Advances in fungal-assisted phytoremediation of heavy metals:A review
2021-07-16MuhammadKHALIDSaeedURRAHMANDanialHASSANIKashifHAYATPeiZHOUandNanHUI
Muhammad KHALID,Saeed UR-RAHMAN,Danial HASSANI,Kashif HAYAT,Pei ZHOU,*and Nan HUI,*
1 Key Laborator y of Urban Agriculture,School of Agriculture and Biology,Shanghai Jiao Tong University,Shanghai 200240(China)
2 Joint International Research Laborator y of Metabolic&Developmental Sciences,Key Laborator y of Urban Agriculture(South)of Ministr y of Agriculture,Plant Biotechnology Research Center,Fudan-SJTU-Nottingham Plant Biotechnology R&D Center,School of Agriculture and Biology,Shanghai Jiao Tong University,Shanghai 200240(China)
(Received March 9,2020;revised May 15,2020)
ABSTRACT Trace metals such as manganese(Mn),copper(Cu),zinc(Zn),and iron(Fe)are essential for many biological processes in plant life cycles.However,in excess,they can be toxic and disrupt plant growth processes,which is economically undesirable for crop production.For this reason,processes such as homeostasis and transport control of these trace metals are of constant interest to scientists studying heavily contaminated habitats.Phytoremediation is a promising cleanup technology for soils polluted with heavy metals.However,this technique has some disadvantages,such as the slow growth rate of metal-accumulating plant species,low bioavailability of heavy metals,and long duration of remediation.Microbial-assisted phytoremediation is a promising strategy for hyperaccumulating,detoxifying,or remediating soil contaminants.Arbuscular mycorrhizal fungi(AMF)are found in association with almost all plants,contributing to their healthy performance and providing resistance against environmental stresses.They colonize plant roots and extend their hyphae to the rhizosphere region,assisting in mineral nutrient uptake and regulation of heavy metal acquisition.Endophytic fungi exist in every healthy plant tissue and provide enormous services to their host plants,including growth enhancement by nutrient acquisition,detoxification of heavy metals,secondary metabolite regulation,and enhancement of abiotic/biotic stress tolerance.The aim of the present work is to review the recent literature regarding the role of AMF and endophytic fungi in plant heavy metal tolerance in terms of its regulation in highly contaminated conditions.
Key Words: arbuscular mycorrhizal fungi,bioremediation,endophytic fungi,environmental stress,metal tolerance,microbe,plant transporter,soil contaminant
INTRODUCTION
Microbes are cosmopolitan in nature and even reside in habitats with extreme environmental conditions.Many of them can make biofilms for tolerating harsh environmental surroundings including arid conditions and high temperatures(Maet al.,2016).The soil around plant roots is a natural habitat for different kinds of microorganisms including fungi,bacteria,protozoa,and algae.The microbes associated with plants are important for their hosts,greatly influencing their overall performance in different situations.At the root surface,they communicate through signal molecules to attain structural stability in an exopolysaccharide matrix;in this way,they perform a particular functionviaquorum sensing(Baiset al.,2004).The rhizospheric and endophytic microbiota contributes to plant protection,stability,productivity,growth,and phytoremediation.The use of metal resistance and plant growth-promoting features of rhizospheric or endophytic microorganisms is one of the most cost effective and ecologically friendly strategies(Rajkumaret al.,2010).
Due to the presence of a chelation system and metal sequestration abilities,fungi can alleviate heavy metal stress(Janoukovet al.,2006;Alyet al.,2011).Fungi develop a symbiotic relationship with most terrestrial plants and are an important component of an efficient ecosystem.Several studies have shown their beneficial interactions with different host plants(Smith and Read,2008).To build symbiotic relationships,plants recruit different beneficial fungi through a communication process using signal molecules(Smith and Read,2008).Such mutualistic symbionts include ectomycorrhizal fungi(EMF),arbuscular mycorrhizal fungi(AMF),and dark septate endophytes(DSEs).Plant associated AMF mainly belong to Zygomycota(150 species),and EMF mainly include basidiomycetes with a few zygomycetes and ascomycetes(600 species).Ectomycorrhizal fungi are mainly found in association with specific woody trees,while AMF have been reported in association with a variety of plants from herbs to woody species.Arbuscular mycorrhizal fungi colonize plant roots and proliferate in the rhizosphere region in the form of microscopic filaments,assisting in the translocation of nutrients to the host plant(Latefet al.,2016).It has been extensively reported that AMF develop symbiotic relationships with almost 90%of plants and provide multifaceted amenities such as enhanced metabolism and growth of the host plant under normal or abiotic/biotic stress conditions.Overall,AMF are immensely important in a well-developed ecosystem because of their capability to build mutualistic relationships with the majority of terrestrial plants,as well as some wetland plants(Latefet al.,2016).Some plant species are non-mycorrhizal,such as a few members of the Cruciferae and Amaranthaceae;however they have alternative substitute relationships with different endophytes(Usuki and Narisawa,2007).
All plant species analyzed so far have their own endophytic microbial community.Endophytes are microbes that colonize the internal tissue of a host plant without causing any harm or infection.Over the last few decades,the attention of the scientific community towards endophytes and their bioactive compounds has increased,with a higher number of publications on the subject especially focusing on the study ofEpichloë,Frankia,Piriformospora,andRhizobiumspecies(Chibucos and Tyler,2009;Stadler,2012).Endophytic fungi are extremely diverse,having broad(non-clavicipitaceous fungi)to narrow(clavicipitaceous mutualistic fungi)host ranges and asymptomatically localizing in internal plant tissues such as roots,stems,leaves,shoots,seeds,and even bark(Rodriguezet al.,2009).Some fungi have a higher level of adaptation to certain physiological conditions and usually colonize specific plant organs or tissues.It is quite interesting that after over 400 years of continuous evolution,there are plants that still rely on a symbiotic association with fungi to cope with stressful conditions and to maintain healthy growth(Alyet al.,2011).Fungi also participate and have a potential role in element cycling,transformation of rock and minerals,bioweathering,cycling of different elements,fungal-metal interactions,and transformation of organic and inorganic compounds(Gadd,2007).Endophytes employ different metabolic pathways which are very important for the biotransformation of organic substances and different types of pollutant remediation(St˛epniewska and Kuźniar,2013).To date,many studies have reviewed bioremediation through bacterial strains,but the role of AMF and endophytic fungi in such studies has been mostly overlooked.The contribution of AMF and endophytic microbiota in plant protection,stability,productivity,growth,and phytoremediation is summarized in Fig.1.The aim of the current review is to provide information regarding the role of AMF and endophytic fungi in phytoremediation and the benefits they provide to their host plants.

Fig.1 Scheme showing the positive influences of endophytic fungi and arbuscular mycorrhizal fungi(AMF)on plant overall performance under stresses caused by heavy metals and trace elements.The heavy metals and trace elements include Cu,Cd,Ni,Zn,As,Hg,La,Pb,Fe,U,B,Mn,Cr,Co,Al,and Cs.
