喀斯特地区不同石漠化程度草地根系分解与周转特征
2021-07-12田晓龙汪依妮金宝成何胜江田思惠赵学春
田晓龙,汪依妮,柳 鑫,金宝成,何胜江,田思惠,张 乾,陈 超,赵学春,*
1 贵州大学动物科学学院,贵阳 550025
2 贵州省草地技术试验推广站,贵阳 550025
3 中国科学院植物研究所,北京 100093
根系是植物吸收养分和水分的主要器官,处于不断生长、衰老、死亡和再生长的动态过程中[1]。作为连接植被地上部分与地下部分的介质[2],根系在植物与土壤进行物质与能量交换中起着重要作用[3]。研究表明,细根作为草地生态系统根系的主要部分,仅占地下总生物量的3%—30%,但其周转却消耗了生态系统净初级生产力的10%—75%[1,4- 5]。根系的动态主要表现为生物量、垂直分布、分解和周转的变化,受生态系统类型、气候和土壤等环境因子影响明显[1]。
石漠化是喀斯特地区植被破坏,水土流失,基岩大面积裸露,土地生产力严重下降,地表呈现石质荒漠景观的演变过程或结果[6- 7],已成为制约我国西南地区可持续发展最严重的生态地质环境问题[8-9]。贵州省是中国典型喀斯特山区,具有多相多层复杂界面体系,环境界面变异度高,环境容量低,稳定性差,承灾能力低等特征,属典型的生态脆弱区[10],对外界的干扰表现出不稳定特征[11]。作为石漠化最严重的省份之一,贵州轻度以上石漠化面积为3.59万km2,占国土面积的20.39%;其中,轻度石漠化面积2.27万km2,中度石漠化面积1.05万km2,强度石漠化面积0.27万km2;处于潜在石漠化趋势的土地面积为4.37万km2[12]。
草地作为贵州喀斯特地区主要植被类型之一,根系对环境条件的反应敏感[13],决定着植物的生长发育和土壤营养状况,其动态对环境变化亦有指示作用,可反映草地生态系统的健康状况[14]。目前,对喀斯特地区根系的研究主要集中于植被恢复过程中根系的形态特征[15-16]、生物量、分布[17-19]和碳储量[20]等方面,而对草地根系研究尤其根系分解与周转的研究报道较少。不同石漠化程度草地在植被、土壤条件等方面均呈现出极大差异,是否预示着地下根系,尤其是根系生物量、分解和周转率是否也会发生相应变化?引起这种变化的原因又是什么?本文旨在通过野外调查,比较分析不同石漠化程度草地根系的生物量、分解和周转及相应的环境因素,探讨不同石漠化草地根系动态变化机理,为更深入地了解喀斯特地区草地养分循环过程,恢复石漠化脆弱生态系统和草地经营提供科学依据。
1 材料与方法
1.1 研究区概况
贵州省位于我国西南地区,介于24°37′—29°13′N,103°36′—109°35′E之间,属中亚热带季风湿润气候,雨热同期,年平均降雨量1000—1300 mm,平均气温15 ℃,植被类型丰富,碳酸盐岩分布广泛,主要为石灰岩和白云岩,92.5%的面积为山地和丘陵[21]。土壤类型以黄壤、红壤和黄棕壤为主。现有各类天然草地面积达4.28 × 104km2,占贵州省土地总面积的24.3%[22]。维管植物203科1025属4725种(包括变种),以禾本科、豆科、菊科、莎草科植物为主要优势种和建群种。
本研究选取3种不同石漠化程度草地(潜在、中度和强度)作为研究对象,其中,潜在石漠化草地位于黔南州独山县上司镇,年降水量1429.9 mm,年平均温度15.9 ℃,土壤类型以黄壤为主;中度石漠化草地位于毕节市大方县星宿乡,年降水量1155.1 mm,年平均温度11.8 ℃,土壤类型以黄壤为主;强度石漠化草地位于安顺市关岭县普利乡,年降水量1205.1—1656.8 mm,年平均温度16 ℃,土壤类型以黄壤为主。基本情况见表1。

