中国典型生态脆弱区生态化学计量学研究进展
2021-07-12李玉强王旭洋牛亚毅
陈 云,李玉强,3,*,王旭洋,牛亚毅
1 中国科学院西北生态环境资源研究院, 兰州 730000
2 中国科学院大学, 北京 100049
3 中国科学院西北生态环境资源研究院奈曼沙漠化研究站, 通辽 028300
生态脆弱区又称生态交错区(Ecotone),主要位于农牧、林草、林牧等不同生态系统交错带[1-3]。我国生态脆弱区分布广泛、类型多样、表现突出[4],中度以上生态脆弱区占我国陆地国土面积的55%[5]。尽管针对生态脆弱区生态保护与恢复的研究成果较为丰富,但是不同研究生态脆弱区范围划分结果不尽一致[4,6-7],且生态脆弱区生态系统稳定性差、抗干扰能力弱、易退化。在极端气候事件频发、氮(N)沉降、CO2浓度上升、土地利用变化等全球环境变化和人类活动的影响下,生态脆弱区生态系统日益敏感、脆弱。然而,退化生态系统恢复与控制对应对气候危机、粮食安全、供水以及生物多样性等至关重要[4],生态脆弱区生态系统恢复是实现我国社会经济可持续发展的必由之路[2],《中华人民共和国环境保护法》也明确规定“国家在重点生态功能区、生态环境敏感区和脆弱区等区域划定生态保护红线,实行严格保护”。本文依据“全球变化及应对”重点专项项目“全球变化对生态脆弱区资源环境承载力的影响研究”[8]前期研究成果,将我国典型生态脆弱区划分为六类:农牧交错带(包括北方农牧交错带和西南山地农牧交错带)、林草交错带、荒漠-绿洲区、黄土高原脆弱区、青藏高原脆弱区和西南岩溶石漠化区(图1)[1]。

图1 我国典型生态脆弱区分布范围
生态化学计量学(Ecological stoichiometry)连接了生物系统内从基因分子结构到生态系统的各个层次[9-11],是一门综合生态学、生物学、化学等多学科基本原理,研究生态系统能量、元素平衡(主要为碳(C)、N、磷(P)元素)的学科[12-13]。开展生态系统C、N、P元素生态化学计量学研究具有重要的生态意义:(1)通过N∶P比值关系来判断植物受两大生命元素N、P的限制关系[14],进而促进对陆地生态系统养分调控和元素生物地球化学循环机制的深入理解;(2)通过生态化学计量学研究生物养分利用效率、策略和生物内稳性机制,促进生物对环境适应性的深入理解[15-17];(3)C、N、P元素关系对生态系统C循环和生态系统对环境变化的响应具有重要指示作用[18-19]。为了深入理解我国生态脆弱区对全球环境变化的响应和生物地球化学循环机制,重点针对这六类典型生态脆弱区的植物、凋落物、土壤和土壤微生物C、N、P生态化学计量特征及其对环境变化响应的研究进展进行总结,以促进生态脆弱区生态系统保护与恢复和生态化学计量学研究的发展。
1 生态脆弱区C、N、P生态化学计量特征
1.1 植物叶片生态化学计量研究
N、P元素是构成蛋白质、磷酸等的基本元素,对调节植物生长和代谢具有重要作用[9],在CO2浓度升高、N沉降等全球环境变化背景下,植物叶片C、N、P元素及其相互关系成为生态化学计量学关注的热点之一。由于区域空间异质性,我国典型生态脆弱区C、N、P生态化学计量存在较大差异(表1)。有研究指出,地理变异和种间变异是造成植物叶片化学计量差异的主要原因[25];此外,植物功能型亦能较好的解释叶片中元素含量的差异[56]。与禾本科植物相比,豆科植物N含量及N∶P比值更高,并以P限制为主[57-59]。在荒漠化地区和青藏高原,灌木和草本植物C储存能力较低,灌木N、P含量和N∶P比值低于草本和乔木[27-28,32],而喀斯特地区灌木叶片C含量最高,草本植物叶片N、P含量最高,各元素化学计量比值表现为乔木>灌木>草本[33]。在植物发育上,随植物年龄增加,植物叶片C、N、P含量增加[60];成熟叶片N、P含量低于幼叶,而N∶P比值高于幼叶[33],主要是因为幼叶生长发育对N、P元素需求大,特别是P元素[61]。
