity of pore spaces,ultimately improving plant growth(Zhenget al.,2018).
Production of EPS by plant-associated microbes plays a significant role in supporting plant growth during times of water scarcity.Plant roots are covered by a hydrophilic bacterial EPS-containing biofilm,providing protection against desiccation.Such biofilms are formed byAcinetobacterandPseudomonasstrains around plant roots that predominantly function as an additional sheath to shield the roots from soil dryness(Rolliet al.,2015).
Polysaccharide constituents of EPS affect its water retention ability,which may exceed 70 g water per gram of polysaccharide.The EPS produced byAzospirillumsignificantly enhances tolerance to water stress in plants by amending the soil structure and its aggregation properties.Sunflower plants treated with EPS-producingRhizobiumhave been shown to improve water and nitrogen uptake,attributed to the increased root-adhering soil(RAS)/root tissue(RT)ratio and macroporosity(Vurukondaet al.,2016).Rhizobia can ameliorate drought stress in okra by improving soil aggregation in the rhizosphereviasufficient EPS production(Yadavet al.,2018).Drought-tolerantBacillusandPseudomonasstrains produce EPS that increases the RAS/RT ratio and macroaggregate stability,which ultimately improves nutrient and water uptake from soils.Additionally,EPS can act as an emulsifier,protecting biomembranes and quenching ROS.Alginate production byB.thuringiensisalso stimulates drought tolerance in wheat(Timmusket al.,2014).
The synthesis and chemical constitution of EPS are governed by the bacterial growth phase,the composition of the growth medium(carbon/nitrogen ratio),and other environmental factors.Stress conditions may also trigger the formation of guanylate cyclase in cells,subsequently leading to the production of EPS.The geneSp7and its mutantsnoeJandnoeL(encoding mannose-6-phosphate isomerase and GDP-mannose 4,6-dehydratase,respectively)are involved in the synthesis of mannose and fructose,which are the major constituents of EPS,inA.brasilense(Lerneret al.,2009).Water deficit conditions can induce the expression ofalg(encoding mannuronan C-5-epimerase)inPseudomonas aeruginosa,which has a complicated role in reducing the effect of drought stress(Muhammadi and Ahmed,2007).Thus,the EPS produced by PGPRduring water shortage conditions can promote the development of roots and shoots and the total dry weight of plants,showing that EPS can influence soil aggregation and water regulation in the rhizosphere.
Osmolyte production
Production of osmolytes in the presence of PGPR in response to drought stress can promote plant growth(Ngumbi and Kloepper,2016).The concentration of proline,an important osmolyte,increases in plants when exposed to water scarcity.Proline promotes plant growth by preserving cell membranes,proteins,and other subcellular structures,reducing the number of free radicals,and buffering cellular redox potential.Inoculating maize plants withP.putidaproduces positive effect due to the accumulation of proline,which leads to increases in the concentrations of amino acid and soluble sugar,relative water content,leaf water potential,and plant biomass,resulting in better drought tolerance(Sandhyaet al.,2010).However,a recent study reported lower osmolyte concentrations in sugarcane after treatment with PGPR(Chandraet al.,2018).Bacterial production and accumulation of some xeroprotectants,such as trehalose,may preserve the vitality of plants by providing osmotic stress tolerance.Trehalose acts as an osmoprotectant in plants when their cells are dehydrated due to external environmental conditions.It also acts as a signaling molecule and a non-reducing sugar that confers stability to enzymes and proteins produced under drought conditions and protects the integrity of the cell membrane through the expression of the trehalose-6-phosphate synthase gene(Vlchezet al.,2016).
The inoculation of PGPRin crop plants including maize,sorghum,pea,potato,chickpea,rice,and soybean results in increased levels of proline in the plants when grown under drought stress(Ngumbi and Kloepper,2016).These proline levels are enhanced by several PGPR,including
B.thuringiensisin the case ofLavandula dentata(which produces higher levels of proline in plant shoots)(Armadaet al.,2016)andBacillus polymyxain tomato(which invokes enhanced growth)(Shintu and Jayaram,2015).Arthrobactersp.,Bacillussp.,andBurkholderiasp.also increase proline levels in plants under drought stress(Dodd and Pérez-Alfocea,2012).It has been shown that a consortium of PGPR strains(B.cereus,B.subtilis,andSerratiasp.)can protect cucumber plants from dehydration,coincident with 4-fold increase in proline level compared to those of untreated controls(Wanget al.,2012).The higher proline contents due to inoculation of PGPR might be beneficial for plants as it protects their cellular structures and functions under drought stress(Sandhyaet al.,2010).