表1 三种不同石漠化程度草地的基本情况
1.2 试验设计
在上述每个草地内分别设置3个25 m × 25 m的样方。沿样方对角线选取3个样点,于2017年3月至2018年1月,每隔2月进行土壤样品采集,采集时去掉地表凋落物,利用土钻依次分别钻取0—10 cm、10—20 cm、20—30 cm土层土样约300 g,用于测定土壤理化指标(含水量、pH值、电导率等)。并在用环刀(内径50.46 mm,高50.02 mm)垂直采样测量上述土层容重,采用土壤温度计测定土壤温度。
同时,沿样方对角线选取3个50 cm × 50 cm的采样点,利用连续土柱法垂直挖取样点内0—10 cm、10—20 cm、20—30 cm土层的土样。将采集的土样置于细筛上用水冲洗,仅留干净的根系,根据根系的颜色、弹性、表皮是否存在区分死根系与活根系,于65 ℃下烘干至恒重后称重,根据每次采样所得死根系与活根系的生物量计算根系年死亡量、根系年生产量、死根系现存量及活根系现存量。
采用尼龙网袋法进行根系分解实验[23]。于2017年3月,称取上述处理好的根系3 g与样地内10 cm深的磨碎土壤混匀,装入尼龙袋(长10 cm,宽15 cm,孔径0.2 mm)中,埋入10—15 cm深的土层中,每个样地放置25袋。每隔2月从各样地取5个分解袋,带回实验室,去除杂质,转移置信封,置于65 ℃下烘干至恒重后称重,计算根系分解率。
1.3 室内分析
土壤样品置于干燥阴凉处风干,过0.15 mm孔径土壤筛,用pH计和DDSJ- 308型电导仪测定土壤pH值和电导率,采用K2Cr2O7外加热法测定土壤有机碳含量,凯氏定氮法测定土壤全氮含量,钼锑抗比色法测定土壤全磷含量,火焰光度计法测定土壤全钾含量。
1.4 数据处理
根系周转率计算方法如下:

M=Mmax-Mmin+D
P=Pmax-Pmin+M
式中,vi为第i次(i=1,2,3,4,5)取样根系的分解速率;di为第i次取样死根系的生物量;M为根系年死亡量(g m-2a-1);P为根系年生产量(g m-2a-1);D为根系年分解量(g m-2a-1);T为根系周转速率(次/a);Mmax为死根系现存量的最大值;Mmin为死根系现存量的最小值;Pmax为活根系现存量的最大值;Pmin为活根系现存量的最小值;Y为活根系现存量的平均值。
应用Olson指数衰减模型对根系分解速率进行拟合[24]:
Ft=F0e-kt
式中,t是分解时间,F0为起始时根系重量,Ft为时间t时根系残留重量,k是根系分解速率系数。
基础数据利用Excel进行整理归类;采用SPSS 20.0进行方差分析和逐步回归分析;利用SigmaPlot 14.0软件作图。图表中所有数据均表示为平均值±标准差(Mean±SD)。
2 结果分析
2.1 土壤理化性质和土壤养分
不同草地类型的土壤理化性质指标见表2。土壤容重、温度、电导率和pH等土壤理化性质指标在3种不同石漠化程度草地中具有显著地差异,随着石漠化程度的增加,土壤温度、电导率和pH均表现为先降低后增加的趋势(P<0.05),而土壤含水量变化不显著(P>0.05)。其中,中度石漠化草地的土壤温度、土壤电导率和土壤pH均显著低于潜在和强度石漠化草地。

表2 三种不同石漠化程度草地的土壤理化性质
不同草地类型的土壤养分情况见图1。土壤全氮、全磷和全钾等土壤养分指标在不同石漠化程度草地均表现为差异显著(P<0.05)。其中,强度石漠化草地的土壤全氮和全磷显著高于潜在和中度石漠化草地(P<0.05);潜在石漠化草地的土壤全钾显著高于中度和强度石漠化草地(P<0.05);各草地类型的土壤有机碳差异不显著(P>0.05)。

