高粱×苏丹草杂交后代产量性状的遗传分析
2021-07-14谭燕卢华雨康晨孙守钧罗峰
谭燕,卢华雨,康晨,孙守钧,罗峰
高粱×苏丹草杂交后代产量性状的遗传分析
谭燕,卢华雨,康晨,孙守钧通信作者,罗峰通信作者
(天津农学院 农学与资源环境学院,天津 300392)
以国产高粱品种‘忻粱52’和苏丹草品系‘美引-48’杂交所获得的F2代群体为试验材料,对单株粒重、着壳率、千粒重和穗重4个产量性状进行测定和分析,并采用主基因+多基因混合遗传模型法进行遗传分析,以确定各性状的最适遗传模型,计算其主基因遗传率,分析各性状的遗传规律。结果表明:model B_2是单株粒重的最适模型,主基因遗传率为76.38%;着壳率、千粒重和穗重这3个性状的最适模型均为model B_1,即受两对主基因控制的加性-显性-上位性混合遗传模型,其主基因遗传率分别为61.25%、52.85%、49.81%。在这4个性状中,单株粒重的遗传率最高,表明该性状向后代稳定遗传的能力较强;但穗重的遗传率较低,表明穗重受环境因素的影响较大,向后代稳定遗传的能力较弱。因此,以穗重性状进行高梁选育应在高世代进行。
高粱;产量性状;遗传分析
高粱和苏丹草为禾本科高粱属中两个不同的种,虽然它们之间具有一定的形态差异但生殖隔离并不显著,因此可以进行杂交育种[1]。培育高产高粱杂交种一直是主要育种目标。二者作为父母本,培育双亲优良性状结合的杂交种被称为高丹草[2]。然而,由于多数产量性状属于数量性状,会受到多个性状的综合影响,每个性状相互关联和相互制约[3]。因此,培育高产量和高质量兼具的优良高粱品种,需要着重考虑单株粒重、千粒重和穗重这3个对提高高粱产量起着十分重要作用的性状,它们不仅是衡量产量的重要指标,在产量上也有着很高的协同效应。此外,虽然着壳率不是一个经济性状,对生产的影响很小,但在实际研究中发现,着壳率与高粱品质密切相关。为保证高粱的品质,较低的着壳率也是一个好品种需要具备的特点[4]。国内外许多学者通过对不同高粱品种的研究表明,产量性状加性效应和显性效应都会对产量产生重要影响[5]。
数量性状间的表型和内在遗传联系的问题,对于许多以加性效应为主的遗传性状,也存在显性效应,以保证后代正常的遗传[6]。而此类研究并不适用于所有区域,不同品种与栽培条件等都有可能对最终研究结果产生不同的影响[7-9]。试验类型、栽培地域、气候条件、试验地环境状况的差异等都会对试验结果和最终产量造成影响[10-11]。
