基于活立木茎干含水量的杨树生长状态评估模型构建*
2021-07-13刘卫平维高赵燕东
刘卫平 宋 维高 超 赵燕东
(1.北京林业大学工学院 北京 100083; 2.北京工商大学计算机与信息工程学院 北京 100048; 3.北京林业大学城乡生态环境北京实验室 北京 100083)
依据人的心跳、血压、呼吸速率等指标,可评判人体生命体征,分析人的生理健康状态,并基于这一原理开发出了很多操作简单、使用可靠、广泛推广的实时监测设备(Levy, 2004),促进了人类健康的自我诊断。植物同样具有呼吸、蒸腾、光合作用等一系列生命活动和体征指标。植物中的水分参与各项生命活动,为更深入理解植物的生命活动,国内外众多学者对植物中的水分开展了大量研究,如利用茎流计测量树干液流(Molinaetal., 2019;Tieetal., 2017; Sevantoetal., 2011)、在水中加入同位素观测水在树体内的流向(李雪松等, 2018)、利用核磁共振获得植物体内水分成像(Byrenetal., 1986)等。但同位素观测和核磁共振成像方法均存在较严重的破坏性、投入过高、操作繁琐等问题,茎流计因利用温度差进行测量,存在滞后性、受环境温度影响、对活立木产生损伤缺点,且茎流计本身造价高昂难以普及。由于获取方法匮乏,造成衡量生命活动的参数采集困难,数据量不足,从而使得这些植物生理参数仍停留在科学研究上,极少应用于实际生产。基于茎干含水量的测量方法,可弥补液流测量的不足,轴向上茎干含水量长距离运输产生液流, 径向上茎干中导管水分与储存组织细胞之间的水分交换影响茎干含水率(赵燕东等, 2016; Zhouetal., 2018; Beedlowetal., 2017)。茎干含水量传感器可实时、精准、快速地采集植物茎干含水量数据(赵燕东等, 2016),对茎干含水量数据的分析可揭示茎干含水量变化规律与植物生理状态变化的关系。研究者不断挖掘出不同生物与环境之间的相互作用规律(顾润源等, 2012; Inoueetal., 2015; Jinetal., 2017),环境因素直接影响植物生理状态,并能反映到茎干含水量变化上; 在植物休眠、萌芽、落叶等不同阶段,植物茎干含水量会呈现出不同变化规律。