图1 三种不同石漠化程度草地的土壤养分
2.2 根系生物量分布和季节动态
不同草地类型的根系生物量垂直分布特征表明(表3),潜在、中度和强度石漠化草地各土层的根系生物量差异显著(P<0.05),随着土层的增加,均表现为逐渐降低的趋势。其中,潜在、中度和强度石漠化草地的根系生物量最大值均出现在0—10 cm土层,0—10 cm占0—30 cm土层根系生物量的比例分别为57.66%、81.02%和72.18%。
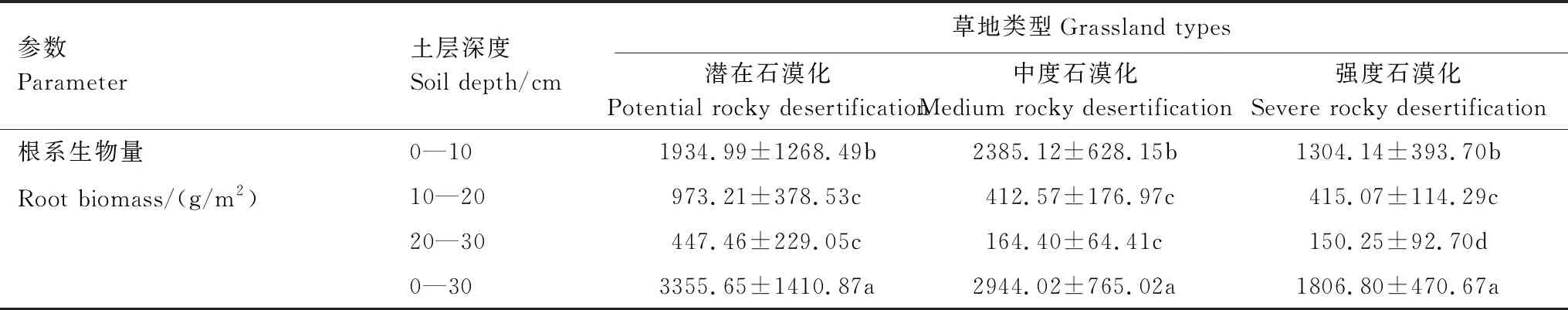
表3 三种不同石漠化程度草地根系生物量的垂直分布
不同草地类型根系生物量的季节变化见图2。潜在、中度和强度石漠化草地根系生物量月平均值分别为3355.65、2944.02、1806.80 g/m2。潜在和中度石漠化草地根系生物量均呈现先增加后降低的趋势,在9月达到最大值,分别为6051.44、3591.38 g/m2;强度石漠化草地群落根系生物量无明显的变化(P>0.05)。

图2 三种不同石漠化程度草地根系生物量的季节变化
2.3 根系分解和周转
不同草地类型的根系分解速率随时间的变化规律见图3,潜在、中度和强度石漠化草地的根系分解速率随时间变化明显。分解初期较大,60 d后逐渐降低,在180 d时下降至最小值,随后逐渐增大。