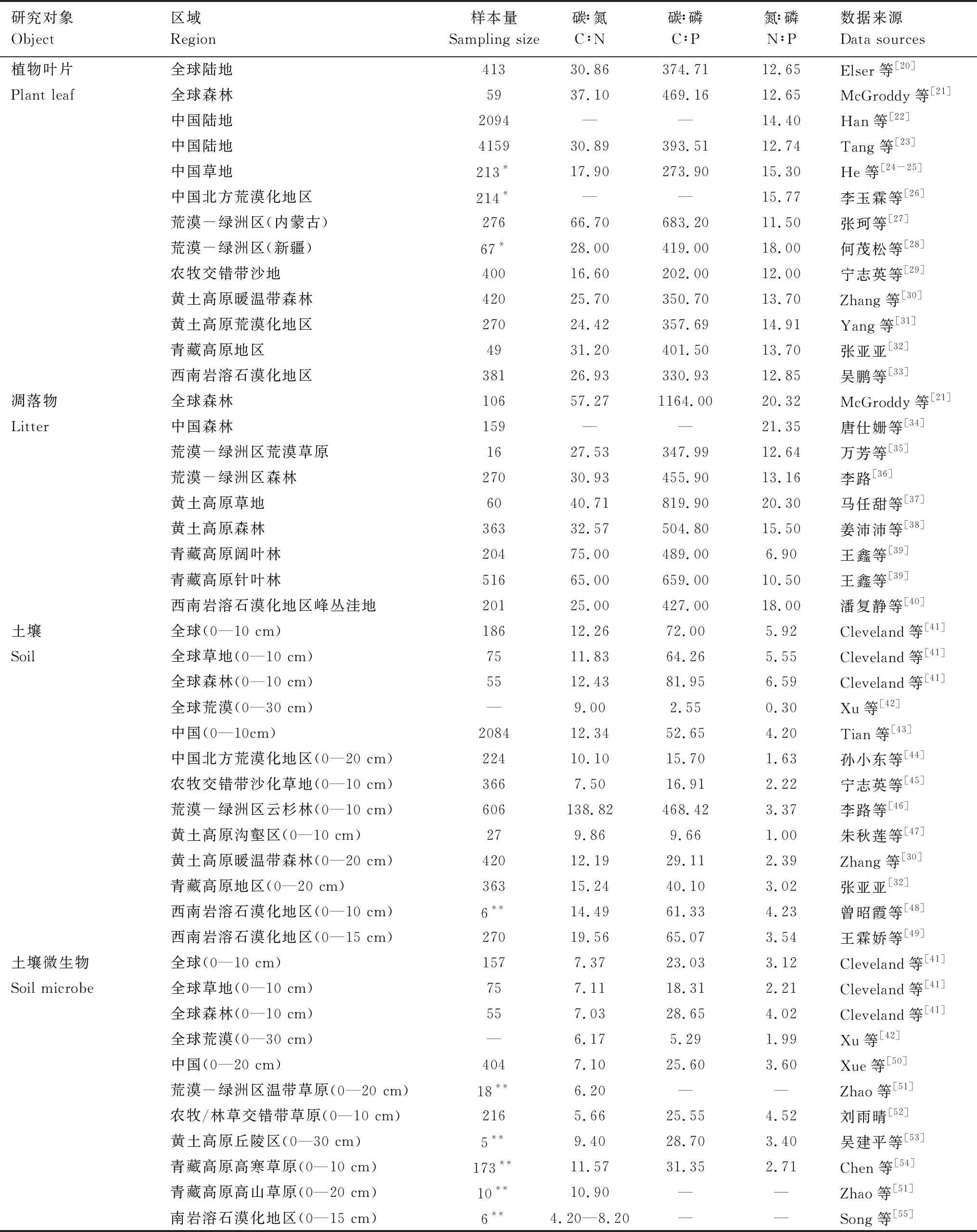
表1 不同区域C、N、P生态化学计量比值(质量比)特征
生长速率假说认为高生长速率对应高N、P含量和低C∶N、C∶P与N∶P比值,N∶P比值高低又被广泛用于营养限制的判断依据。基于N∶P比值可以反映群落水平营养限制状况的假设,Koerselman和Meuleman对欧洲淡水湿地生态系统施肥实验的回顾研究表明,当N∶P<14,在群落尺度上受N限制;当N∶P>16,受P限制;当N∶P比值介于14—16时,可能同时受或不受N、P元素限制[14]。但受区域环境、物种组成、植物年龄、组织器官等多因素的共同作用,对于不同的研究对象,营养限制判断阈值存在差异[62]。Güsewell对温带和寒带植物群落的研究提出N∶P<10和N∶P>20通常是判断植物群落N、P限制的临界值,但由于施肥实验的方法差异、时间依赖等诸多原因,营养限制并非总是明确的,施肥的长期效应或对个别物种N、P关系的影响具有差异[62]。再者,多因素的综合作用导致难以就某一地区植物营养限制状况做出准确判断,已有研究表明N∶P比值关系不能有效预测荒漠及荒漠化地区植物养分限制状况[26,29,63]。事实上,养分限制状况是复杂的,在群落内,不同物种具有不同的养分利用策略,以占据不同生态位而达到一定的平衡[15]。总体而言,在我国北方荒漠及荒漠化地区植物叶片似乎具有更高的N∶P比值而更容易受P元素限制[26,28,31],而在青藏高原、喀斯特地区和黄土高原等生态脆弱区则更容易受N元素限制[30,32-33]。
此外,内稳性是生态化学计量学的重要内容之一,但不同物种化学计量内稳性特征存在较大差异,如Yu等[64]研究表明内蒙古草原维管植物内稳性指数变化范围为1.9—14.5。在物种水平上,化学计量内稳性与优势度和稳定性呈正相关;在群落水平上,内稳性多与生态系统功能和稳定性呈正相关[65]。这在宁志英等[17]的研究中得到验证,即在N相对贫瘠的沙化草地,拥有更高的N、N∶P内稳性特征的植物在多变环境中表现出更强的适应性和竞争力。因此,在全球环境变化背景下,植物化学计量所表现出的内稳性特征有助于我们深入了解物种稳定性,以及物种和生态系统对全球环境变化的适应能力[66-68]。
1.2 凋落物生态化学计量研究
凋落物养分积累与分解是陆地生态系统C和养分循环的重要过程,反映了生态系统动态过程与能量平衡过程,联系了植被与土壤[30,69]。同时,凋落物所含养分也是土壤养分的重要补给来源,并与土壤养分呈显著正相关[40]。凋落物养分积累与释放过程主要受其初始化学元素控制[69],其中,有效性N、P养分及其比值是影响凋落物分解的重要决定因素[70]。对内蒙古草地生态系统的研究表明,N有效性增加,将促进凋落物分解[71-72],但有研究认为当凋落物具有较高N含量和较低P含量(即高N∶P比值),尤其是当N∶P大于25,且P含量小于0.22 mg/g时,会限制凋落物分解速率[73]。总之,凋落物C、N、P含量高低及其比值大小显著影响土壤养分积累的数量和速率,进而影响植被生产力[74-75]。