茎干含水量与植物生理状态密切相关,但目前缺乏具体的评价方法。本文以欧美杨107杨(Populus×euramericancv.‘74/76’)为研究对象,利用活立木茎干含水量传感器实时获取植物茎干含水量参数,分析茎干含水量在不同季节的变化规律及与所在环境中各参数的变化关系,构建一种通过茎干含水量反映杨树生命状态的数学模型,以期为未来搭建树木生命状态评价体系提供模型基础和解决思路。
1 材料与方法
1.1 试验地点与材料
试验地点位于北京市共青林场(116°43′39.9792″E, 40°6′35.82″N),土壤类型为沙土,土壤pH在7~8之间。选取树势良好、茎干通直的欧美杨107杨作为试验样本,并采用相同的栽培管理方式,株行距约为4 m×3 m。 2014年林分平均胸径为(25.7±1.6)cm,平均树高为(17.5±1.6)m,平均叶面积指数为2.43±0.43。欧美杨107杨年内生长规律呈慢-快-慢的特点,其生长周期可划分为萌芽期(3—5月)、生长期(6—9月)、落叶期(10—11月)和休眠期(12—2月)。试验时间为2017年初—2018年初,试验地点年均气温12.5 ℃,年降水量592 mm,年晴天数131天。
1.2 参数测定
土壤温湿度会影响植物根系吸水速率,空气温湿度和饱和水汽压差会影响植物叶片蒸腾速率,光合有效辐射会影响植物叶片光合速率,这6个参数均与植物生理水分调节活动密切相关,将其统称为微环境参数集。试验中需直接测定的参数包括土壤温湿度、空气温湿度、光合有效辐射和茎干含水量,饱和水汽压差由空气温湿度计算得到。利用美国Decagon公司5TM型传感器(温度: -40~60 ℃,±1 ℃; 湿度: 0~100%,±2%)测量土壤温湿度,芬兰Vaisala公司HMP50-L6型传感器(温度: -10~50 ℃,±0.25 ℃; 湿度: 0~100%,±2%)测量空气温湿度,美国Li-Cor公司LI-190 R型传感器(0~10 000 μmol·m-2s-1,±5%)测量光合有效辐射,北京林业大学BD-IV型植物茎干含水量传感器(0~60%,±1%)测量茎干含水量。基于以上4种传感器搭建如图1所示的植物微环境参数与茎干含水量监测站,其中土壤温湿度传感器安装于地下50 cm处,空气温湿度和光合有效辐射传感器安装于距地面30 m高的监测塔上,茎干含水量传感器安装于距地面1.5 m高的杨树茎干上。

图1 植物微环境参数与茎干含水量监测站Fig.1 The monitoring station of plant micro-environment parameters and stem water content