本研究以山西高粱‘忻粱52’和苏丹草品系‘美引-48’为亲本,通过杂交、自交得到F2代群体。分别对单株粒重、着壳率、千粒重和穗重进行测定和遗传分析,根据主基因+多基因遗传分析方法,得出各性状的极大似然值、值及基因各成分的分布,确定出最适合的遗传模型。同时,进行相应的遗传参数估计,计算出主基因遗传率,整理分析各性状的遗传规律,为进一步杂交和育种研究奠定基础。
1 材料与方法
1.1 供试材料
供试材料包括杂交亲本国产高粱‘忻粱52’和苏丹草‘美引-48’群体、‘忻粱52’和‘美引- 48’杂交获得F1代群体,以及F1代自交获得的F2代群体。
1.2 试验设计
2016年12月,在海南省乐东黎族自治县利国镇官村将‘忻粱52’和苏丹草品系‘美引-48’进行杂交,获得杂交种F1代。2018年5月,在天津市宁河区实验林场将亲本及F1代进行播种,通过正反交比较排除细胞质遗传,将F1套袋使其自花授粉以获得F2代杂交种。2019年5月,播种亲本各1行、F1代1行、F2代50行,对F2代单株套袋,自花授粉。行长5 m,行距0.5 m,株距0.25 m,田间管理同大田生产。在完熟期取样测定。
1.3 测定方法
在F2代每行中随机取样,共取400株植株进行测定。F2代群体中需要测定的单株性状包括:
(1)单株粒重(g):将主穗完整脱粒,对全部籽粒进行称重,3次重复,取平均值。
(2)着壳率(%):随机选取1 000粒籽粒,数取具有颖壳的籽粒数,计算着壳率。
(3)千粒重(g):主穗脱粒后,随机选取1 000粒称重,3次重复,取平均值。
(4)穗重(g):在完熟期,从高粱穗基部位置剪下主穗,称重。
遗传率通过主基因+多基因遗传分析模型软件进行分析得出。
1.4 数据处理
使用Microsoft Excel2010和SPSS Statistics19.0对试验数据进行分析,然后根据盖钧镒[12-13]提出的主基因+多基因遗传分析模型方法进行基因遗传分析,结合方法中的值最小原则,筛选确定各性状的最适遗传模型,并计算主基因遗传率,分析各性状的遗传规律。
2 结果与分析
2.1 高粱×苏丹草杂交F2群体各产量性状相关性分析
单株粒重、千粒重和穗重对高粱产量有重要影响。从表1中可以看出,单株粒重与着壳率呈显著正相关,与千粒重、穗重均呈极显著正相关;着壳率与穗重呈显著正相关;千粒重与穗重呈极显著正相关。

表1 F2群体产量性状相关性分析
注: *表示在0.05水平上显著相关,**表示在0.01水平上显著相关。=400
2.2 高粱×苏丹草杂交F2群体各产量性状统计分析