图3 三种不同石漠化程度草地根系分解速率随时间的变化
不同草地类型的根系分解残留量随分解时间的变化表明(表4),潜在、中度和强度石漠化草地根系的年分解速率分别为64%、56%和58%。当根系分解量达到50%时,潜在石漠化草地需要173 d,中度石漠化草地需要304 d,强度石漠化草地需要233 d;当根系分解量达到95%时,潜在石漠化草地需要1174 d,中度石漠化草地需要1351 d,强度石漠化草地需要1878 d。
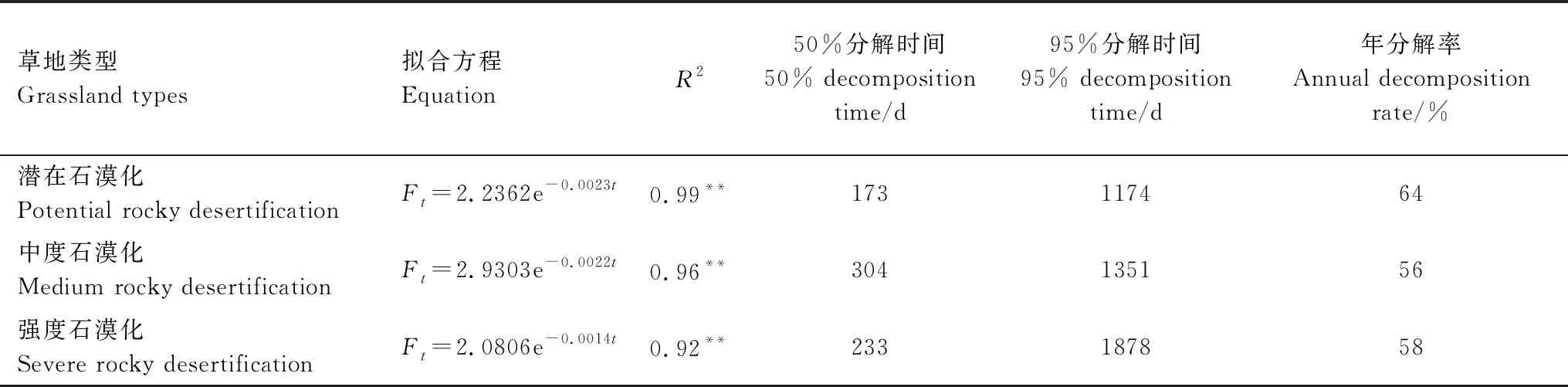
表4 三种不同石漠化程度草地根系分解残留量随分解时间的变化
不同草地类型根系的年分解量、死亡量、生产力和周转速率如表5所示。潜在、中度和强度石漠化草地根系的年分解量、年死亡量、年生产力和周转速率差异变化显著(P<0.05),随着石漠化程度的增加,根系年死亡量、年生产力和周转速率逐渐降低。其中,潜在石漠化草地的根系年死亡量分别为中度石漠化草地、强度石漠化草地的1.40、2.46倍;潜在石漠化草地的根系年生产力分别为中度石漠化草地、强度石漠化草地的1.61、3.83倍;潜在石漠化草地的根系周转速率分别为中度石漠化草地、强度石漠化草地的1.24、1.94倍。根系年分解量最大的是中度石漠化草地,为强度石漠化草地的2.65倍及潜在石漠化草地的3.36倍。

表5 三种不同石漠化程度草地根系年分解量、死亡量、生产力和周转速率
2.4 根系动态与土壤因子的关系
不同草地类型的根系分解速率与土壤有机碳、全氮、全钾、全磷等土壤养分因子的逐步回归分析表明(表6),影响潜在石漠化草地根系分解速率的主要因子是土壤全氮;影响中度石漠化草地根系分解速率的主要因子是土壤全钾、土壤全氮和土壤有机碳;影响强度石漠化草地根系分解速率的主要因子是土壤有机碳

表6 三种不同石漠化程度草地根系分解速率与土壤养分的逐步回归分析
不同草地类型的根系分解速率与土壤物理性质的逐步回归分析表明(表7),影响潜在石漠化草地根系分解速率的主要因子是土壤电导率;影响中度石漠化草地根系分解速率的主要因子是土壤含水量;影响强度石漠化草地根系分解速率的主要因子是土壤温度、土壤含水量和土壤电导率。