潘复静等[40]对喀斯特地区不同植被演替序列的研究表明,凋落物C、N、P含量呈现常绿落叶阔叶林>灌丛>草丛的趋势,N∶P比值随正向演替(演替序列依次为草丛群落、灌丛群落、常绿落叶阔叶林群落)而升高。对于青藏高原不同林型,针叶林凋落物C、N含量及C∶P、N∶P高于阔叶林,P含量低于阔叶林,N、N∶P比值与纬度呈显著负相关[39];但姜沛沛等[38]认为黄土高原森林凋落物N∶P比值与经纬度、海拔的相关性不显著。然而,凋落物化学计量的全球格局研究表明,在低纬度地区N∶P最高,中纬度地区N∶P最低,向高纬度地区N∶P又逐渐增加,因为低纬地区P含量相对较低,而高纬地区虽然P矿化速率较低但再吸收率较高[76]。此外,不同尺度研究结果表明,凋落物N、P、N∶C和P∶C低于植物叶片,说明养分再吸收是植物活有机体保持养分、提高生产力的重要机制[35,38],养分再吸收率高低反映了植物在贫瘠、脆弱生态系统中的适应能力[35]。
1.3 土壤生态化学计量研究
土壤是陆地生态系统重要的C库和营养源,土壤C库储量是大气C库的3.3倍,是生物C库的4.5倍[77],土壤养分是植物生长的物质基础,土壤C、N、P含量及其化学计量比是衡量土壤质量的重要指标,但受成土因素和人类活动的影响而存在较大差异[18]。在生态脆弱区,土地荒漠化造成土地质量严重退化,土壤养分大量流失,改变了土壤C、N、P化学计量特征与平衡,限制了植被生产力[45,78]。有研究表明,在荒漠-绿洲区、黄土高原、农牧交错带等生态脆弱区表层土壤C、N、P元素平均含量低于我国表层土壤平均水平[43,45-47];同时,生态脆弱区具有相对较低的土壤C∶N∶P化学计量比值,这或许可以在一定程度上解释生态脆弱区植被生产力相对较低的原因。
土壤C∶N∶P化学计量存在明显的空间依赖。研究表明,我国草地生态系统表土C∶N、N∶P比值在青藏高原沿东南向西北、在内蒙古高原沿东北向西南递减,且内蒙古高原温带草原土壤N∶P比值明显高于青藏高原高寒草原,而新疆北部草原土壤C、N、P化学计量空间变异性较小[79]。在黄土高原,土壤C、N、P含量及其比值呈现显著的纬度变化趋势,C、N和C∶N∶P随纬度升高呈指数下降,P则先增后减[80]。喀斯特地区土壤N、P含量较低[81],随纬度增加,土壤C、N、P含量下降[82]。此外,土壤养分具有“表层聚集”效应,即随土壤深度的增加,C、N、P含量呈减少趋势[31,83-85],但在喀斯特地区,也有研究认为表层土壤N供应能力较强,而深层土壤P供应能力更强,因为降水导致P元素向下迁移[81]。在时间演变上,近期一项在内蒙古高原和青藏高原的研究表明,从20世纪80年代起,N∶P比值呈增加趋势,主要是因为N、P元素非耦合变化,即P在强风化土壤中易淋溶损失,而N通过生物固N和大气N沉降保持相对稳定,从而可能导致P限制[79]。
凋落物质量、植被类型及覆盖度与土壤养分也呈现较强的相关性[74,82,85-86]。凋落物养分化学计量影响微生物的分解速率,进而影响养分向土壤归还与释放过程[31,74]。植被类型对生态脆弱区土壤养分的影响主要表现在乔木、灌木、草本等不同功能型植被下土壤养分的差异。刘秉儒[87]在荒漠-绿洲区山地的研究认为,土壤C、N含量表现为草地>森林>荒漠;朱秋莲等[47]认为黄土高原丘陵沟壑区土壤养分含量表现为森林>草原>森林草原区;而喀斯特地区呈现林地>灌丛>草地的趋势[85]。