由‘忻粱52’和‘美引-48’杂交得到的F2代群体的产量性状统计分析表(表2)和F2代群体产量性状频数分布图(图1)可以看出,单株粒重的极大值和极小值分别为0.09和41.98,说明F2代群体中个体间单株粒重的差异较大。同理,着壳率、千粒重和穗重的个体间差异也较为明显。单株粒重、着壳率、千粒重和穗重这4个产量性状的频数分布连续且呈正态(偏正态)分布,与数量性状遗传特征一致。因此,单株粒重、着壳率、千粒重、穗重这4个性状均属于数量性状。

表2 F2群体产量性状统计分析

图1 F2群体产量性状频数分布
2.3 遗传模型检验
采用12、22、32(均匀性检验)、2(Smirnov检验)、(Kolmogorov检验)对单株粒重、着壳率、千粒重和穗重这4个产量性状的备选模型进行适合性检验。结合表3,选出粒重、着壳率、千粒重、穗重这4个产量性状的备选模型(表4)。根据最小值原则,在所选的4个备选模型中选择各检验方法统计量达到显著水平最多的模型作为该性状的最适模型。
在Smirnov检验和Kolmogorov检验中,单株粒重和着壳率的4个备选模型均未达到显著水平(<0.05)。在均匀性检验中单株粒重model B_2模型均达到显著水平,且model B_2的值最小,因此将model B_2模型确定为该群体单株粒重性状的最适遗传模型;着壳率在均匀性检验32中model B_1模型达到显著水平,在12和22中model B_2模型达到显著水平,其中model B_1模型的值最小,因此将model B_1模型作为该群体着壳率的最适遗传模型。说明单株粒重与着壳率的最适模型均为受2对主基因控制的加性-显性-上位性混合遗传模型。
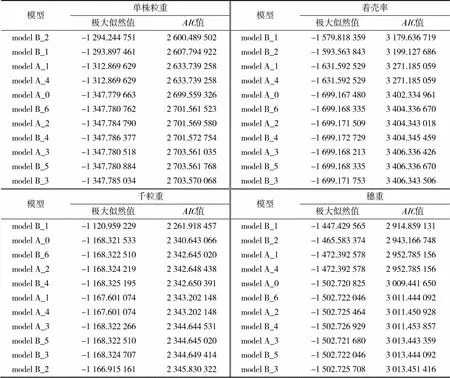
表3 高粱单株粒重、着壳率、千粒重、穗重遗传模型的极大似然值和AIC值
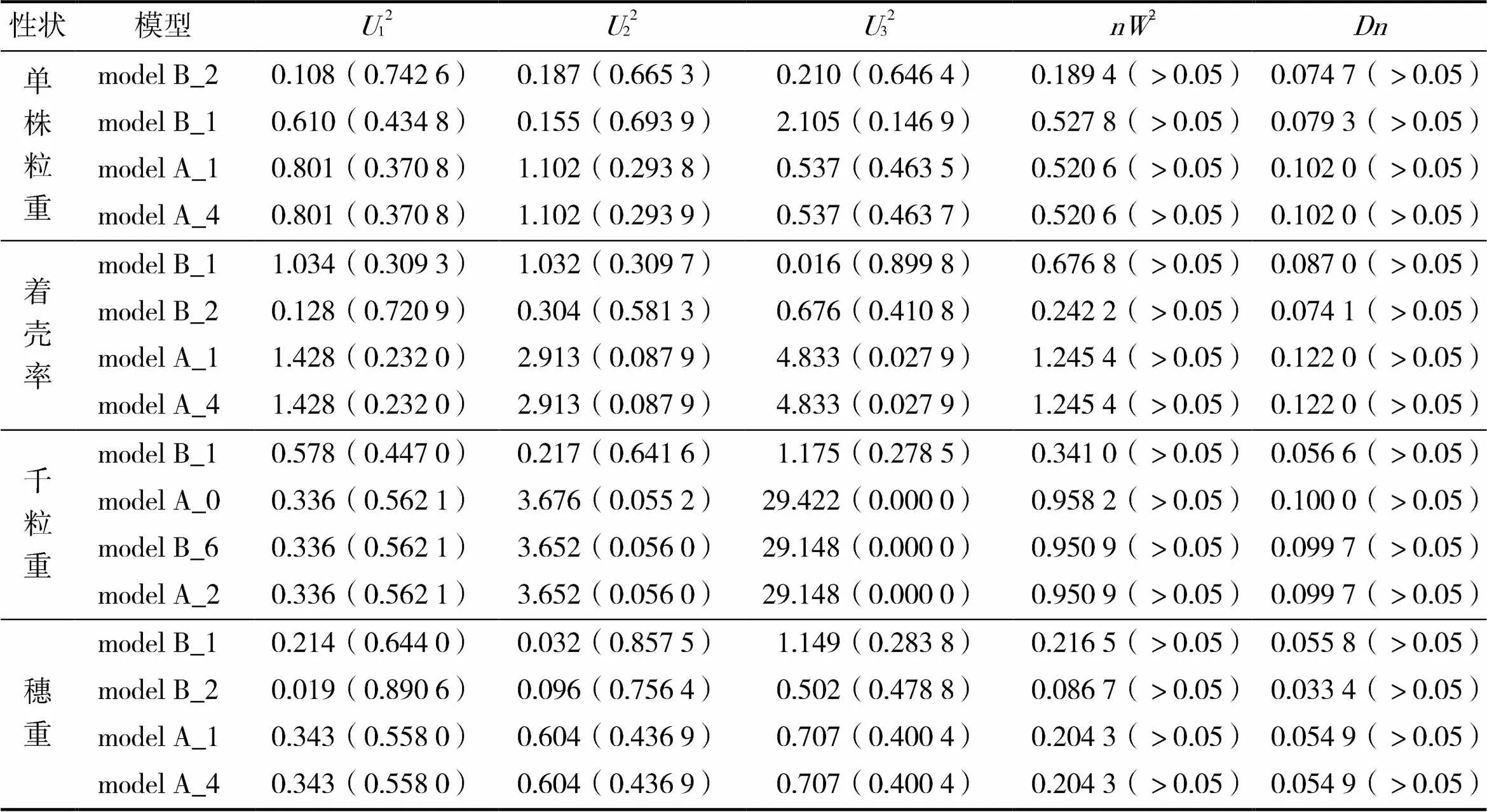
表4 高粱各产量性状备选模型的适合性检验
在千粒重的4个备选模型中,Smirnov检验和Kolmogorov检验均未达到显著水平,在均匀性检验12中model A_0、model A_2、model B_6模型均达显著水平,在22中model B_1模型达到显著水平,而最适备选模型中model B_1模型的值最小,因此确定model B_1模型作为该群体千粒重性状的最适遗传模型。