1.3 茎干含水量传感器简介
基于驻波率原理(赵燕东等, 2002; 冯磊, 2005)的植物水分在线无损实时检测装置由100 MHz信号源、50 Ω同轴传输线、高频检波电路和平行不锈钢探测环组成(赵燕东等, 2011; 刘贺等, 2011)。茎干含水量传感器测量原理如图2所示。
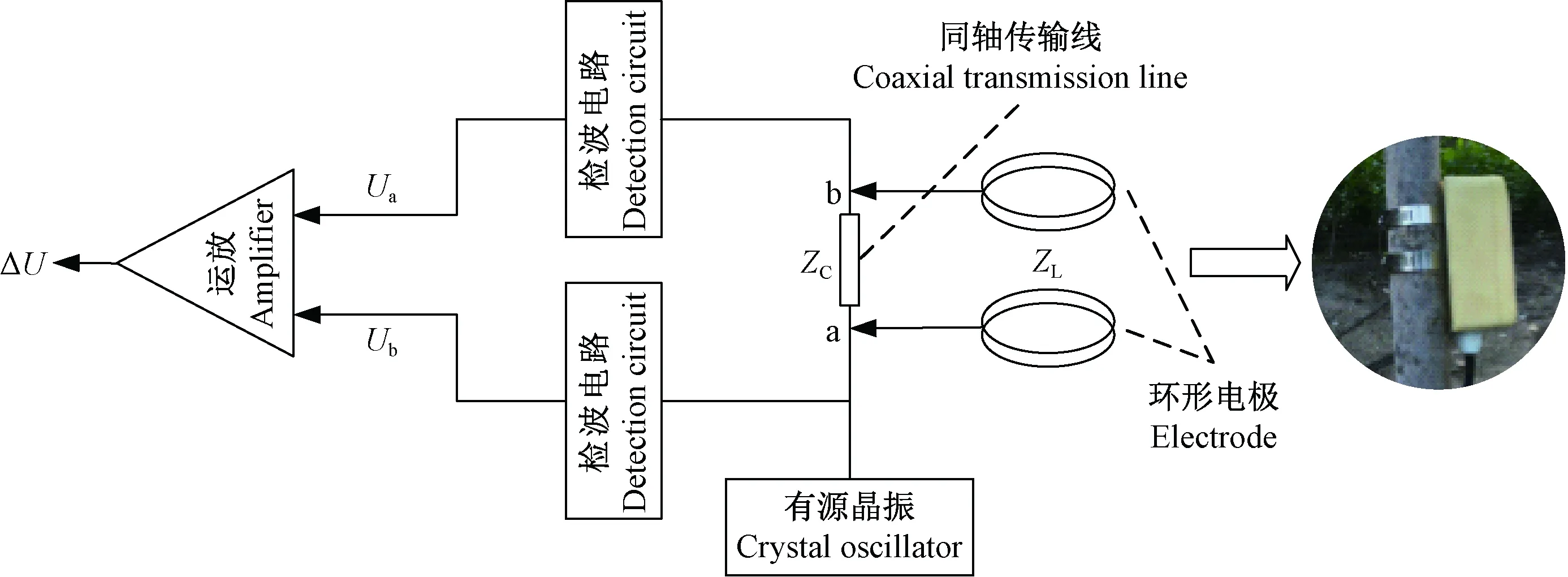
图2 传感器的测量原理Fig. 2 The measuring schematic diagram of the sensor
信号源产生的高频电磁波沿着传输线传送到探头(岳志勤等, 2014; 冯良平等, 2005),由于探头的阻抗与传输线的阻抗不匹配,一部分信号将反射回来。传输线上,高频入射波与反射波叠加形成驻波,在传输线上各点的电压幅值存在变化。根据驻波率测量原理,取图2中a、b两点的差动信号作为传感器变换电路的输出,即得到下式:
式中:A为信号的幅值;Zc为同轴传输线的特征阻抗(50 Ω);ZL为探测环阻抗。
为了进一步验证StWC传感器的测量性能,对标定后的StWC传感器、木材水分检测仪Testo606-2(德国Testo公司生产,测量范围0~60%,精度±1%)以及烘干法的测量结果进行对比,测量对象为紫薇(Lagerstroemiaindica)茎干,结果如图3所示,图中横轴为烘干法的测量结果,可将其视为StWC真值,纵轴为StWC传感器和Testo606-2测量结果,相比于Testo606-2,StWC传感器测量结果更接近于真值。当StWC真值大于2%时,Testo606-2的测量值显著小于真值,这是由于较短的探针(小于10 mm)限制了其有效测量深度,而在茎干烘干过程中,表层水分的蒸发速度大于深层。

图3 传感器与烘干法的测量结果Fig. 3 The measuring results by using sensors and oven-drying method
1.4 数据采集与处理
以上各种传感器均连接到自主设计的基于AVR128的多通道数字采集器上,每10 min采集1条数据,每日总计采集144条数据。剔除原始数据中因电压不稳或环境影响导致的奇异值,并使用插值方法填补缺失部分。采用6点均值滤波对数据进行平滑处理,用6个值的平均值作为1小时的数据,依次处理每日的144条数据得到以1小时为间隔的每日24条数据。监测6株树的茎干含水量,选取始终健康且数据最完整的1株树的数据进行试验。为了分析环境参数与茎干含水量的变化规律,减少环境参数之间的共线性问题,使用主成分分析法将环境参数数据转化为几种主成分,利用最大主成分中包含的信息与茎干含水量进行分析。