ARBUSCULAR MYCORRHIZA AND PLANT INTERACTIONS:METAL TOLERANCE
Generally,mycorrhizal fungi are associated with plant roots in the form of an internal or external network,thus increasing the surface area and absorption capacity of a plant’s root system.Excessive uptake of essential or non-essential heavy metals from soil by plants can cause adverse effects.The AMF enhance plant resistance and heavy metal tolerance,but their influence on plants growing on contaminated medium largely depends on plant species,fungal species,and the type of associated heavy metal.Inducing resistance to heavy metals is not related to reduced heavy metal uptake;there are some reports which show hyperaccumulation by plants associated with AMF(Carvalhoet al.,2006;de Andradeet al.,2008;de Souzaet al.,2012).However,convincing evidence proves that AMF hinder heavy metal transport from root to shoot,thus regulating the partitioning of heavy metals(Christieet al.,2004;Wanget al.,2007;Wuet al.,2016).For example,Diversispora spurcumandFunneliformis mosseaecause retention of heavy metals in the underground part of plants and thus have been shown to reduce zinc(Zn),lead(Pb),and cadmium(Cd)contents in the shoot in comparison to the roots in maize plants(Zhanet al.,2018).In some cases,potentially toxic effects decreased because metal concentrations were reduced as a consequence of the improvement in phosphorus(P)nutrition and growth of the mycorrhizal plant(Chenet al.,2003).For instance,the mycorrhizal poplar has been shown to have lower stress symptoms even with higher concentrations of copper(Cu)and Zn in its tissues(Linguaet al.,2008).Some studies have shown that AMF alleviate heavy metal stress by hindering its uptake by the host plant.For example,AMF attenuated the heavy metal impact onCalendula officinalisdevelopment by reducing the uptake of heavy metals(Cd and Pb)and enhancing the beneficial secondary metabolites in comparison to non-mycorrhizal plants(Hristozkovaet al.,2016).Similarly,reduced Zn uptake and concentration was observed in mycorrhizal tomato plants(Watts-Williamset al.,2013).Immobilization of heavy metals has also been attributed to the changes in biogeochemistry made by mycorrhizae in the rhizosphere region(Kangwankraiphaisanet al.,2013).This was found in indigenous plants growing at Zn smelting and disposal sites,which have a higher capacity for Zn accumulation and sediment emending(72 540±5 092 mg kg-1dry weight)compared to the same plants treated with fungicide,halting AMF colonization.Likewise,Zn immobilization and prevention of arsenic(As)translocation have been shown by some studies(Chenet al.,2012;Chen X Wet al.,2013).Heavy metals also cause a degree of damage to the fungal cells themselves.The propagules of AMF remain in soil as infectious units if a favorable environment and compatible host are not available.In pre-symbiotic states,the walls of the hyphae are thinner and highly exposed to the noxious heavy metals in soil.Spore counts are generally used to determine successful colonization rate,and usually spore count frequency is lower in heavy metal-contaminated areas(Lpezet al.,2000).Arbuscular mycorrhizal fungi propagate by producing mycelia or spores that separate through a fragmentation process and develop into new hyphae.However,these indigenous young propagules always persist in soil to some extent and are resistant to heavy metals,enhancing host plant heavy metal tolerance upon colonization.For instance,the isolateRhizophagus intraradicesBr1 from the heavy metal-resistant plantViola calaminariainduces high resistance in maize,tomato,andMedicago truncatula,unlike the ecotypes belonging to the same species(Kaldorfet al.,1999).Some reports have shown the reduction of spore production by mycelia;however,this depends on the species,degree of tolerance,and even the type of metal(Klauberg-Filhoet al.,2002).Studies regarding the contribution of AMF to heavy metal and trace element tolerance are listed in Table I.
ROLE OF AMF IN METAL UPTAKE AND DISTRIBUTION
A number of advanced techniques have been developed to investigate the uptake,translocation,and transformation of metals by AMF such as inductively coupled plasmamass spectrometry(ICP-MS),scanning electron microscopy with energy dispersive spectroscopy(SEM-EDS),and transmission electron microscopy with energy dispersive spectroscopy(TEM-EDS).For example,the direct influences of AMF on chromium(Cr)translocation and transformation in the soil-plant continuum have been studied,showing that AMF can immobilize quantities of Crviareduction of Cr(VI)to Cr(III),forming Cr(III)-phosphate analogues(Wuet al.,2015).Similarly,light microscopy(LS)and transmission(TEM)and scanning electron microscopy(SEM)studies showed that inoculation ofLeucaena leucocephalawith AMF resulted in better growth in soil with high As concentration,as well as significant As removal from the soil(Schneideret al.,2013a).The movement of essential metals through the mycorrhizal uptake pathway begins with their uptake through the plasma membrane of the extraradical mycelium(Fig.2).

Fig.2 Scheme showing plant heavy metal(HM,red star)distribution and acquisition(uptake,detoxification,translocation,and accumulation).Uptake of metals takes place through either the mycorrhizal or direct pathway.The extraradical mycelia(ERM)are in direct contact with HMs in the soil and possess high-affinity metal transporters that translocate HMs via dense hyphae to intracellular spaces,before finally delivering them to the roots at the symbiotic interface.Fungal and plant metal transporters include CCC2(Cu-ATPase),CTR(fungal Cu transporter),COPT(plant Cu transporter),CDF(cation diffusion facilitator),ZRT1(Zn transporter),FTR1(Fe permease),ZIP(Zn-Fe permease),SMF1(Mn transporter),and OPT(oligopeptide transporter).The metal transporters,which are located in both the epidermal cells and root hairs,are involved in the direct pathway of HM translocation.Once HMs are taken up by the roots,they are then sequestered,detoxified,or translocated along the apoplastic or symplastic pathways towards vascular tissues.Heavy metals enter the root cell via plasma membrane channels or transporters,including Ca2+channels(implicated in Cd2+uptake in endodermal cells),NRAMPs(natural resistance-associated macrophage proteins),ZIPs(Zn2+transporters),ZNT1(transporting apoplastic Zn2+and Cd2+into the cytosol),IRT1(transporting apoplastic Mn2+,Fe2+/Fe3+,Zn2+,and Cd2+into the cytosol),Mte1(exporting HMions outside of the cell or transporting Cu2+/Ni2+into vacuoles),and HMA5(translocation and detoxification of Cu).The detoxification of HMs in root cells is carried out by chelators such as glutathione(GSH),metallothioneins(MTs),and phytochelatins(PCs).Moreover,free and chelated HM ions are sequestered in cell vacuoles by HMA3(transporting cytosolic Co2+,Zn2+,Cd2+,and Pb2+into vacuoles),MTP1/8(mobilizing excessive Zn2+from the cytosol into vacuoles),and ABCC1/2(transporting cytosolic Cd-PCs and Hg-PCs to vacuoles).Such sequestering in Golgi bodies occurs via MTP11(metal tolerance proteins)and ZnT1(transporting excessive Zn2+from the cytosol into vacuoles).Sequestered HMs inside the vacuole can also be excreted back to the cytoplasm through NRAMP3/4(exporting vacuolar Fe2+/Mn2+into the cytosol)and ZIP1(Zn2+transport).Other putative proteins such as ZNT1(transporting excessive Zn2+from the cytosol into vacuoles)and HMA2/4 are mainly involved in the loading of HMs to the vascular tissues of plant roots.