表7 三个石漠化草地根系分解速率与土壤物理性质的逐步回归分析
3 讨论
3.1 根系生物量对石漠化响应
根系生物量的季节动态能够揭示草地植物的生长状况、土壤理化现状及根系周转快慢等特征。研究认为根系生物量在1年中常出现1个[25]或2个[26]峰值,峰值出现在春季、晚夏或秋季等,受植物自身特性及外界环境条件的影响,存在一定的波动[27]。本研究结果表明,潜在、中度石漠化草地的根系生物量均在9月份最大,峰值出现在晚夏,与Du等[25]对西南喀斯特地区植被恢复过程中根系生物量的研究结论相似,其原因是此阶段土壤养分有效性较高,温湿条件较好[28],利于植物的生长发育,根系生物量较高。
喀斯特地区土壤具有高度的空间异质性,不同土层间养分差异明显[29]。土壤空间异质性是导致根系分布空间差异的主要原因[30],石漠化程度越剧烈,土壤越瘠薄且土壤持水力越差,草地根系生长和分布的越具有特殊性。不同石漠化程度的草地根系生物量分布均随土层深度增加而逐渐减少,这与其他研究结果基本一致[18,25]。根系在0—10 cm土层的生物量占0—30 cm土层总生物量的57%以上,且10—30 cm土层的根系生物量急剧下降,可能与喀斯特特殊的地貌结构有关。研究区属亚热带季风气候区,水热资源丰富但分布不均,岩石裸露率高,土层浅薄且不连,二元结构明显,湿润气候条件下的特殊岩溶干旱重[18],水分成为限制因子。因此,喀斯特地区植物常具有发达而强壮的根系,通过攀附岩石、穿透裂隙,吸收储存在岩石空隙、裂隙中的水分和养分,发展高密度的浅层根系提高浅层水分利用效率,以适应特殊的环境[31-32],从而喀斯特植物根系发育具有明显的表聚现象[18]。另一方面,3个群落的建群种均为禾本科植物,其根系属浅根性根系,加之石漠化的土层相对较浅,有机质和养分都储存于土壤表层,根系为获取更多的资源而趋于浅层分布。
3.2 根系分解和周转对石漠化的响应
根系分解主要是在淋溶、破碎等物理作用下,伴随土壤生物代谢对化学元素的吸收和释放,不断与外界环境进行物质交换的过程[33],其分解速率主要受土壤的理化性质、环境条件和根系本身的化学特性影响[34]。逐步回归分析表明在生态环境脆弱的石漠化地区,土壤水分和养分是其生态系统功能实现的主要限制因子,根系分解和周转作为养分循环的关键环节受水分影响明显,土壤养分亦成为根系分解和周转的间接限制因子。3种不同石漠化程度草地的根系分解均表现为“快—慢—快”的变化特征,与前人的研究结果相似[35- 36]。主要因为分解初期受非生物作用的影响较大,根系中碳水化合物等可溶解物质快速淋溶;分解后期主要受生物作用影响,随着分解进行,大多数可溶性化合物消耗,木质素、纤维素等难溶物质残留下来,由微生物缓慢降解[34]。另一方面,根系年分解量在中度石漠化草地中显著高于潜在和强度石漠化草地(表5,P<0.05),可能是因为典型喀斯特石漠化生态系统土壤随着石漠化程度增加是一个先退化后改善的趋势[37];土壤养分缺乏时,植物会显著提高根系生物量在总根系中的比例[38],以提高根系对土壤资源的获取能力,加快根系周转,从而促进根系分解。结合表1和图1可知,中度石漠化草地的土壤情况较潜在、重度石漠化草地差,中度石漠化草地根系通过快速分解以弥补土壤养分的缺乏。
根系周转主要是植物为了适应环境的变化,需发育新根系代替老根系以保证对土壤水分和营养物质的吸收,从而保持植物生长和生命活动的继续[39]。Jackson等分析表明,植物的细根周转速率在0.02—2.64次/a之间,平均周转速率为0.56次/a[40]。本研究中潜在、中度和强度石漠化草地的根系周转率分别为1.70次/a、1.36次/a和0.87次/a,处于Jackson等报道的周转速率范围内,但高于其研究的平均周转速率。主要因为随着石漠化等级不断增加,裸露岩石聚集效应逐渐明显,汇集大气沉降养分和岩溶产物,增加了土壤氮素和有机物的输入;同时随着石漠化程度增加,流失的土壤越来越少,导致水土流失越来越弱,土壤养分流失微弱,强度石漠化环境土壤养分和物理性能得到了明显的改善[37]。因此,较高的根系周转速率可能是由于石漠化环境下的裸露岩石聚集效应,使土壤环境得到改善从而加速了根系的周转,以利于植物适应石漠化环境。另一方面,3种石漠化草地群落建群种和优势种均为禾本科植物,根系为须根系,相关研究表明植物根系直径越小,比根长和比根面积越大,与土壤微生物接触机会就更多,被微生物分解利用的可能就更大[41],因此周转速率较高。也有研究报道表明,植物维持相对较高的根系生物量需要消耗大量的营养物质和能量,保持较高的根系周转率可以以较低现存根系生物量满足植物旺盛的生命活动对水分和营养物质的需求,是植物降低能耗的一种适应方式[42]。