2 结果与分析
2.1 各生长周期茎干含水量的变化规律
2.1.1 萌芽期茎干含水量变化规律 萌芽期杨树茎干含水量的月变化规律见图4。从3月下旬开始,当旬均气温达到12 ℃以上时,杨树叶芽开始萌动,4月芽苞增大开放展叶,主茎和侧枝随着叶片展开开始生长,这一过程延续到5月上旬(宋玉光等, 2016)。萌芽阶段初期,杨树未长出叶片,光合作用和蒸腾作用强度低,植物体内需水量小。随着植物长出叶片,生命活动逐渐旺盛,需水量增大,茎干含水量逐渐上升至正常生长状态数值。此时叶片完全展开,杨树进入正常生长状态,茎干含水量停止上升保持稳定。4月初日均值20%,月末达到43%。
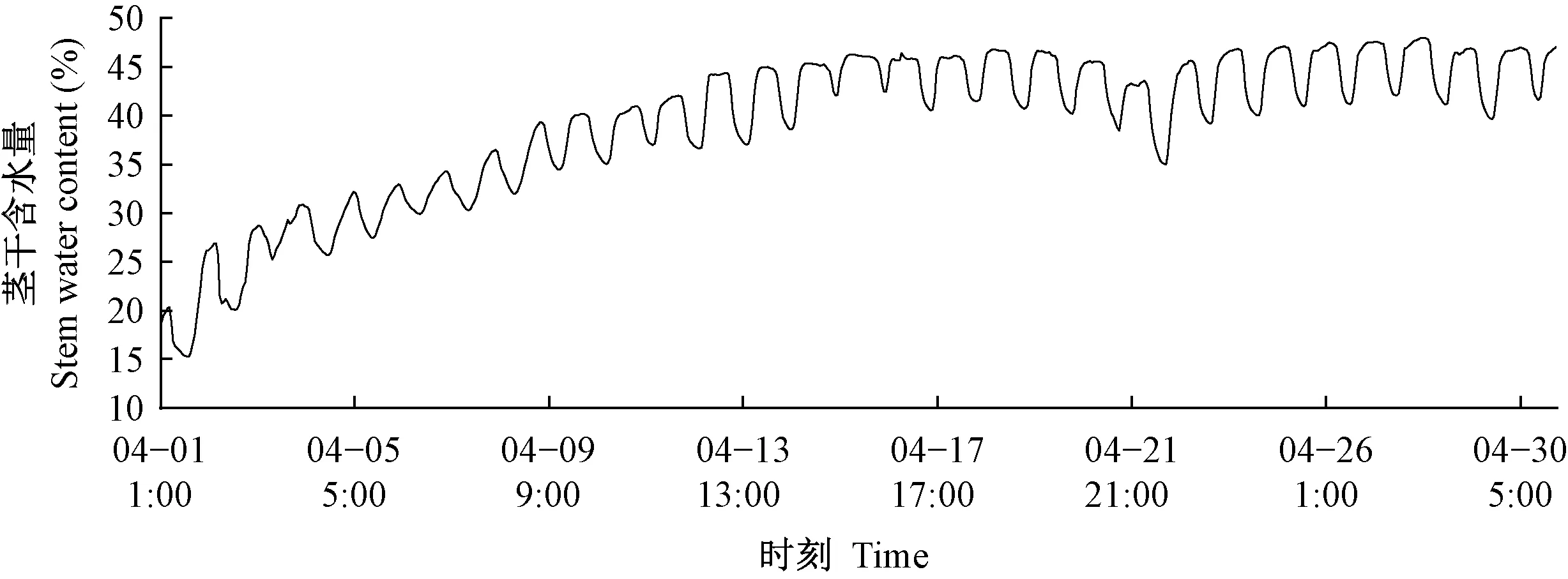
图4 2017年4月茎干含水量变化曲线Fig. 4 Stem moisture curve of April 2017
萌芽期杨树茎干含水量的日变化规律见图5。4月6日—8日茎干含水量变化与温度变化呈正相关。茎干含水量的日最小值30%出现在凌晨4点左右,日最大值39%出现在16:00左右,茎干含水量的日均值为33.8%,平均日波动幅值为8%。这3日处于茎干含水量上升时期,茎干含水量随空气温度保持增长趋势。这一阶段杨树处于生长期初期,该阶段空气温度较低,生命活动强度随太阳辐射强度增强而增强,同时蒸腾耗水量很低(贾天宇等, 2020),使得每日中茎干含水量与空气温度呈正相关。
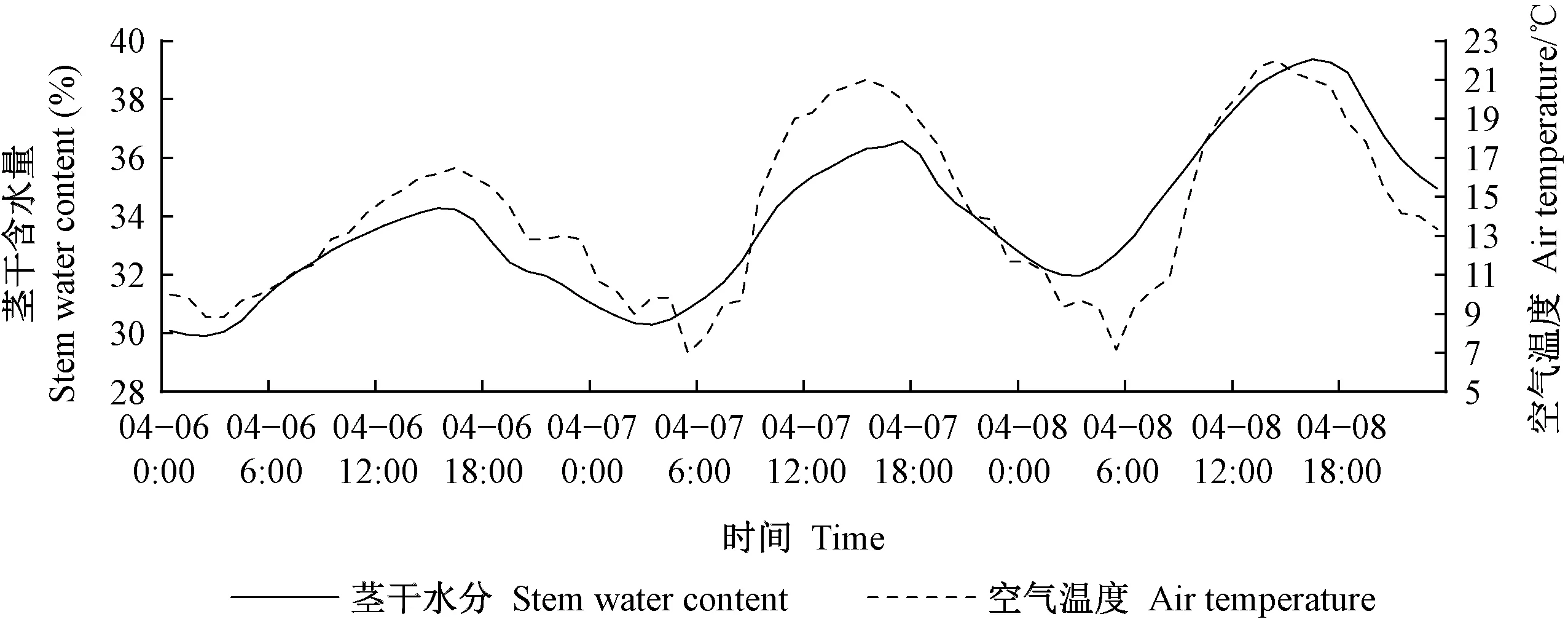
图5 2017年4月6—8日茎干含水量变化曲线Fig. 5 Stem moisture curve from April 6 to 8, 2017
2.1.2 生长期茎干含水量变化规律 随着气温升高和日照增强,杨树进入夏季生长期。杨树茎干含水量的月变化规律见图6。夏季炎热,白天植物蒸腾作用消耗大量水分导致含水量快速下降,夜晚进行水分补充使得含水率大幅提高(闫年等, 2011),因此夏季植物水分波动较春季更频繁和剧烈。夏季多雨,阴雨天气下杨树蒸腾作用大幅减弱,水分波动幅值较晴天差距较大。例如,7月9日前后为晴天,茎干含水量的日均值为47%,日波动幅度达到9%; 7月18日前后为阴雨天,日均值达到51.6%,日波动幅度仅为1%。茎干含水量的日变化规律与气象环境密切相关。