At a molecular level,the genome ofRhizophagus irregularishas been analyzedin silico,identifying several genes that code for proteins that mediate the transport of important metals such as iron(Fe),Zn,and Cu from the soil medium(Tisserantet al.,2013;Tamayoet al.,2014).Specifically,the Cu-ATPases,natural resistance-associated macrophage proteins(NRAMPs),vitamins,the influx mediator of Fe/Zn(RiFTR1,RiZRT1),and the vacuolar mobilizer of Cu/Zn(RiCTR2,RiZRT3)have been identified,but currently there is no evidence regarding their exact location and medium specificity.It is possible to speculate that these transporters are involved in homeostasis of metals,especially Zn,Cu,and Fe.Most of the metals taken up by the fungus may be used to maintain fungal functioning,while surplus metals are transferred to the plant.In contrast,for excess metal ions in the cytoplasm,organisms have developed a different mechanism through which heavy metals are modified to bounded form and sequestered.Vacuoles are usually considered as store houses and detoxification sites for extra metal concentrations,while later mobilization is induced by enzymes such as in the case of Fe permease/oxidase complex mediating its mobilization on nutritional demand(Hijikataet al.,2010).Similarly,SMF3 proteins have been shown to transport Znto the cytosol from the vacuole and(in combination with CCC1)induce homeostasis inside the cell(Diffelset al.,2006).Considering this,metals are essential micronutrients,and all organisms compete for them when in short supply,activating transport systems that enable the uptake of metals required for their development.In contrast,excess levels of these heavy metals are toxic,and therefore suitable transport systems exist in mycorrhizae.Their transport is thus probably accomplished by transport systems for the essential elements Cu,Zn,Fe,and manganese(Mn),or even through phosphate transporters,as has been shown for the uptake of the metalloid arsenate that occursviathe high affinity phosphate transporter GiPT(Gonzlez-Chvezet al.,2011).
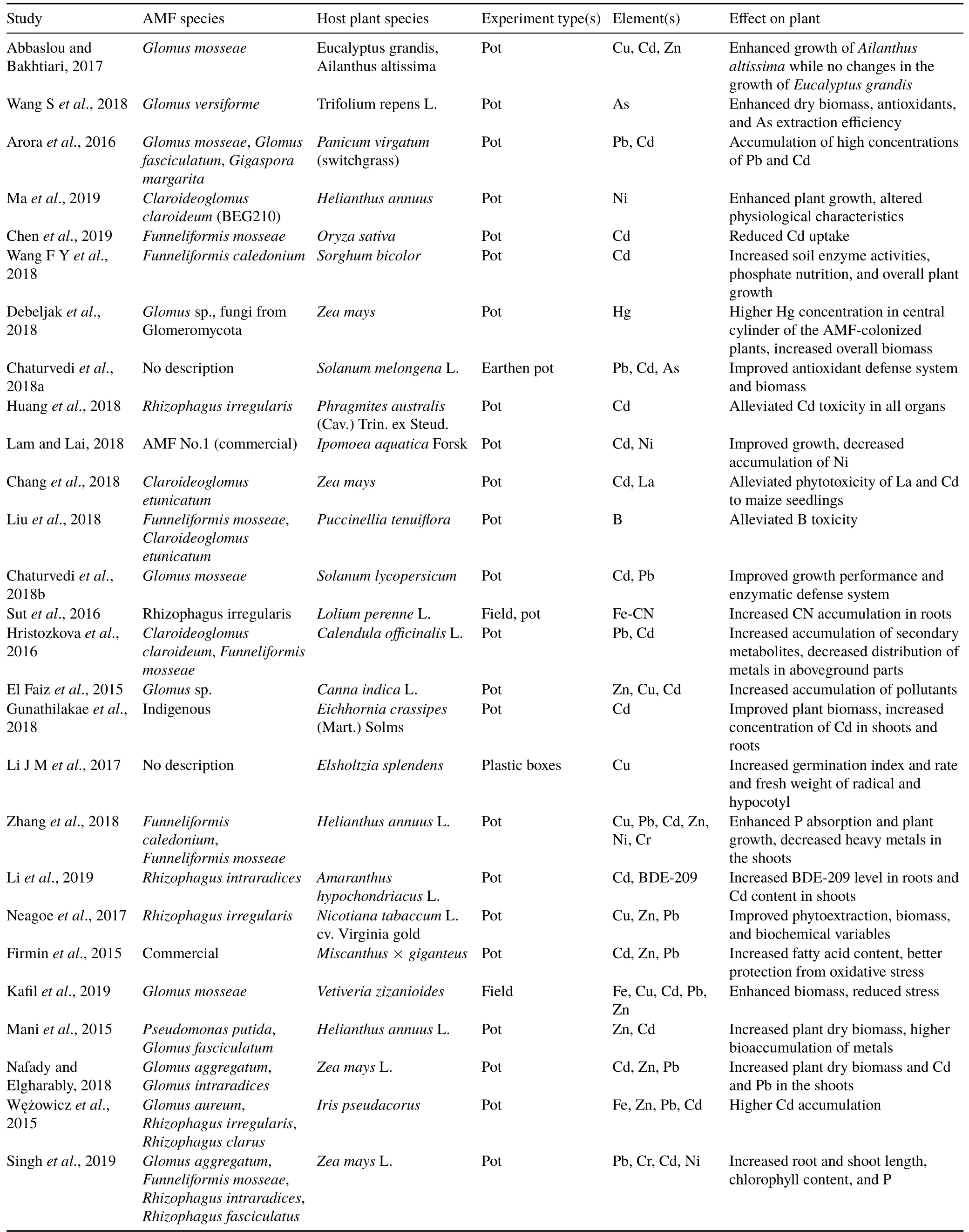
TABLE IStudies regarding contribution of arbuscular mycorrhizal fungi(AMF)to phytoremediation of pollutions caused by heavy metals and trace elements

TABLE I(continued)
ASSOCIATION OF AMF INTERFACES WITH PLANT TRANSPORTERS
Arbuscular mycorrhizae establish a direct link between soil and roots of the host plant,thus participating in the transport of materials,including metals,from soil to plant.Extraradical mycelium provides an uptake pathway for the different metals through its plasma membrane.An inducing effect on Cu and Zn genes has been observed inM.truncatularoots colonized by mycorrhizae(Kaldorfet al.,1999).Regulation of proteins related to Fe transport such as ferric chelate reductase and cell organelle-based Fe transporters,as well as Zn transporters,has been studied inSorghum bicolor(Handaet al.,2015).The metallothionein family set of genes has also been reported extensively in a wide range of different organisms that express them differentially upon exposure to metals such as Zn,Cu,or Cd.Distinct regulation of a metallothionein gene(BI451899)in extraradical mycelium associated withGlomus intraradiceshas been studied in the presence of Zn,while its role in Cu detoxification has also been reported(Lanfrancoet al.,2002).Genome-wide analysis of transcription patterns in laser-microdissected cortical cells colonized by arbuscules revealed that a Cu transporter was specifically expressed in arbuscule-containing cortical cells,suggesting that it may be involved in Cu acquisition by the periarbuscular membrane(Hogekampet al.,2011;Gaudeet al.,2012).However,some studies show that transport of metals is not restricted to arbuscule-colonized cortical cells.For example,the Zn and Mn transporter gene,MtZIP7,shows the same expression pattern in arbuscule-colonized and non-colonized cortical cells of plant roots.Another study revealed the genetic expression pattern of strategy II plants;AMF do not induce strategy II-related genes,but regulate other putative genes(OPT8aandOPT8b),performing a role in increasing Fe transport by more than 50-fold(Kobaeet al.,2014).Nicotianamines are also important metal chelators and the main building blocks for phytosiderophores synthesis.Their functions have been characterized in many model plants such as rice,barley,andArabidopsis.Studies have mainly identified their function in Fe transport at both intracellular and extracellular levels.Furthermore,it was suggested that they play a role in chelating the Fe supplied by mycorrhizae(Zhouet al.,2013).The study of expression patterns of 53 genes in the model plantArabidopsishas shown that Zn2+transporter(ZIP)genes,ZIP2andZIP4,are prominently involved in Fe transport(Wintzet al.,2003).Along with stress augmentation and transportrelated gene