Volatile organic compound production
The VOCs produced by PGPR may directly or indirectly influence optimal plant growth,disease tolerance,and drought stress tolerance.These microbial VOCs have also been known to affect and regulate plant endogenous auxin homeostasis and regular nutrient uptake(Ngumbi and Kloepper,2016).Upregulation of auxin homeostasis inArabidopsisupon treatment withB.subtilishas been reported,highlighting the fact that these microbial VOCs possess the potential to stimulate the biosynthesis of the osmoprotectants choline and GB,which induce drought tolerance in plants(Sharifiand Ryu,2018).The VOCs produced byB.subtilisalso induce tissue-specific gene regulation of high-affinity K+transporter(HKT1),which inhibits uptake of Na+ions by roots while enhancing the shoot to root translocation process,thus imparting IST to plants under saline conditions(Sharifiand Ryu,2018).
Bacillus amyloliquefaciensandB.subtilisproduce the VOCs 3-hydroxy-2-butanone(acetoin)and 2R-3R-butanediol,which promote increased growth inArabidopsisby monopolizing gene expression involved in the maintenance of cell wall structure.Application of 2R-3R-butanediol produced byPseudomonas chlororaphiscan influence stomatal aperture closure inArabidopsis.This VOC also influences numerous phytohormone-signaling pathways that include ethylene,jasmonates,and salicylic acid(Asariet al.,2016).Under drought stress,wheat leaves produce several terpenoids and benzenoid compounds,includingαphellandrene,α-pinene,benzaldehyde,β-pinene,camphene,geranyl acetone,limonene,and para-cymene(Timmusket al.,2014).Production ofβ-pinene,benzaldehyde,and geranyl acetone in plants also increases under stressed conditions.However,it has been observed that treatment withB.thuringiensisdecreases these VOC emissions compared to untreated plants under the same conditions,thereby decreasing the loss of carbon content and thus increasing plant growth.Bacteria also produce certain chemicals used in cell to cell communication,generally known as quorum-sensing molecules.These signaling molecules,including AHLs,may also induce systemic tolerance against stress(Filgueiraset al.,2020).
Antioxidant defense
Drought stress enhances the production of ROS that affects the ambient redox regulatory state of plant cells(Laxaet al.,2019).In such conditions,the assimilation of antioxidant enzymes such as catalase(CAT)and peroxidase have been shown to protect the plant from oxidative injury by decreasing the degree of damage caused.The presence of soil bacteria may induce drought tolerance by modulating the antioxidant system.Inoculation withB.pumilusandBacillus firmusresults in significant increases in levels of ROS-scavenging enzymes such as CAT,ascorbate peroxidase,and SOD in potato plants,with the CAT activity being 1.8 times higher than that in untreated plants(Gururaniet al.,2013).The same increases in CAT activity have been shown after inoculation withP.fluorescensandB.subtilisin potato plants,revealing that the ability of the plants to tolerate drought stress and resist the associated damage is linked to increased levels of ROS-scavenging enzymes.This positive correlation between the increased levels of CAT in plants and drought tolerance has also been observed in other plants,including cucumber,maize,and wheat(Sandhyaet al.,2010;Wanget al.,2012).Rhizophagus intraradicesandPiriformospora indicareduced the malondialdehyde and hydrogen peroxide contents as well as electrolyte leakage in finger millet seedlings under drought conditions as compared with control plants;proline content,along with secondary metabolite and phenol accumulation,also increased significantly,with antioxidant enzyme activity resulting in an up to 25%decrease in drought-induced lipid peroxidation(Tyagiet al.,2017).Bacterially mediated proline accumulation and glutathione reductase activity have also been reported as mechanisms that can reduce drought stress in Guinea grass(Moreno-Galvánet al.,2020a).Based onBacillus-maize interactions,antioxidant response to bacterial inoculation under drought stress has been demonstrated to be dependent on proline accumulation and decreases in ascorbate peroxidase and glutathione reductase activities(Moreno-Galvánet al.,2020b).
During plant-microbe interactions,soil bacteria produce several chemical compounds for signaling purposes.These molecules,which are produced by PGPR in the rhizosphere,are known as microbe-associated molecular patterns(MAMPs).They include simple amino acids and sugars as well as large polysaccharides and proteins(Rosieret al.,2018).These MAMPs may also trigger plant defense systems,causing them to produce ROS,which further results in high proline production and activation of different antioxidant systems.Proline accumulation during different plant stresses has been well documented and described.Proline is a signaling molecule,which may act as an effective osmolyte,metal chelator,and antioxidative defense molecule(Hayatet al.,2010).It has also been reported that different signaling molecules produced by soil bacteria commonly known as nodulation factors(NFs)may enhance cell division in symbiotic associations between microbes and plants.The higher rates of cell division and endophyte interactions may increase ROS production at the early stage of infection(Montielet al.,2018).To combat the ROS produced during abiotic and biotic stresses,plants produce enzymatic and nonenzymatic antioxidants and increase proline accumulation.Soil bacteria associated with plants may also produce extracellular enzymatic and nonenzymatic antioxidants along with some phenolic compounds,which further help the plant to detoxify the ROS and convert these species into stable nonreactive species(Fig.3).