图6 2017年7月茎干含水量变化曲线Fig. 6 Stem moisture curve of July 2017
生长期杨树茎干含水量的日变化规律见图7。与春季相反,夏季茎干含水量与空气温度呈负相关。茎干含水量的日最小值39%出现在16:00左右,日最大值46%出现在16:00左右。茎干含水量的日均值为43.1%,平均日波动幅度为7%。夏季日出后光照强度增大,空气温度上升,此时是全年蒸腾量最大的阶段,蒸腾耗水大于根系吸水,导致茎干含水量下降。日落后光照强度减弱,空气温度降低,此时蒸腾耗水小于根系吸水,导致茎干含水量回升。由于夜间植物各项生理活动减弱,直到日出前茎干含水量会维持在一个相对稳定的水平。
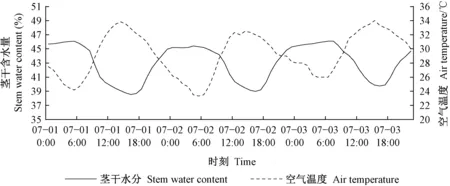
图7 2017年7月1—3日茎干含水量变化曲线Fig. 7 Stem moisture curve from July 1 to 3, 2017
2.1.3 落叶期茎干含水量变化规律 秋季杨树进入落叶期,并开始进行越冬准备。枝条生长停止,芽木质化加速,含水量逐渐降低。落叶期杨树茎干含水量的月变化规律见图8。茎干含水量日最大值从月初的47.1%降低至月末的39.3%。秋季空气温度下降,杨树叶片脱落,光合作用、蒸腾作用等生命活动强度减弱,杨树需水量下降,茎干含水量逐渐降低。这一降低趋势会延续到叶片完全脱落,直至杨树进入休眠期,自由水含量降到最低值,茎干含水量变化才稳定。
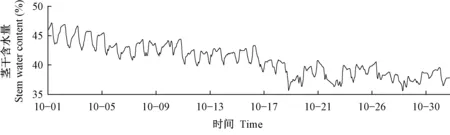
图8 2017年10月茎干含水量变化曲线Fig. 8 Stem moisture curve of October 2017
落叶期杨树茎干含水量的日变化规律见图9。秋季的环境日变化情况与夏季类似,因此茎干含水量的日变化规律与夏季类似。

图9 2017年10月2—4日茎干含水量变化曲线Fig. 9 Stem moisture curve from October 2 to October 4, 2017