expression,evidence increasingly shows that symbiotic fungi contribute to the modulation of various biochemicals(antioxidative system)in plants.The mycorrhizal-induced antioxidative system(enzymatic and non-enzymatic antioxidants)in host plants can scavenge excess reactive oxygen species(ROS)produced by heavy metal exposure(Gill and Tuteja,2010;Emamverdianet al.,2015).There is overwhelming evidence that mycorrhizal association can augment antioxidant enzyme systems,including monodehydroascorbate reductase,dehydroascorbate reductase,and other enzymatic and nonenzymatic ROS scavenging systems(Vadasseryet al.,2009;White and Torres,2010).Recently,it has been shown that the activities of ascorbate peroxidases,glutathione peroxidase,and superoxide dismutase are elevated in arbuscular mycorrhizalRobinia pseudoacaciaL.,which probably contributes to the mitigation of Pb-induced toxicity(Yanget al.,2015).Similarly,modulation of stress-related enzymes in plants associated with AMF under As stress has been highlighted inL.leucocephala,emphasizing the potential ofL.leucocephala-AMF symbiosis for As phytostabilization in moderately As-contaminated soils(Schneideret al.,2017).In addition,non-enzymatic antioxidants also play key roles in ROS homeostasis in mycorrhizal plants exposed to heavy metals(Coninxet al.,2017).For instance,under higher Cu and Zn concentrations,Populus alba(AL35)pre-inoculated with AMF showed an overall upregulation of metallothionein(MT)and foliar polyamine(PA)biosynthetic genes,together with increased PA levels(Cicatelliet al.,2014).Although the exact mechanisms are still unclear and need to be elucidated,these biochemicals may act as free radical scavengers,stabilize membranes,and retard senescence(Haradaet al.,2010).
ARBUSCULAR MYCORRHIZAL FUNGI:INSIGHTS INTO DIVERSITYAT METAL-CONTAMINATED SITES
Associations of AMF with plant roots in heavy metalcontaminated soils have been widely recognized.In addition,they can play an important role in heavy metal tolerance,accumulation,and transportation from roots to aboveground plant parts(Karimiet al.,2011).Generally,plant roots(mostly from native species)at mine wastes are associated with AMF.For example,the evaluation of occurrence and diversity of AMF in areas with different As concentrations showed that the most frequent species occurring in all these areas wereParaglomus occultum,Acaulospora morrowiae,andGlomus clarum,indicating their high tolerance to excess As(Schneideret al.,2013b).Similarly,evaluation of the diversity of AMF and plant species,as well as their interactions,in soil contaminated with Pb found thatAcaulosporaandGlomusgenera exhibited the highest occurrence in both bulk and rhizosphere soils(Schneideret al.,2016).In another study,AMF diversity varied in the roots ofVeronica rechingeri,ranging from six to two types in unpolluted plots and plots with high heavy metal contamination,respectively(Zareiet al.,2008a).Diverse responses have been shown by AMF to heavy metals at the level of species,ecotype,or both.For instance,Glomus mosseae(isolated from heavy metal-contaminated soils)was found to be more tolerant to Cd stress than the same species isolated from uncontaminated substrate(Weissenhornet al.,1994).Similarly,Glomus etunicatumis more sensitive to Pb,Cd,and Zn thanG.intraradices(Pawlowska and Charvat,2004).Furthermore,Rhizophagus clarus(average hyphal diameter of 4 μm)absorbs more Cd,Cu,and Pb thanGlomus gigantea(average hyphal diameter of 13μm),indicating that absorption potential varies among different species of AMF(Cabralet al.,2015).It is important to understand how the AMF community reacts to heavy metal-stress,which can play crucial role in phytoremediation or re-vegetation of heavy metal-contaminated regions(Hildebrandtet al.,2007).It has been found that several plant species,such asSolidago giantea,Thlaspi praecox,V.rechingeri,Fragaria vesca,Thymus polytrichus,andV.calaminaria,colonized by AMF grow well in metal-contaminated areas,while their AMF isolates positively regulate the resistance of plants to heavy metal stress(Zareiet al.,2008b).The mycorrhizal effect of two strains of the AMFClaroideoglomus claroideum(S1 and S2),isolated from heavy metal-polluted soils,was investigated(Hristozkovaet al.,2015).The highest rate of colonization was shown by S1,while no significant differences were found between S2 and S3(F.mosseae,another strain isolated from an industrial area).Interestingly,AMF can affect the transformation of metals(especially trace metals)in the rhizosphere using various approaches,such as modification of root exudates,acidification,chemical precipitation in soil,and immobilization of heavy metals(Upadhyayaet al.,2010).Heavy metal-contaminated soils can be restored and re-vegetated using AMF that affect the rate of phytostabilization and phytoextraction processes(Ghre and Paszkowski,2006).