1.4 土壤微生物生态化学计量研究
土壤微生物是陆地生态系统植物生产力与多样性的重要调节因子和驱动力[88],在生态脆弱区,土壤微生物量C、N、P和土壤养分关系密切,两者化学计量具有相对一致性[50,87]。一方面,虽然凋落物质量影响土壤微生物的分解速率,但土壤微生物也可通过调整自身养分利用效率,如提高养分利用效率以适应低养分含量(高C∶养分比值)下凋落物分解,进而促进凋落物养分释放[69];另一方面,土壤因子(C、N、P含量及其比值,土壤温度等)是引起微生物群落结构空间变异的主要驱动力,土壤微生物量化学计量主要受土壤养分控制[89-92]。同时,土壤微生物群落结构和微生物量C、N、P化学计量也表现出显著的随土壤深度变化的规律,即随土壤深度增加,土壤微生物量C、N、P呈降低趋势[53,93]。土壤深度引起土壤微生物群落组成变化主要表现在表层土壤微生物以好氧菌为主,深层以厌氧菌为主[93]。除了土壤因子,气候因子和生物因子(地上生物量、植物丰富度等)也是影响土壤微生物量C、N、P的重要因子,如气候因子、土壤因子和植物群落共同解释了内蒙古草原土壤微生物变异[94],但对高寒草地土壤微生物量变异的解释程度低于土壤因子[89]。Zhao等[51]则认为气候因子对内蒙古至西藏温带草原和高寒草地土壤微生物量C、N的影响大于土壤因子,解释了26%的变异。刘秉儒[87]认为气候因子、土壤因子是影响贺兰山土壤微生物量C、N沿垂直梯度变化(微生物量C、N随海拔增加而增加)的关键因子。
土壤微生物量的全球格局表明,随着纬度增加,微生物量C、N、P含量增加,N∶P比值降低[42,95],在内蒙古高原和青藏高原草地样带上也发现了相同的N∶P变化规律[52,54]。在内蒙古草原,土壤微生物量N∶P比值表现为荒漠草原>典型草原>草甸草原,C∶N、C∶P无显著差异[52];在荒漠-绿洲区山地,土壤微生物量C、N表现为高山草甸>林地>灌丛>荒漠化草原,而C∶N无显著差异[87];在青藏高原高寒草原和高寒草甸,C∶N、C∶P和N∶P比值表现为高寒草原>高寒草甸,C∶N∶P比值显著高于全球平均水平,土壤微生物量C∶N∶P化学计量的变异主要由微生物群落结构变化引起[54];在喀斯特地区,土壤微生物量N∶P无显著变化,而C∶N、C∶P草地最高[55]。总体而言,土壤微生物量C、N、P化学计量关系受植被影响显著,与荒漠、灌丛和森林相比,草地土壤微生物可能具有更高的养分利用效率[55,87]。
有研究指出,土壤微生物量C、N存在明显的季节变化,微生物量C、N春季最高,而秋季最低,因为春季气温上升,土壤微生物活性提高,促进养分吸收利用,返青期根系分泌也为微生物提供能源,随着植物进入生长旺盛期,养分利用竞争加剧,限制土壤微生物活动,而秋季之后,凋落物增加和根系死亡促进了微生物活动;同时,C∶N比值也存在季节动态,即夏季低,冬季高[93]。此外,土壤微生物熵也被用来指示生态系统的动态变化,它指土壤微生物量C、N、P含量分别与土壤有机C、总N、总P含量的比值,即单位资源所支持的微生物量,常被用于生态系统养分积累与恢复研究[96],表征土壤有机质输入、养分转化效率、养分流失等状况[97],是快速识别环境变化的稳定性指标[98]。
2 生态化学计量对环境变化的响应
2.1 水热条件