2.1.4 休眠期茎干含水量变化规律 休眠期杨树茎干含水量的月变化规律见图10。北京地区冬季最低气温可降至零下15 ℃,在冬季日均气温长时间低于0 ℃,杨树体内自由水分会产生冻结。茎干含水量日最小值从12月初的40%下降至日最小值12%。同时由于茎干含水量变化受冻融影响程度超出植物自身调节能力,冬季茎干含水量的日波动幅度也最剧烈。12月25日茎干含水量的日最小值为15%,最大值为35%,波动幅度为20%,这已远远超过了植物正常进行生命活动时茎干含水量的日波动幅度。水分冻结期间茎干含水量大幅度降低,茎干含水量日均值远低于植物其他时期进行正常生命活动时的日均值。
12月2—4日茎干含水量的日变化曲线见图11a。这一阶段茎干含水量未冻结,日均值为43.6%,日变化幅值小,保持在2%以内,波动趋势与温度变化趋势相同。当温度降低至0 ℃以下时,茎干含水量的日最小值降低至40%,随着温度上升又恢复至43.6%,之后持续降温使茎干含水量产生冻结。

图11 不同状态下茎干含水量日变化曲线Fig. 11 Diurnal variation curve of stem moisture under different conditions
12月12—14日茎干含水量的日变化曲线见图11b。这一阶段茎干含水量已产生冻结,基于仪器读数的日均值为20.2%,远低于其他时期。茎干含水量的日最大值29.3%出现在18:00左右,比空气温度的日最大值时刻滞后1 h,日最小值12.8%出现在上午10:00左右,与空气温度日最小值时刻相同。茎干含水量的日波动幅度为16.5%,是全年其他时期日波动幅值的2倍以上。这一阶段茎干含水量变化主要受温度变化产生的水分冻融影响,茎干含水量与空气温度相关性很强,日变化幅值为全年最大。
2.2 各生长周期茎干含水量随微环境参数变化的规律
2.2.1 模型构建 经过4个生长周期的茎干含水量变化分析,可以看出水分随时间推移呈现周期性变化,且不同程度地受环境因素影响。本研究拟建立一种模型,客观表征茎干含水量与环境因素的关系。
微环境参数之间的有较强的相关性,引发严重共性问题,且部分参数(如空气温度与空气湿度)对植物生理状态产生相反的影响(高超等, 2017)。为了解决这一问题,本文采用主成分分析法将环境参数变换为一系列主成分。为了保证不同阶段主成分的一致性,将4个阶段的环境参数统一进行主成分分析。变换为主成分后,环境参数特征得到保留,几种主成分之间相互独立。变换后主成分特征值及贡献率如表1所示,其中最大主成分PC1贡献率达到了81%以上,可以保证数据信息的完整性。表2为最大主成分PC1中4种环境参数系数值,可以看到空气湿度系数值最大且为正值,其他3个系数值比空气湿度系数值小很多且为负值,即PC1与空气湿度呈正相关且相关性较强,与空气温度、光合有效辐射、饱和水气压差呈负相关。