在穗重的4个备选模型中,Smirnov检验均未达到显著水平,而Kolmogorov检验中model B_2模型达到显著水平(<0.05),在均匀性检验12中4个备选模型均达到显著水平,在22中model B_1、model B_2模型达到显著水平,其中model B_1模型的值最小,确定模型model B_1为该群体穗重的最适遗传模型。
2.4 遗传参数估计
本试验群体中单株粒重的最适混合遗传模型为model B_2,是加性-显性-上位性混合遗传模型的最佳模型。根据已确定的最适模型和盖钧镒的IECM估算方法,对F2代群体单株粒重、着壳率、千粒重、穗重4个性状的遗传参数进行计算。
由表5可知,单株粒重的平均值为16.163 62,控制单株粒重的2对主基因的加性效应之和为12.414 23,显性效应之和为-12.414 18,加性效应显著大于显性效应,表明控制高粱单株粒重性状的2对主基因为加性效应。该性状主基因遗传率较高,为76.38%,说明环境因素对高粱单株粒重的影响较小。
着壳率、千粒重和穗重均符合model B_1模型,F2代群体的分离符合加性-显性-上位性的混合遗传模型。着壳率、千粒重、穗重的平均值分别为 25.836 50、23.772 39和21.623 52;主基因遗传率分别为61.25%、52.85%和49.81%。其中穗重的遗传率最小,说明在该群体中穗重这一性状早期选择效果差,后代遗传易受到环境因素的影响而不稳定。
控制着壳率的2对主基因的加性效应之和为31.773 31,显性效应之和为-16.976 88;控制千粒重的2对主基因的加性效应之和为11.811 36,显性效应之和为-11.641 72;控制穗重的2对主基因的加性效应之和为18.108 77,显性效应之和为-9.329 19。由此可以看出,着壳率、千粒重和穗重3个性状的加性效应显著大于显性效应,表明控制该群体着壳率、千粒重和穗重的主基因以加性效应为主,显性效应不明显。
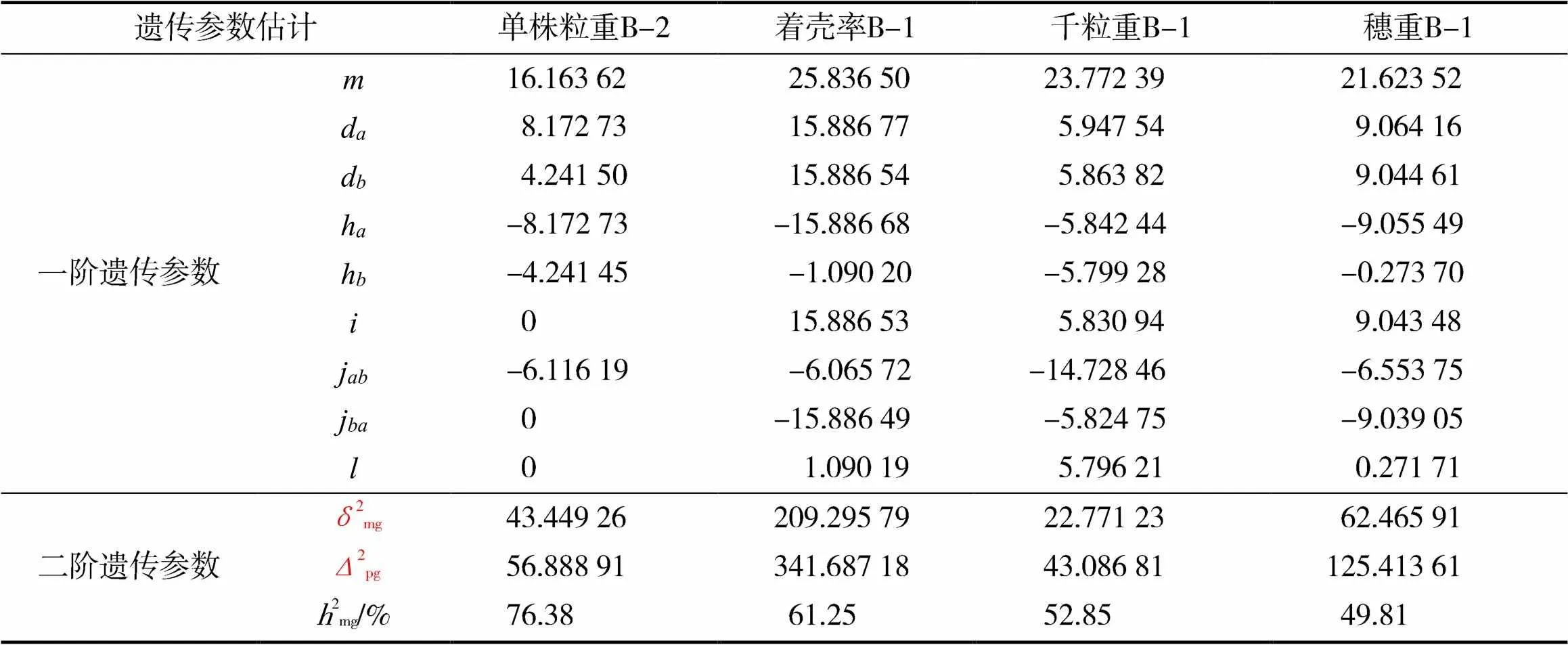
表5 高粱各产量性状遗传参数估计
注:为群体均方;d为主基因的加性效应;d为主基因的加性效应;h为主基因的显性效应;h为主基因的显性效应;为2个主基因的加×加效应;j为加性()×显性();j为加性()×显性();为2个主基因的显×显效应;2mg为遗传方差;2pg为表型方差;2mg为遗传率
3 讨论与结论