BENEFITS OF ENDOPHYTIC FUNGI IN PHYTOREMEDIATION
It has been reported that endophytes can play a significant role in host plant adaptation to different heavy metal-polluted environments.In addition,they can enhance phytoremediation by degrading,mobilizing,or immobilizing contaminants in soil,decreasing phytotoxicity,enhancing plant growth,and improving plants’metal tolerance(Zhanget al.,2011).Being suitable candidates for remediation,endophytic fungi can be useful in soils,used for long-term cropping.Besides promoting seedling growth,a significant increase in seed germination(69.8%)has been reported with application of endophytic fungi(Chen Yet al.,2013).Co-cultivation of watermelon seedlings withCeratobasidium stevensii(an endophytic fungus)degrades phenolic compounds and significantly increases stems growth and leaf length compared to uninoculated(control)plants(Xiaoet al.,2014).Similarly,in an aged petroleum-polluted soil,petroleum hydrocarbons were degraded by two grass species(Festuca pratensisandF.arundinacea)infected by endophytic fungi(Soleimaniet al.,2010a).Furthermore,phytoremediation efficiency can be improved by adopting a number of strategies including the management of microbial consortia:managing the beneficial effects of microbes on phytoremediator plants and their metal-modifying capabilities,along with the selection of certain microbes and engineering for metal tolerance.Certain fungi can reduce metals and provide resistance to metals,resulting in accumulations of gold,platinum,and silver in the form of nanoparticles in their intracellular spaces,while periplasmic and cytoplasmic hydrogenases play a crucial role in the metal transformation.Plants depend on a pseudoequilibrium between solid and aqueous soil phases for metal availability,but not on total metal content.The adsorption of metal to soil greatly restricts the metals available for plant uptake.Furthermore,soil components and their interactions with root exudates prevent or restrict the increase of water-soluble organometallic chelates in the rhizospheric region;consequently,the organic compounds(root exudates)can be rapidly sorbed to soil.The reduction in the labile pools of metals or water-soluble chelates is less than 10%of the total metal uptake by plants,suggesting that metals are generally acquired from less available pools(Durnet al.,2011).A number of transgenic species have also been used to enhance phytoextraction of metals,such as Cu,Cd,and Pb,and metalloids(selenium(Se),As)as they accumulate in the aerial parts of soil.Such approaches mainly involve the production of phytochelatins and metallothioneins(metal-detoxifying chelators),enhanced synthesis of enzymes involved in sulphur(S)metabolism,and implementation of metal transporters(Kotrbaet al.,2009).An endophytic fungus,Penicillium janthinellumLK5,minimized Cd-induced oxidative stress and membrane injury by reducing lipid peroxidation and electrolytes as well as increasing catalase activity and glutathione content in comparison with experimental control plants(Khanet al.,2014).Therefore,endophytes are suggested to counteract metal stress,which has drastic effect on plant growth and development.However,heavy metal accumulation,absorption,or tolerance by endophytes and their effects on host plant growth are not associated with the origin of endophyte(Shenet al.,2013).Intriguingly,endophytic microbes are not only resistant to heavy metals,but they are also capable of degrading organic contaminants by producing specific enzymes.Thus,remediation through these microorganisms(endophyte-assisted phytoremediation)is a promising technology forin-situremediation of multi-metal-polluted soils.Endophytes significantly enhance plant growth,enhance metabolism,and reduce phytotoxicity by augmenting metal translocation and accumulation during the phytoremediation process(Liet al.,2012b).They can interact efficiently with their host plants and more closely degrade heavy metals in contaminated soil than other microorganisms in the rhizosphere(Zhanget al.,2011).Host plant resistance to multi-metal contamination can be increased through endophytic fungi.For instance,
AspergillusG16 associated withBrassica junceaL.andTrichodermaH8 associated withAcacia auriculiformisimproved plant growth(44%and 167%,respectively)in Cdand nickel(Ni)-contaminated soils.Collectively,these two strains induced a greater plant yield(178%)compared to uninoculated plants in such contaminated soils(Jianget al.,2008).Another Zn-,Cd-,and Pb-resistant endophytic fungus(Lasiodiplodiasp.MXSF31),isolated from the stem ofPortulaca oleracea,increased the Cd extraction amount and enhanced canola biomass in soils contaminated with Pb and Cd(Denget al.,2014b).It has also been reported that certain endophytic yeasts can confer host plant resistance to different metals.InoculatingBrassica alboglabrawithCr yptococcussp.CBSB78 increased the extraction amounts of different metals such as Zn,Pb,and Cd in multi-metal-contaminated soils(Denget al.,2012).Studies showing contributions of endophytic fungi to the phytoremediation of different metals and trace elements are summarized in Table II.
ENDOPHYTIC FUNGI AND THEIR PHYSIOLOGICAL CHARACTERISTICS
Plant interactions with the microbial community aresymbiotic relationships that play an important role in structuring different plant communities and ecosystems(Mller and Krauss,2005).Root tissue transport systems of hyperaccumulator plants are correlated with excessive amounts of metalloid and metal accumulation.In addition,root exudates are thought to have an effect on the diversity of microbes(Weyenset al.,2009).Attention has been given in the recent past to the capacity of endophytes to promote plant growth,as well as the relationship between endophytes and microbes living in the rhizosphere.After getting inside plants through the roots(main site of endophytic colonization)and establishing themselves,endophytes confer resistance to insects and pathogens.Besides inducing heavy metal resistance in plant(Khan and Doty,2011),endophytes can also promote plant growth in different ways,such as producing growthregulating substances(cytokinins and indole acetic acid)and metabolizing the ethylene precursor 1-aminocyclopropane-1-carboxylic acid produced under stress(Glick,2003).TheCr yptococcusspecies CBSB78(endophytic yeast)possesses low 1-aminocyclopropane-1-carboxylic acid deaminase activity and is a low indole-3-acetic acid producer(Denget al.,2012).Furthermore,endophytic fungi boost plant fitness and other competitive abilities by enhancing growth rate and germination or by increasing the plants’absorption capacity for nutritional elements(Alyet al.,2011).Possession of a metal sequestration or chelation system and suitable degradation pathways by fungal endophytes increases host plant tolerance to heavy metal stress and assists the plants to survive in metal-contaminated soils(Alyet al.,2011).The fungal interaction ofTrichoderma atrovirideF6 withB.junceaL.significantly increases resistance to Ni2+and Cd2+and alleviates cellular toxicity.Additionally,plants inoculated with this F6 strain results in 170%,40%,and 110%increases in fresh weight in Cd-Ni-,Ni-,and Cdpolluted soils,respectively(Caoet al.,2008).Some yeasts,such asRhodotorula mucilaginosa,have been reported to promote plant growth by producing indole-3-pyruvic acid and indole-3-acetic acid.However,such effects have been scarcely reported in soils contaminated with metals(Xinet al.,2009).The tolerance mechanisms of endophytic fungi in toxic environments includes compartmentation,intracellular sequestration and complexation,extracellular metal precipitation and sequestration,volatilization,and binding of metal to the fungal cell walls(Fominaet al.,2005).