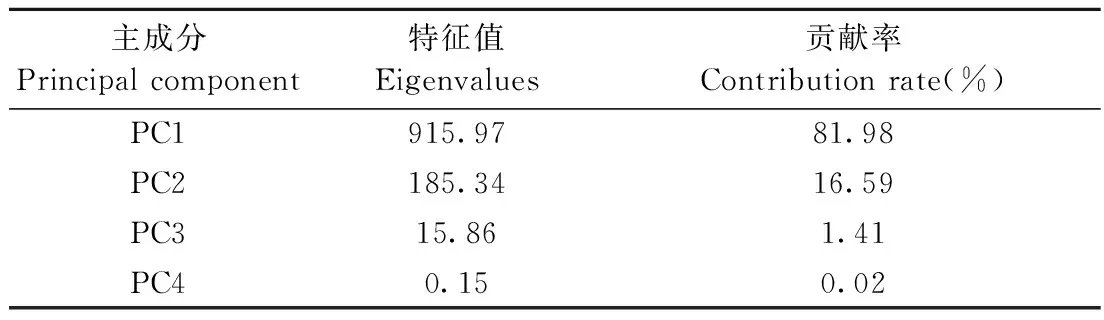
表1 主成分特征值及贡献率Tab.1 Principal component eigenvalue and contribution rate

表2 PC1中4种环境参数特征系数Tab.2 Four environmental parameter characteristic coefficients in PC1
选择最大主成分PC1为横轴变量,与茎干含水量绘制日变化曲线(图12)。茎干含水量与PC1呈现斜椭圆的变化规律,箭头方向为下一时刻的观测值。
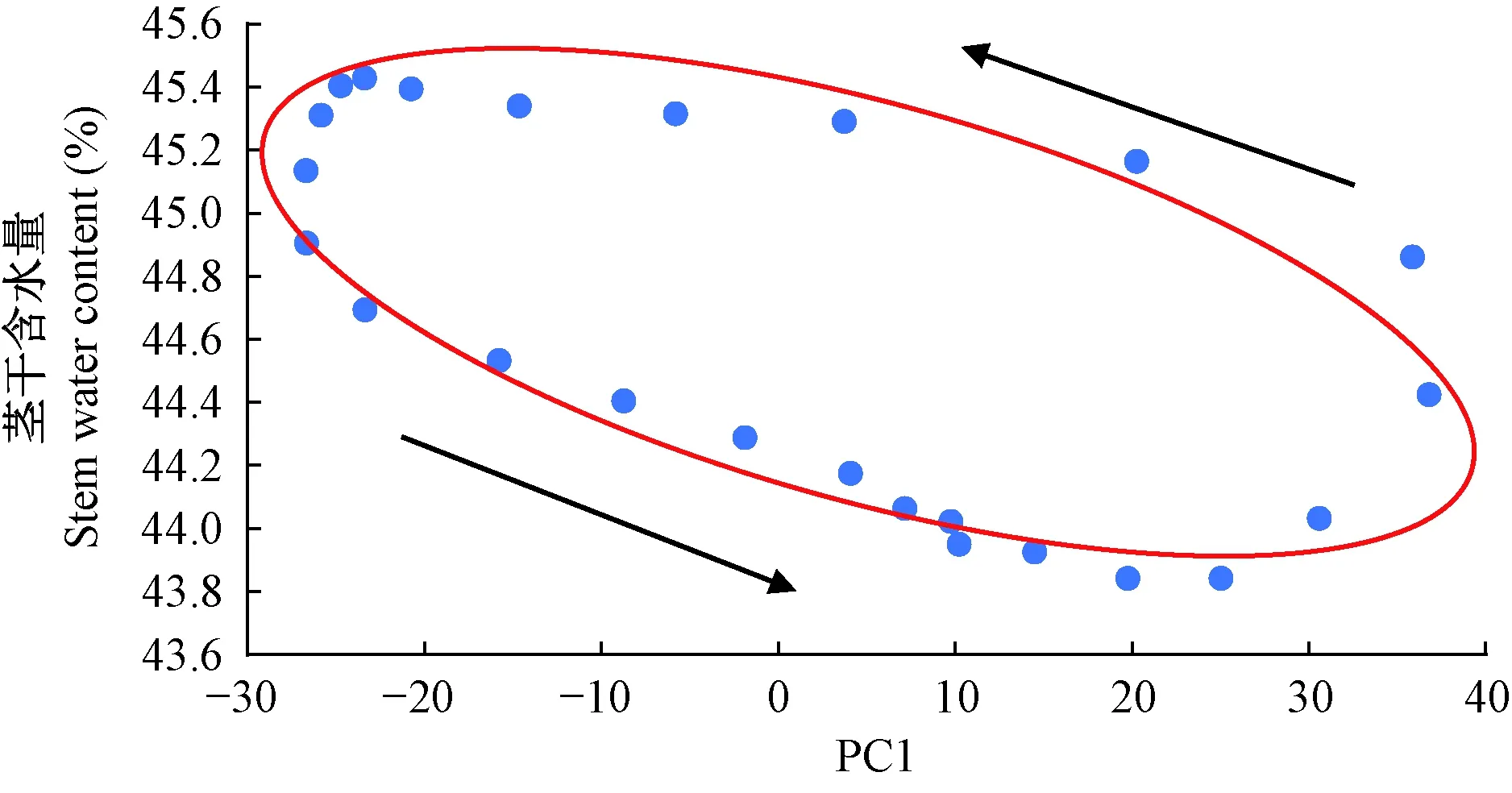
图12 茎干含水量与PC1日变化关系Fig. 12 Relationship between stem moisture and PC1 diurnal variation
式中:A为斜椭圆的长半轴;B为斜椭圆的短半轴;S为斜椭圆中心点的横坐标;T为斜椭圆中心点的纵坐标;θ为斜椭圆长轴按顺时针旋转与X轴正方向的夹角(0°<θ<90°);X为微环境参数集的最大主成分PC1;Y为茎干含水量(0~100%)。
该模型中A、B、S、T、θ均为预估参数,可由数值稳定的直接最小二乘法估算得出(Mulletietal., 2015; Panagiotakisetal., 2016; Maini, 2006),其中S表示日平均最大主成分,T表示日平均茎干含水量。tanθ表示斜椭圆长轴的斜率,可定义tanθ为茎干含水量相对于微环境参数集最大主成分的灵敏度,扁率e=(A-B)/A表示斜椭圆的扁平程度,可定义e为茎干含水量相对于微环境参数集最大主成分的相关度。X为唯一输入变量,Y为输出变量。
对4个生长阶段中单日的数据进行曲线拟合,结果如图13所示。
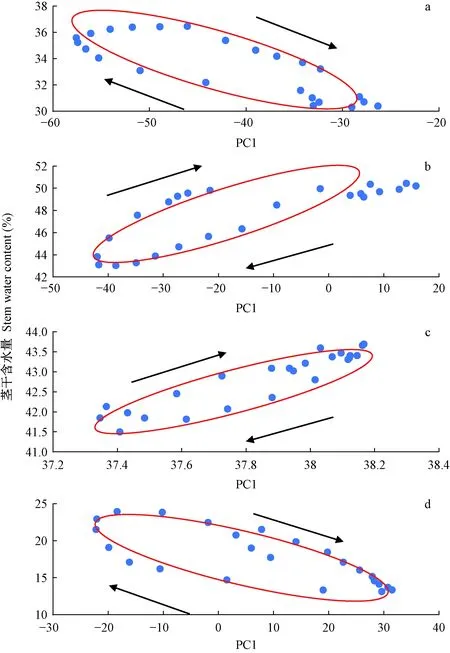
图13 4个阶段的茎干含水量拟合椭圆曲线Fig. 13 Four stages of stem moisture fitting elliptic curve