区域水热条件是生态系统重要的环境因子。尽管C、N、P化学计量在大尺度上存在沿纬度(温度)变化的规律,但对于不同生态脆弱区而言,这种变化在植物、凋落物、土壤及微生物中具有差异[38-39,52,80,82,89,99]。在高寒地区,温度是限制植物生产力的主要因子之一。在青藏高原高寒草地,植物N含量和N∶P比值较高,P含量较低,以P限制和N、P共同限制为主,这可能是导致高寒地区植物生长速率较低的原因[100-101]。模拟增温实验表明,增温提高了高寒草甸植物光合作用速率和生长速率,促进植物地上部分生长,但降低了植物对N的吸收,提高了C∶N比值和N素利用效率[102]。但有研究指出,在群落水平上,植物叶片N、P含量随海拔升高而增加,植物通过增加N、P含量补偿低温下光合速率的降低,即温度-植物生理假说[99]。同时,气候变暖将加速土壤微生物呼吸作用和代谢活动,促进土壤微生物的分解活动,加速土壤C流失[103-104]。
在我国北方干旱半干旱地区,植物生长更容易受N限制,但植物对N添加的响应需要较长时间,而水添加可显著提高植物地上净初级生产量[105],说明水分是北方干旱半干旱地区生态系统初级生产力的重要限制因子[106-107]。植物叶片和土壤C、N、P含量与降水量呈正相关[54,80,82,99],一是因为降水增加,叶片代谢活动增强;二是降水促进土壤矿化作用,使土壤N、P含量增加[99]。研究表明,随年均降水量减少,植物叶片N∶P比值增加[52,108],随干旱程度增加,植物生长受P限制作用加强,这种响应取决于功能型[109]。增雨可增加表土C储量,但增温、增雨改变C、N含量平衡,可能导致土壤固C过程的N限制[103]。在荒漠地区的雨量控制实验表明,干旱处理显著提高土壤微生物量C、N[93]。然而,在喀斯特地区,水土流失严重,气温、降水、植被覆盖率和岩石出露率是土壤养分积累和循环的主要驱动因子,高强度降水与岩石作用,加速岩石风化和N、P养分释放,使岩石周围斑块N、P含量增加;同时,高降水与高气温的相互作用,将增强土壤微生物活性和加强有机质分解,加速土壤C流失[82]。
2.2 CO2浓度升高
人类活动导致大气CO2浓度持续上升,CO2排放增加是全球气候变暖的主要原因之一[110]。CO2作为光合作用的底物,其浓度增加将影响植物光合速率和养分利用。对荒漠植物的模拟实验表明,CO2浓度增加使植物光合速率先增后降,即短期CO2浓度增加,植物固C速率提高,增加植物生物量,但在长期高CO2浓度下植物光合速率将下调,即“光合适应”;同时,气孔导度和蒸腾速率随CO2浓度增加而下降[111]。由于植物对N吸收的主要方式是质流,气孔导度和蒸腾速率下降将导致植物N吸收和同化速率下降[112]。Shi等对内蒙古干旱半干旱草地禾本科植物的研究则发现, CO2浓度升高导致N含量相对下降是“同化抑制”和“稀释效应”共同作用的结果[113]。这种稀释效应在内蒙古荒漠和西南农牧交错带也有发现,对于荒漠灌木中间锦鸡儿(Caraganaintermedia),CO2浓度升高导致N素分配至叶片的比例下降[114];而对于川西亚高山红桦(Betulaalbosinensis),尽管CO2浓度升高使N、P含量升高,但P含量增加的幅度更大,且N分配至叶片的比例也是下降的[115]。相反,Xiao等人在黄土高原的研究认为CO2浓度升高增加了植物N、P和N∶P,促进了N向地上部分分配[116]。说明植物对CO2浓度变化的响应因物种和功能型而异。许多研究都报道了CO2浓度升高促进了地下部分发育和养分增加[114-116],原因可能是CO2浓度升高促进了土壤N矿化和增加了土壤有效P含量,有利于根系吸收[116-118]。总体而言,CO2浓度变化对生态脆弱区生态系统C、N、P化学计量的影响研究十分匮乏,影响机制也尚不明确,有待深入研究。