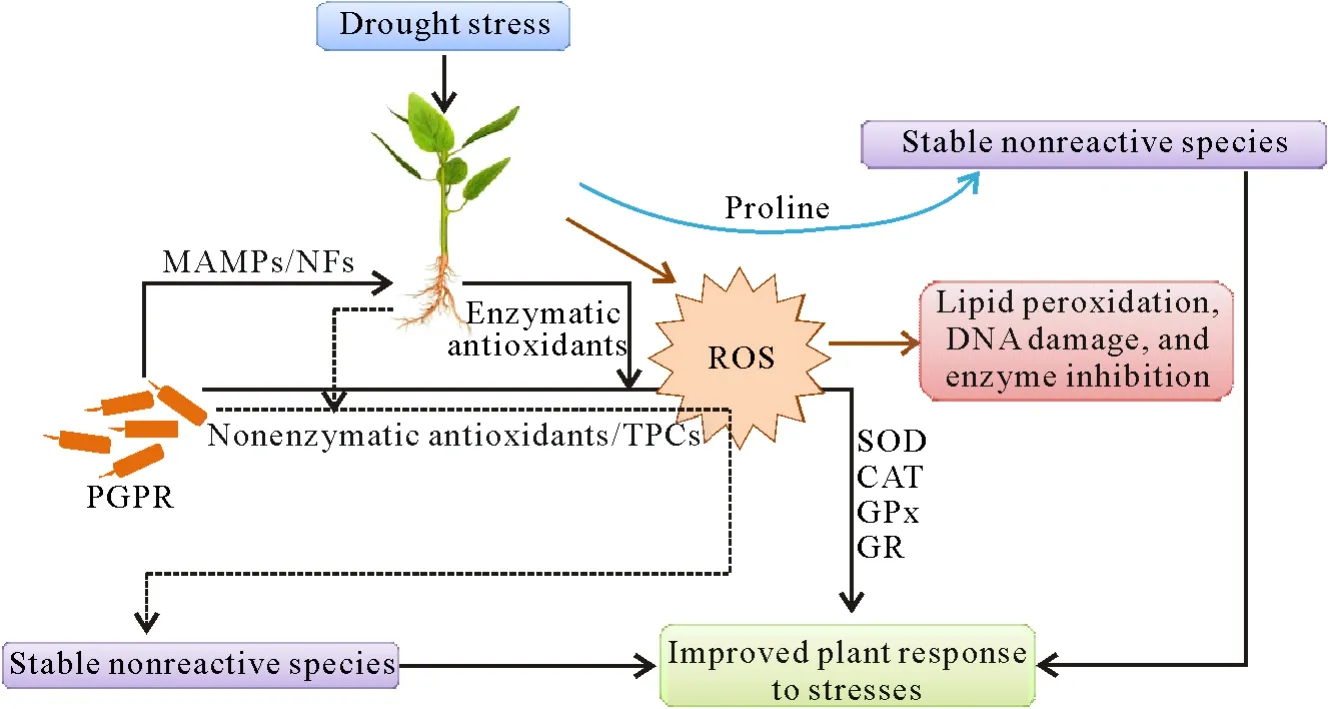
Fig.3 Detoxification of reactive oxygen species(ROS)to stable nonreactive species during drought stress by the enzymatic and nonenzymatic antioxidants produced by the plant itself and plant growth-promoting rhizobacteria(PGPR).MAMPs=microbe-associated molecular patterns;NFs=nodulation factors;TPCs=total phenolic compounds;SOD=superoxide dismutase;CAT=catalase;GPx=glutathione peroxidase;GR=glutathione reductase.
Metabolic reprogramming
Recent studies have reported that microbes may be involved in metabolic and molecular reprogramming in plants under drought stress.Some metabolites including succinic acid,pyruvic acid,dihydroxyacetone,thiamine pyrophosphate,and uridine diphosphate are significantly reduced in wheat during drought stress,while treatment withBacillus velezensissignificantly enhances these metabolites to help combat the stress(Abd El-Daimet al.,2019).Azotobacter chroococcumandA.brasilenseenhance the accumulation of ABA,proteins,soluble sugars,phenols,flavonoids,and oxygenated monoterpenes in pennyroyal under drought stress(Asghariet al.,2020).Additionally,a bacterial consortium includingB.subtilis,B.thuringiensis,andB.megateriumhas been shown to induce significant accumulation of aspartate,glycerol,riboflavin,L-asparagine,nicotinamide,and 3-hydroxy-3-methyglutarate in the leaves of chickpea under drought stress(Khanet al.,2019b).Thus,the metabolic alterations induced by drought can be further modified by plant-associated bacteria to reduce drought impacts.