TABLE IIStudies showing contribution of endophytic fungi(EF)to phytoremediation of pollutions caused by different metals and trace elements
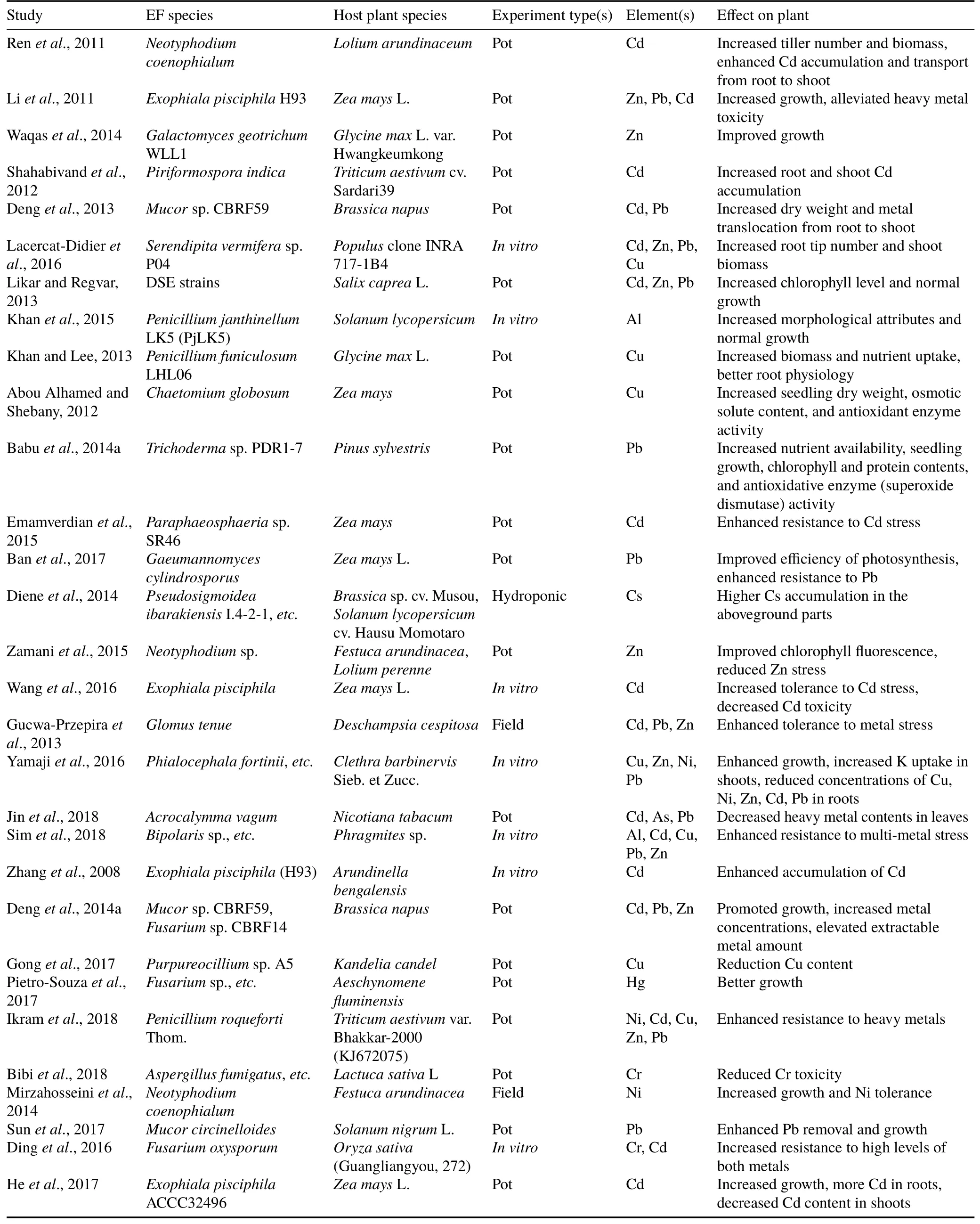
TABLE II(continued)
MECHANISMS BEHIND FUNGAL ENDOPHYTEDERIVED BENEFITS UNDER HEAVY METAL STRESS
Different molecular and cellular systems have been adopted by fungi living in metal-contaminated soils to cope and survive in such environments(Chiapelloet al.,2015).Detoxification mechanisms can be intracellular,such as binding to non-protein thiols and transporting into intracellular compartments(to reduce metal burden in the cytosol),or extracellular,such as avoidance of excessive metal entry,chelation,and cell wall binding(Ruytinxet al.,2013).All such processes contribute to the protection of fungal species from metal toxicity.Among these mechanisms,metal transporters are thought to be the most important contributors.They maintain suitable concentrations of metal ions in the cellular compartments in two ways:by allowing metal sequestration into intracellular compartments and by extruding toxic metal ions on inherent metal-tolerant ions from the cytosol to the outside of cell(Parket al.,2012).Various members of theNrampfamily have been reported to play a major role in homeostatic mechanisms,thereby preventing toxicity of metal ions in living cells(Ehrnstorferet al.,2014).Sub-cellular sequestration of various metal ions is mediated by diverse metal transporters,which not only control metal partitioning,but also contribute to cellular metal tolerance,especially when the rates of metal influx into organisms are high.Ametal transporter gene,homologous to the natural resistance associated macrophage protein(Nramp),was cloned fromExophiala pisciphila(DSE metal-tolerant fungi)and analyzed for functional characterization.The gene(Nramp,1716 bp)encodes a polypeptide of 571 amino acid residues.In addition,green fluorescence protein assay of EpNramp revealed it to be a plasma membrane metal transporter;this was confirmed by bioinformatics analysis with 11 transmembrane domains.Further experiments with yeast functional complementation showed that the growth defect of the Feuptake yeast mutant could be complemented by mediating the transport of Fe2+.Furthermore,Cd2+accumulation and sensitivity increased with the expression ofEpNramp.Expression ofEpNrampwas significantly downregulated with elevated Cd2+exposure,as revealed by quantitative polymerase chain reaction(qPCR)studies(Weiet al.,2016).Glutathione(GSH)and GSH-related tolerance are also one of the metal tolerance mechanisms in fungi,induced in response to various environmental factors(Na and Salt,2011).Endophytic fungi are capable of accumulating high quantities of toxic metals in their mycelia.However,the mechanisms of metal transport,sequestration,and detoxification are not well documented(Zahooret al.,2017).Transcriptome analysis of E.pisciphila(growing in the presence of Cd)using RNA sequencing(RNA-Seq)revealed that several genes associated with ROS,S assimilation,redox homeostasis maintenance,and metal binding,transport,and detoxification were upregulated.The researchers suggested that various molecules including MTs,GSH,and phytochelatins could increase under Cd stress(Na and Salt,2011).Another tripeptide GSH(γ-glu-cys-gly)frequently found in cell compartments has been shown to sequester metals and also acts as a potent antioxidant under oxidative stress(Freemanet al.,2004).A transient increase in GSH content after Pb treatment was recorded inGaeumannomyces cylindrosporusthat enhanced tolerance to Pb(Banet al.,2017).To date,there are no reports showing that endophytic fungi may affect metal translocation by interfering with the expression of metal transporters.However,an increased expression level ofpcr2(encoding plant cadmium resistance 2)results in higher metal accumulation in the shoots of inoculated plants(Rozp˛adeket al.,2018).In another study,an unspecifiedzipgene was inhibited with endophytic fungus inoculation,which coincided with the decreased uptake of Cd.Furthermore,mpt1(encoding metal transporter protein 1)expression was also upregulated in a group inoculated with endophytic fungi under Cd stress(Wanget al.,2016).In addition,several genes includingzif1(encoding zinc-induced facilitator 1),hma3(encoding heavy metal-associated 3),cax2(encoding cation exchanger 2),andmtp1that encode tonoplast-bound vacuole carrier protein were upregulated inArabidopsis arenosasubjected to metal stress.The authors suggested that these genes are involved in the reduction of metal accumulation,distribution,exclusion,sequestration in the vacuole and ROS scavenging(Rozp˛adeket al.,2018).The overexpression or complementation of these genes improves resistance to various metal stresses in yeast and plants(Morelet al.,2009).