2.3 N沉降
在植被破坏、生物量燃烧、工业扩张等人类活动影响下,我国大气N沉降发生了深刻变化,作为生物地球化学循环的重要部分,大气N循环和沉降速率变化引起了对人类健康和生态系统可能造成不利影响的担忧[119]。模拟大气N沉降的N添加试验,不仅可以探索生态系统对N沉降变化的响应,也可以揭示生态系统养分限制情况。在植被水平,N添加提高地上C库和生物量N浓度,养分利用效率也随N添加水平增加而增加;而在物种水平,豆科植物N和N∶P基本保持稳定,非豆科植物则显著增加[120]。在受N限制的北方半干旱草地,施N提高群落N、P含量和N∶P比值,N添加促进了N、P元素耦合和植物对P的吸收[121]。在内蒙古温带草原的实验表明,P对植物叶片养分和化学计量的影响依赖于N供应,在环境N含量下,P添加对叶片N、P含量和N∶P比值没有显著影响,结合N添加会显著增加植物叶片N、P含量,同时改善土壤性质,但随N含量增加,P限制会增强[122]。植物对N添加的响应并不总是一致,在P含量相对较低的荒漠草地、高寒草地,N添加抑制植物P含量[123-124],因为P元素依赖于岩石风化[25],N添加不能有效增加土壤有效P含量,限制了植物对P元素的吸收,植物通过提高对老叶P再吸收率来达到新的养分平衡[124]。而Zhang等[125]认为N添加对植物P含量和N∶P无显著影响,P添加对植物N无显著影响,但显著降低了N∶P比值。总之,物种、叶龄、土壤肥力、N和P添加水平以及水分、养分利用特征不同是导致植物群落对环境变化差异性响应的重要原因[124-126]。
N添加处理增加半干旱草地凋落物N含量和N∶P比值,降低C∶N比值,对P含量和C∶P的影响不显著[126-127]。在N添加下,凋落物分解的第一年以P固定为主,随后为净释放,随分解时限的增加,凋落物分解速率提高[128]。但随N添加水平的提高,凋落物N、P的分解能力降低[72]。在温带草原,N、P添加对土壤有效N、P及其比值无交互作用,即N添加显著增加土壤无机N含量,对土壤P含量无显著影响,提高了N∶P比值;P添加对土壤N无影响,增加了土壤P含量,降低了N∶P[126]。不同N添加水平土壤C、P保持稳定,土壤有效N在表层增加,在深层减少,但对青藏高原高寒草地C∶N∶P化学计量比值无显著影响[129]。
2.4 人类活动
人类活动加剧超过生态承载力是生态系统退化的直接诱因,在我国典型生态脆弱区,过度放牧、开垦、乱伐和土地利用变化等活动改变了生态系统的结构与功能。放牧主要通过减少植物产量、改变植被组成引起草地生态系统变化,牲畜排泄物则引起养分输入的增加,践踏增加了土壤微生物丰度,造成植物向地下组织再分配[130]。在内蒙古地区,放牧使物种丰富度和地上生物量呈先增后减的“单峰”变化,放牧改变草原生态系统化学计量特征,降低了地上组织和凋落物中的C、N、P库,并加速N循环,降低土壤N含量[131]。在荒漠化地区,过度放牧和人类干扰是导致荒漠化的主要原因,造成土壤C库和养分流失,土壤质量下降,单位土壤资源所能支持的微生物量也降低,随荒漠化程度加剧,土壤微生物量C、N、P和微生物熵均呈降低趋势,即荒漠草地>固定沙地>半固定沙地>流动沙地[96]。而放牧增加了土壤微生物量C∶N、C∶P比值,加剧植物和土壤微生物N、P竞争,长期放牧增加荒漠草原N、P共同限制和草甸草原N限制的可能性[131-132]。同时,开垦直接破坏地表优势植被类型,导致土壤有机C、总N、总P储量的减少,并影响土壤微生物量[133-135]。