本研究采用主基因+多基因遗传分析法,对单株粒重、着壳率、千粒重和穗重进行遗传分析。结果表明,单株粒重的主基因遗传率为76.38%,由2对主基因控制以加性效应为主,其加性遗传率比较高,可在后代中稳定遗传。这与杨伟光等[14]的研究结果不一致,其研究认为高粱穗粒重既有明显的显性效应,又有不容忽视的上位性效应。原因可能是供试材料不同,试验类型、亲本选择、栽培地域、气候条件、试验地环境状况的差异等都会对试验结果造成影响[15]。着壳率的主基因遗传率为61.25%,着壳率最适模型可以被认为受2对主基因控制的加性-显性-上位性的混合遗传模型;千粒重符合model B_1模型,主基因遗传率为52.85%,且以加性效应为主。杨伟光等[16]研究中国地方高粱千粒重的加显性遗传模型和规律认为,千粒重的遗传模型符合加性-显性,其中加性效应更重要。这与本研究对高粱千粒重的研究结果一致。穗重的最适模型为model B_1,受2对主基因控制,且F2群体的分离符合加性-显性的混合遗传模型,主基因遗传率为49.81%,遗传率相对较低。这与于澎湃等[17]研究不一致,其研究认为高粱穗重最适模型为B_2模型,说明穗重在后代遗传稳定性上受环境因素影响较大。现已有研究表明,土壤条件、速效磷含量、日均温度、昼夜温差是影响穗重的主要因素[9]。高粱产量性状受多种因素影响,例如栽培条件和环境因素等。本试验中采用的高粱品种具有较高的单株粒重遗传率,故该性状受环境影响较小,而低遗传率的穗重则受环境因素的影响较大。因此,需要控制穗重在环境中所受到的影响,才能达到提高产量的目的。
[1] 胡春花,徐红军,冯浔. 高粱品种数量性状的遗传及育种目标探讨[J]. 新疆农垦科技,1995(4):26-27.
[2] 杨小翠,詹秋文,刘言龙,等. 苏丹草与高粱的判别分析和聚类分析[J]. 中国草地学报,2016,38(1):41-46.
[3] 包红霞,张春华. 高粱主要数量性状遗传分析[J]. 内蒙古农业科技,1997(2):11-12.
[4] 李树华,张文杰,白海波,等. 春小麦穗部性状的主基因+多基因遗传分析[J]. 中国农学通报,2017,33(6):20-26.
[5] MADHUSUDHANA R,PATIL J V. A major QTL for plant height is linked with bloom locus in sorghum [(L.)Moench][J]. Euphytica,2013,191(2):259-268.
[6] 游录鹏. 高粱遗传连锁图谱的构建及重要农艺性状QTL初步定位[D]. 南京:南京大学,2013.