ASSOCIATION OF ENDOPHYTIC FUNGI WITH PLANTS UNDER CONDITIONS OF METAL STRESS
The diversity of endophytic fungi is thought to be abundant,especially in tropical forests.However,their diversity in hyperaccumulators has not yet been well documented.It has been reported that endophytic fungi of plants at Pb-Zn sites,especially mine wastelands,harbor fewer endophytic fungi in their leaves than in their stems.For instance,Alternaria,Phoma,andPeyronellaeawere the dominant genera among twenty identified fungal taxa,some of which showed marked adaptation to Zn2+and Pb2+(Liet al.,2012a).In addition,Rhodotorulasp.CBSB79 andCr yptococcussp.CBSB78(endophytic yeasts)have been found in the roots ofBrassica chinensisin soils contaminated with Pb,Cu,Cd,and Zn.These species have shown resistance to various metals in multi-metal-contaminated soils(Wanget al.,2013).Various endophytic species in genera such asAspergillus,Trichoderma,Penicillium,Fusarium,Cladosporium,Lasiodiplodia,andPaecilomycesare harbored inPortulacaspp.,multiple heavy metal-hyperaccumulating plants(Denget al.,2014b).Similarly,A.auriculiformiscontains fungiPenicillium,Aspergillus,Alternaria,Fusarium,Trichoderma,andMycelia sterilia(Caoet al.,2008).Trichodermaspp.andPenicilliumspp.are famous and frequently isolated species that can resist heavy metal stress(Babuet al.,2014b;Khanet al.,2014).Conventional culture methods are not enough to isolate all fungi;however,the bias towards fast-growing ubiquitous species is the prominent shortcoming of cultivation-dependent studies for unraveling endophyte diversity.Consequently,rare species with more specialized requirements and minor competitive strengths may remain undiscovered.Methods that have been used for bacterial isolation do not favor fungal growth and other fungal colonizers in the same samples.Besides culture studies,advanced techniques such as barcoded amplicon pyrosequencing and next-generation sequencing can be utilized to analyze endophytic flora where diverse fungal species would be found in hyperaccumulator plants(Deng and Cao,2017).Heavy metal-resistant endophytes in contaminated areas have attributes that decrease metal phytotoxicity,enhance plant growth,and affect metal accumulation and translocation in plants(Liet al.,2012b).Different sensitivities to metals have been shown by endophytes isolated from different plant species or even from different tissues of the same plant.This suggests that isolates are different in metal tolerance due to long-term adaptation in plant tissues with different concentrations of heavy metals(Liet al.,2012a).Intriguingly,many metal-resistant endophytes have not only been isolated from hyperaccumulating plants such asAlnus firma,Nicotiana tabacum,Solanum nigrum,Thlaspi caerulescens,Brassica napus,Alyssum bertolonii,andThlaspi goesingense,but also from non-hyperaccumulating plants such asAcacia decurrens,Symplocos paniculata,andArabis hirsuta.These isolates from non-metal-polluted plants were also found to be resistant to heavy metals(Mooreet al.,2006).Another root-colonizing endophytic fungus with a wide number of hosts,Piriformospora indica,can colonize the roots of a number of plants growing on metal-contaminated soils and confer resistance against Cd,Cu,As,Zn,and Pb(Mohdet al.,2017).In addition,DSEs have been isolated from the roots ofErica herbaceagrown on Cd-,Pb-,and Znpolluted soils(Čevniket al.,2000).In another study,fungal endophyte communities ofSalix capreain locations highly enriched with Cd and Pb contained species belonging to the generaLeptodontidium,Phialophora,andPhialocephala.Furthermore,root colonization ofSalix capreaand the resulting frequency of the sequences have an affinity towardsPhialophora,showing correlation with available phosphate concentration and soil Pb and Cd,probably indicating that these endophytes confer tolerance to heavy metal contamination(Likar and Regvar,2009).Furthermore,the interactions of endophytic fungi with a number ofSalixspecies in heavy metal-polluted areas are well documented(Baumet al.,2006).
ECOLOGICAL AND ENVIRONMENTAL CONDITIONS AND THEIR EFFECTS ON POPULATION STRUCTURE OF ENDOPHYTIC FUNGI
Different environmental or ecological conditions such as humidity,temperature,and different levels of soil nutrition are amongst the significant factors that determine the amounts and types of secondary metabolites from endophytic fungi and host plants,which indirectly affect the population structure of endophytic fungi.If the environmental conditions comprise a high mean annual humidity with a low mean annual sunshine hours,host plants may produce an abundance of nutrients that might be suitable for reproduction,colonization,and dissemination of the endophytic fungi(Wuet al.,2013).In contrast,only a few types of host plants can grow successfully if environmental conditions are cold with unsuitable pH value,oxygen concentration,and respiration rate.Consequently,certain endophytic fungi(in a limited number)can form symbiotic relationships with corresponding host species,indicating a definite degree of regional specificity on the population structure of endophytic fungi(Jianget al.,2010).Generally,the population structures of endophytic fungi and their related host plants(usually the same species from different climatic regions)have shown a very low degree of similarity(Jianget al.,2010).However,a high degree of similarity has been shown in terms of endophytic fungus species taxonomy in the same regions,indicating a certain degree of regional specificity(D’Amicoet al.,2008).
PHYTOREMEDIATION
Some plants have the capability to retain high concentrations of heavy metals,while their normal development remains unaffected;using such plants for the remediation of heavy metals is known as phytoremediation.Phytoremediation is the most feasible option to decontaminate the serious pollution areas,especially when the harvested biomass is valorized to produce bioenergy,as in the production of biodiesel.Such plants can maintain homeostasis while growing in highly polluted soil medium and can be used to degrade,transfer,or remove heavy metal contaminants(Wong,2003).A significant number of plants have been reported for their capability to hyperaccumulate metals in their tissues.Approximately 500 plant species,belonging to 45 families,have been extensively reported on and recognized as having high potential for metal-accumulating capabilities.Among these plants,members of Euphorbiaceae,Fabaceae,Lamiaceae,Brassicaceae,Scrophulariaceae,and Asteraceae are the most prominent(Ghosh and Singh,2005;Jaffret al.,2013).Eleocharis acicularis,A.bertolonii,andT.caerulescensare some of the highly effective examples studied for their accumulation of Ni,Cd,and Zn(Lasat,2002;Assunoet al.,2003;Glick,2010),andT.caerulescenshas been shown to accumulate Cd of 0.3—1 020 mg kg-1and Zn of 500—52 000 mg kg-1(Zhaoet al.,2003).
Phytoremediation approaches involve various plantbased strategies with different mechanisms and actions,the most common of which are phytostabilization,phytofiltration,phytoextraction,and phytodegradation or phytotransformation.Phytostabilization is the reduction of heavy metal bioavailability in soil by plants(Saltet al.,1995).Some plants gain metal resistance capabilities through gradual evolution or get them inherently(Wu,1990).Plants used for phytostabilization are selected based on their resistance to certain contaminants,along with their capability of holding the contaminant in roots and hampering its distribution towards the upper parts,avoiding its entrance to the food chain(Padmavathiamma and Li,2007).Recently,Agrostis castellanahas been reported to be a good candidate for accumulating high concentrations of pollutant elements including Zn,Cu,Pb,Cd,and As.Analysis of its below-and aboveground biomass have shown that it accumulates most of the pollutant elements in the roots,proving that this plant is a good phytostabilizer(Pastoret al.,2015).In another study,the phytostabilization potential ofGentiana pennellianahas been shown at Zn-,Cu-,and Pb-contaminated sites(Yoonet al.,2006).