随着退耕还林还草、禁牧还草、天然林保护工程等一系列生态恢复工程的实施,生态脆弱区生态环境得到显著改善。研究表明,在禁牧和退耕还林过程中,植被恢复普遍提高了植物、土壤和土壤微生物量C、N含量,土壤C∶N、N∶P也显著增加[16,53,55,85,136-137],而P含量由于主要来源于岩石风化而保持相对稳定[55,84,138]。同时,由于天然林N、P化学计量内稳性优于人工引种林,更能适应本地的环境[139]。总体而言,植被恢复对改善土壤结构和土壤肥力具有积极作用[136,140],并且随着植被恢复,养分逐渐由N限制(恢复前期)向P限制(恢复后期)转变[15,84,138]。
3 研究展望
综上所述,生态化学计量学是生物学、化学、生态学、地理学等多学科交叉研究的前沿领域,经过多年的发展,陆地生态系统生态化学计量学得到快速发展。在生态脆弱区,也形成了大量基于样带调查的多尺度生态化学计量格局研究和基于观测台站的生态化学计量环境变化响应研究。尽管成果丰富,但由于受气候、地形、土壤、生物等地理因子和人类活动的复杂交互作用,生态脆弱区C、N、P化学计量在全球环境变化响应机制、生物地球化学循环过程等诸多问题上并不明确,在未来研究中有待加强。
(1)加强植物-凋落物-土壤-土壤微生物系统的研究。生态化学计量研究从水生生态系统向陆地生态系统转移的过程中,陆地植物与土壤受到广泛关注,也形成了相对丰富的成果。但是作为元素生物地球化学循环的关键环节,凋落物和土壤微生物的研究相对较少,特别是土壤微生物。当前对于元素生态化学计量如何影响凋落物分解、元素生态化学计量在营养链上的转移过程和格局、生态化学计量对环境变化响应的内稳性特征等问题尚不明确。而植物、凋落物、土壤及土壤微生物C、N、P化学计量存在较强相关性[31,132]。因此,将各个要素联系起来形成体系链进行研究对深入了解生态系统养分循环与能量流动、生物地球化学循环具有重要意义。
(2)生态化学计量的尺度依赖。目前,生态化学计量研究主要在物种和种群水平开展,当向群落和生态系统水平拓展时,生态化学计量格局会发生何种变化,在群落内占据不同生态位物种的生态化学计量特征和对环境变化响应的机制,以及生态化学计量和生态系统结构与功能的关系,是未来研究的重要内容。有研究指出,构建生态化学计量生态模型,进行归纳和演绎[18],不失为未来研究的切入点。
(3)开展长期、多因子耦合控制实验。当前生态化学计量对环境变化的响应还存在争议[38,99,102],生态化学计量是受生物、非生物多因素共同作用的结果,基于单因子短期控制实验的研究具有局限性;同时,生物地球化学循环具有一定周期,环境变化对生态化学计量的影响也是缓慢且持续的过程。因此,结合多因子相互作用,开展长期控制实验,是揭示生态化学计量演变机制的途径之一。此外,当前的控制实验多围绕草地生态系统开展,山水林田湖草是生命共同体,在未来研究中阐明这些系统对环境变化的调控机制和协同效应,可以为生态脆弱区生态恢复提供新思路。