[7] 白晓倩,卢华雨,于澎湃,等. 粒用高粱×苏丹草杂交F2代农艺性状的数量遗传分析[J]. 江苏农业科学,2019,47(19):188-193.
[8] 刘静,曹雄,李婷,等. 不同高粱品种主要农艺性状及产量的相关分析[J]. 安徽农业科学,2019,47(20):29-30,36.
[9] 罗峰,李欣禹,唐朝臣,等. 影响甜高粱主要农艺和品质性状的环境因子剖析[J]. 中国农业大学学报,2019,24(10):10-17.
[10] 詹鹏杰,平俊爱,楚建强,等. 不同种植环境和密度对机械化栽培高粱农艺性状及产量的影响[J]. 山西农业科学,2019,47(10):1783-1788,1814.
[11] 袁苗苗,张烈,曹高燚,等. 磷肥对玉米产量及青饲品质的影响[J]. 天津农学院学报,2020,27(2):18- 22,27.
[12] 盖钧镒. 植物数量性状遗传体系的分离分析方法研 究[J]. 遗传,2005,27(1):130-136.
[13] 盖钧镒,章元明,王建康. QTL混合遗传模型扩展至2对主基因+多基因时的多世代联合分析[J]. 作物学报,2000,26(4):385-391.
[13] 杨珍,李斌,赵军,等. 甜高粱主要农艺性状与产量相关和通径分析[J]. 中国糖料,2018,40(4):16-19.
[14] 杨伟光,李殿申,魏晓明,等. 高粱产量构成因素的遗传模型测验[J]. 吉林农业大学学报,1995,17(4):1-6.
[15] MI X J,ESKRIDGE K,WANG D,et al. Regression-based multi-trait QTL mapping using a structural equation model[J].Stat Appl Genet Mol,2011,9(1):17-20.
[16] 杨伟光,王奇,韩立军,等. 中国高粱千粒重的遗传模型测验[J]. 吉林农业大学学报,2001,23(2):1-4.
[17] 于澎湃,卢华雨,谭燕,等. 高粱主要穗部性状遗传分析[J]. 现代农业科技,2020(4):6,8.
Genetic analysis of yield traits in the hybrid progeny of(Piper) Stapf
Tan Yan, Lu Huayu, Kang Chen, Sun ShoujunCorresponding Author, Luo FengCorresponding Author
(College of Agronomy and Resource Environment, Tianjin Agricultural University, Tianjin 300392, China)
In this experiment, the F2population obtained from the cross between the domesticvariety ‘Xinliang 52’ and the(Piper) Stapf line ‘Meiyin-48’ was used as the test material to determine and analyze the four yield traits of single plant grain weight, hulling rate, 1 000-grain weight and ear weight, mixture model of major gene plus polygene was used for genetic analysis to determine the most suitable genetic model for each trait, calculate the heritability of the major gene, and analyze the genetic law of each trait. The test results showed that model B_2 was the most suitable model for single-plant kernel weight, with a major gene heritability rate of 76.38%; the best models for the three traits of hulling rate, thousand-grain weight and ear weight were all model B_1, which was affected by two pairs of major genes. In the controlled additive-dominant-epistatic mixed inheritance model, their major gene heritability rates were 61.25%, 52.85% and 49.81%, respectively. Among these four traits, the heritability of grain weight per plant is the highest, indicating that the trait has a strong ability to be inherited stably to offspring, but the heritability of ear weight was low, indicating that ear weight is more affected by environmental factors, and that the ability of stable inheritance is weak. Therefore, selection ofwith ear weight traits should be carried out in the higher generation.
; yield traits; genetic analysis
S514
A
1008-5394(2021)02-0006-06
10.19640/j.cnki.jtau.2021.02.002
2020-10-19
国家公益性行业(农业)科研专项项目(201503134)
谭燕(1994—),女,硕士在读,主要从事饲用作物遗传改良研究。E-mail:529283778@qq.com。
孙守钧(1961—),男,教授,博士,主要从事饲用作物遗传改良研究。E-mail:sunshoujun@tjau.edu.cn。
罗峰(1980—),男,副研究员,硕士,主要从事高粱遗传育种研究。E-mail:luofeng1868@126.com。
责任编辑:宗淑萍