Regulation of stress-responsive genes
The presence of PGPR may modulate and modify stressresponsive genes that impart IST in plants.Certain genes,e.g.,erd15(encoding early response to dehydration 15)andbab18(encoding late embryogenesis abundant protein),are induced inArabidopsiswhen inoculated withP.polymyxa(Sukweenadhiet al.,2015).Inoculation of a consortium includingB.cereus,B.subtilis,andSerratiasp.has been shown to improve tolerance to drought in cucumber plants by conserving photosynthetic activity through inhibition of the downregulation of ascorbate peroxidase genes,rbcL(encoding RuBisCO large subunits)andrbcS(encoding RuBisCO small subunits).A macroarray analysis of sequence tags after treatment withRhizobium etlishowed overexpression of trehalose-6-phosphate synthase gene,a gene that oversees regulation of carbon and nitrogen metabolism,in bean plants(Suárezet al.,2008).Bacillus licheniformiscan induce the over expression of several stress-responsive proteins(adenosine kinase,protein 10,dehydrin-like protein,early nodulin,S-adenosylmethionine synthetase,and vacuolar H+-ATPase)under water-scarce conditions in pepper plants(Vurukondaet al.,2016).Similarly,B.amyloliquefaciensandA.brasilenseinduce upregulation ofapx1(encoding ascorbate peroxidase),sams1(encodingS-adenosyl-methionine synthetase),andhsp17.8(encoding heat shock protein)in wheat leaves(Tiwariet al.,2017).Gluconacetobacter diazotrophicuscan activate ABA-dependent signaling genes in sugar cane(Vargaset al.,2014),andP.aeruginosacan up-regulate the expression ofdreb2 A(encoding dehydrationresponsive element-binding protein),cat1(encoding CAT),anddhn(encoding dehydrin)in mung bean plants(Sarma and Saikia,2014).Enterobactersp.has been shown to induce expression of salt stress-responsive genes(rab18,rd29 A,rd29B),regulons of ABA-responsive elements(abre),dehydration-responsive elements(DRE),and dehydrationresponsive element-binding proteins(dreb2b),which cause ABA-independent activation under high salinity conditions inArabidopsis(Kimet al.,2014).Pseudomonas chlororaphisprompts transcription and upregulation of the jasmonic acid marker genesvsp1andpdf-1.2,the salicylic acid-regulated genePR-1,and the ethylene-responsive genehel,while causing downregulation of drought-signaling response genes inA.thaliana(Choet al.,2013).
CONCLUSIONS AND FUTURE PROSPECTS
Plant growth-promoting rhizobacteria have the potential to confer tolerance to abiotic and biotic stresses including drought in plants.Three important interconnected factors affect how productive ecosystems are:plants,soils,and microbes.Soil microbes may enhance plant growth indirectly or directly by producing a variety of chemicals.The presence of consortia of microbes having different properties to deal with drought stress and enhance the plant growth may provide better drought tolerance.Similarly,the presence of bacteria with multiple properties that enhance plant survival under different stresses(e.g.,metal toxicity,salinity,xenobiotics,and pathogen attack)can also increase the chances of plant survival when more than two stress conditions occur,including drought.Furthermore,it is important to understand the chemical signaling and molecular mechanisms through which PGPR affect plants and interact with them during stressed conditions.Specific microbial products and processes could be developed for exogenous application to increase their beneficial properties,allowing more extensive application in agricultural systems,especially in drought-prone areas.Similarly,the application of genetically modified PGPR strains could also be assessed in the field.However,the risk involved must be evaluated before the use of such techniques at larger scales.Overall,the treatment of crop plants with PGPR is likely to be beneficial in conferring drought tolerance in plants,allowing improved plant growth,metabolism,and yield.The development of microbe-based ecofriendly techniques for sustainable agriculture under drought stress conditions should be encouraged.
ACKNOWLEDGEMENT
The authors are thankful to Dr.Deepa Sarabhai,assistant professor,Department of Languages,Manipal University,Jaipur,India for improvements in the language of the manuscript.
杂志排行

Pedosphere的其它文章
- Preface Earthworms in soil ecology and organic waste management
- Letter to the Editor Field and laboratory investigations of Lumbricus badensis ecology and behaviour
- Advances in fungal-assisted phytoremediation of heavy metals:A review
- Study of oxidative stress cadmium(Cd)-induced in Eisenia fetida based on mathematical modeling
- Behavior and respiration responses of the earthworm Eisenia fetida to soil arsenite pollution
- Biological weathering of phlogopite during enriched vermicomposting