椭圆的标准方程经过平移和旋转变换后,可得斜椭圆的标准方程,其数学解析式为:
2.2.2 模型分析 将图13中4个阶段的椭圆方程参数值提取整理后得到表3,展示拟合椭圆的5种参数,表征椭圆的具体形状及位置:A与B的长度与θ联立得到在坐标轴的投影为环境参数与茎干含水量每日的变化幅度。横纵坐标的均值即为椭圆圆心坐标值S与T,S为多种环境参数综合得到的指标,S的大小为抽象表现不同阶段的环境的区别,无实际意义; T为茎干含水量日均值。

表3 椭圆拟合曲线参数Tab.3 Ellipse fitting curve parameter
θ的区别:θ为椭圆长轴与X轴夹角,设定夹角θ的范围为-90°~90°。萌芽期和休眠期的θ为负值,生长期和落叶期的θ为正值。θ>0°表明茎干含水量与PC1为正相关,θ<0°则为负相关。因为PC1由空气湿度为主导,空气湿度与空气温度变化趋势相反,所以茎干含水量与PC1的变化规律与茎干含水量与空气温度变化规律相反。其中生长期θ绝对值比其他阶段小,表明该阶段茎干含水量受环境影响最小。
超前与滞后: 根据椭圆旋转方向与倾角,可以推断出茎干含水量与环境参数的超前与滞后关系。在图13a和13d中,PC1在茎干含水量达到最大值前到达最小值,PC1在茎干含水量到达最小值前到达最大值,即茎干含水量负相关滞后于PC1。在图13b和13c中,茎干含水量在PC1到达最大值前到达最大值,茎干含水量在PC1到达最小值前到达最小值,即茎干含水量正相关超前于PC1。与图5、7、9、11对照,符合时序状态下茎干含水量与空气温度超前、滞后关系。即茎干含水量在萌芽期与休眠期滞后于环境参数,在生长期与落叶期超前于环境参数。
3 结论
本文针对杨树茎干含水量在不同生长期间内的每日规律性变化,利用椭圆曲线拟合植物茎干含水量与环境参数的关系。通过分析拟合椭圆方程的参数发现,杨树茎干含水量在不同生长阶段有不同的变化规律,对外界环境变化响应也不同。萌芽期茎干含水量不断增大,与空气温度变化呈正相关; 生长期茎干含水量稳定波动,与空气温度变化呈负相关; 落叶期茎干含水量逐渐降低,与空气温度变化呈负相关; 休眠期茎干含水量受冻融影响波动剧烈,与空气温度变化呈正相关。茎干含水量椭圆模型的各参数[倾斜度(θ)、椭圆圆心的纵轴坐标(茎干含水量日均值)、椭圆的横轴坐标(PC1日均值)]在不同生长状态下有很强的区分度,可利用这些参数对植物生长状态进行判断。