Phytofiltration implies the removal of contaminants from water sources(wastewater)by utilizing different plants to purify the aquatic ecosystem(Mesjasz-Przybyłowiczet al.,2004).For example,removal of Cr,Cd,Ni,and Cu has been reported in the wetland plantEichhornia crassipes,showing that it is a promising candidate for cleaning polluted wastewater.However,in rhizofiltration,terrestrial plants are preferred because of their extensive root system,which provides a wide surface area.Another reason for this preference might be the high water storage contents of aquatic plants,making later composting,drying,and incineration difficult.Ricinus communisL.has been shown to accumulate higher Cd in its root system compared to shoots and leaves grown in hydroponic solution(Hadiet al.,2015).Similarly,B.junceahas shown maximum accumulation of Cr,Cu,Zn,Cd,Ni,and Pb from hydroponic solution(Dushenkovet al.,1997).
Phytoextraction,phytoaccumulation,and phytoabsorption are the most highly used techniques for the uptake of heavy metals in vast quantities by plants and shifting them to their harvestable parts or components(Maet al.,2011;Kamranet al.,2014).After harvesting,the metalhyperaccumulated harvested biomass of plants can be processed to extract and recover the possible pollutant or contaminant(Haet al.,2011).Phytomining technology employs hyperaccumulator plants to take up metal in harvestable plant biomass.Harvesting,drying,and incineration of the biomass generate a high-grade bio-ore.Thus,“agromining”(a variant of phytomining)could provide local communities with an alternative type of agriculture on degraded lands:farming not for food crops,but for metals.However,phytoextraction of some elements to remediate soils results in recovered biomass with little economic value(for example Cd and As),and disposal of the biomass would be costly.A few elements such as Ni,cobalt(Co),and uranium(Au)may have sufficient economic values for phytomining biomass to support commercialization.The development of phytomining requires:i)selection of high-biomass hyperaccumulator plant species,ii)evaluation of genetic diversity and breeding of improved strains with higher yields of the phytoextracted element,iii)development of agronomic practices to maximize economic return,and iv)development of methods to recover the phytomined element from the plant biomass.Full commercial phytomining of Ni is commencing in Albania usingAlyssum murale,while major trials in Malaysia are underway usingPhyllanthus securinegioides.The variable prices of commodity metals add confusion to the development of commercial phytomining(van der Entet al.,2015).
Many flowering plants(500 species)have been reported with the potential to accumulate various metals(Cd,Co,Cu,As,Ni,Mn,stibium(Sb),Pb,thallium(Tl),Zn,and Se),and scientists are still in search of new plant species with hyperaccumulation capabilities(Padmavathiamma and Li,2007;Rascio and Navari-Izzo,2011;Jaffret al.,2013).Some recent examples includeT.goesingense,Sedum alfredii,andArabidopsis halleri,which are hyperaccumulators of Zn.Nicotiana tabacumL.has been shown to have potential for Cd hyperaccumulation,whileS.alfredii,T.caerulescens,Thymus praecox,andA.hallerihave been shown to have both Zn and Cd hyperaccumulation potential(Rascio and Navari-Izzo,2011;Yanget al.,2017).Another common mechanism is phytodegradation or phytotransformation involving the external degradation of pollutantviaexcreted metabolic compounds or breaking down the pollutant after being taken in by plant tissues through various metabolic processes(Tangahuet al.,2011).The endophytic or rhizospheric microbes in association with plants have also been extensively reported to assisting in the biodegradation of contaminants(Tangahuet al.,2011;Bibiet al.,2018).For instance,the endophytic strainsAspergillus fumigatusML43 andPenicillium radicumPL17 have been shown to biotransform Cr from its hexavalent form to its trivalent form(less toxic),while maintaining healthy growth and development of the host plantLactuca sativa(Bibiet al.,2018).Similarly,Microbacteriumsp.G16 andPseudomonas fluorescensG10 have been reported to produce organic acids and increase the accumulation of Pb in rape plants(Shenget al.,2008).
CONCLUSIONS AND FUTURE PROSPECTS
Soils contaminated with toxic heavy metals not only have drastic effects on the diversity and composition of microbial communities,which play a significant role in the reduction of important microbial species,but also result in the enrichment of metal-tolerant or resistant microbial strains.Interestingly,these microbial strains(fungi,bacteria,algae,and other microorganisms)are not only metal tolerant,but also promote plant growth on soils contaminated with the toxic metals.In the recent past,the mycorrhizal effects on plants growing under metal stress conditions and plant adaptations to these metals have been extensively studied.Based on the recent research articles discussed here,fungal-assisted(particularly AMF and endophytic fungi)phytoremediation is the most suitable and ecofriendly technology for the remediation of soils polluted with heavy metals.Fungal interactions with plants at metal-contaminated sites can enhance plant growth,mobilize heavy metals,degrade organic contaminants,improve nutrient uptake,promote seed germination,significantly affect contaminant accumulation,and decrease phytotoxicity.Besides other important factors,this review article highlights fungal-plant interactions,particularly in metal-polluted soils,as well as the benefits that these fungi provide to their host plants.Although worldwide attention has been given to the removal of heavy metals and pollutants from the environment(water,soil,and sediment),there are many areas which remain poorly understood and require additional research.For example,much of the reported work is confined to experimental studies;more effort should be made into field trials.Additionally,fungal-plant interaction mechanisms have not been clearly explained.Besides a bioengineering system to understand antagonistic or mutualistic symbiosis of fungi with their host plants,a rapid screening guide is necessary for beneficial fungi rather than isolating the whole microflora uncritically.Being a dynamically growing area of green biotechnology,bioremediation(despite a few disadvantages)is a cost-effective and environmentally friendly(without any hazardous effects)technology;however,the various pending questions should be addressed.For instance,how much the analogous impacts of mycorrhizal fungi differ from the influences of endophytes on plant toxic metal tolerance?Do fungi adapted to heavy metal stress directly affect specific metal tolerance mechanisms,or is this a result of indirect action such as improving the uptake of water and nutrients from the soil and conferring plant fitness?Thus,a greater understanding of the mechanisms behind metal tolerance in beneficial fungi will provide essential tools for their efficient use in phytoremediation practices.Additional studies regarding the role of fungal symbionts in plant responses to toxic metals should be carried out to answer these and other important questions.
ACKNOWLEDGEMENTS
This work was supported by the National Key Research and Development Program of China(No.2016YFD-0800807),the National Nature Science Foundation of China(Nos.31702003 and 31902105),the Young Elite Scientists Sponsorship Program of China Association for Science and Technology(CAST)(No.2017QNRC001),the China Postdoctoral Science Foundation(No.2019 M651505),the“Chenguang Program”of the Shanghai Education Development Foundation and Shanghai Municipal Education Commission,China(No.17CG07),and the Shanghai Agriculture Science and Technology Development Project,China(No.2018-4-13).
杂志排行

Pedosphere的其它文章
- Preface Earthworms in soil ecology and organic waste management
- Letter to the Editor Field and laboratory investigations of Lumbricus badensis ecology and behaviour
- Strategies to mitigate the adverse effect of drought stress on crop plants—influences of soil bacteria:A review
- Study of oxidative stress cadmium(Cd)-induced in Eisenia fetida based on mathematical modeling
- Behavior and respiration responses of the earthworm Eisenia fetida to soil arsenite pollution
- Biological weathering of phlogopite during enriched vermicomposting