喀斯特山区土地利用方式转变对土壤酶活性及其化学计量特征的影响
2021-07-12孙彩丽王艺伟王从军黎庆菊吴志红袁东昇张建利
孙彩丽,王艺伟,王从军,黎庆菊,吴志红,袁东昇,张建利
1 贵州民族大学生态环境工程学院,贵阳 550025
2 紫云苗族布依族自治县林业局,贵州紫云苗族布依族自治县 560800
土壤胞外酶主要来源于土壤微生物的活动、植物根系分泌物和腐解的动植物残体,通过催化、分解、转化及合成等过程参与土壤生物地球化学循环,是土壤生态系统代谢的一类重要动力,在很大程度上反映土壤物质循环与转化的强度,是重要的土壤肥力指标[1- 3]。水解酶是极为重要的一类胞外酶, 它可以催化土壤中的高分子和其它难利用物质水解为植物、微生物可利用的营养物质, 在物质循环和能量流动中的作用不容忽视[4]。关于土壤胞外酶的研究多以参与碳循环的β- 1,4-葡萄糖苷酶(BG)、β- 1,4-木糖苷酶(BX)和纤维素二糖水解酶(CBH),参与氮循环的β- 1,4-N-乙酰葡萄糖苷酶(NAG)和亮氨酸氨基肽酶(LAP),和参与磷循环的酸性磷酸酶(AP)为研究对象,因为这些酶主要参与末端的催化反应,可以反映土壤的碳氮磷代谢水平[5- 7]。大量研究显示,研究区域、地形条件和人为活动等均显著影响土壤胞外酶活性[3,8- 9],说明胞外酶能灵敏地反映自然条件和人类活动影响下土壤生物化学过程的方向和强度,因此,研究土壤胞外酶活性对预测土壤生态系统变化具有重要意义。
生态化学计量学以定量的方式明确反映营养物质分解、补给速率的快慢以及土壤质量状况等生态系统动态变化和能量平衡过程,是研究碳、氮和磷等元素在生态过程中耦合关系的有利工具[10- 11]。近年来,随着生态化学计量研究的深入, 土壤酶的生态化学计量特征日益受到科学家们的重视,主要研究与元素(主要是碳氮磷元素)循环相关的酶活性之间的比值[12]。酶化学计量比能更好地反映微生物的代谢和营养需求之间的生化平衡,是表征土壤能量和养分资源限制状况的重要指标[13]。通过meta分析发现,全球尺度上参与碳氮磷循环的土壤酶化学计量比ln(BG)∶ln(NAG+LAP)∶ln(AP)趋于1∶1∶1[14],意味着全球尺度上碳氮磷相关代谢酶相对稳定。然而,在特定的生态系统中,土壤微生物的生长受到环境胁迫、养分限制及地上植被等生物因子的影响,进而影响酶的表达和活性,土壤酶化学计量比可能不遵循 1∶1∶1 的关系。Xu等对我国不同区域森林土壤研究表明,随着 pH 增大,ln(BG+CBH)∶ln(AP) 和ln(NAG+LAP)∶ln(AP) 呈减小趋势[15]。Peng 等研究发现我国温带草原全磷与ln(NAG+LAP)∶ln(AP) 呈显著正相关[16]。高秋雨等研究发现亚热带人工林根际土壤酶活性及计量比受植被种类、林分类型及取样季节影响,且基质有效性在其中发挥重要作用[4]。为了准确评估土壤生态系统养分周转状况,具体分析特定环境条件和研究背景下土壤酶的化学计量特征显得尤为重要。
我国西南喀斯特区域景观异质性强,土壤瘠薄且不连续,水土流失和石漠化严重,具有明显的脆弱性和易损性,破坏容易恢复难[17],因而在该地区进行植被恢复和生态环境建设显得极为重要和紧迫。在国家“退耕还林还草”和“生态环境重建”等政策的推动下,西南喀斯特地区开展了大规模的以土地利用结构调整为主的综合治理措施,许多坡耕地转变为林地、梯田等其它土地利用方式[18]。实践表明,土地利用结构调整不仅控制了水土流失和石漠化发展,还有效地改善了土壤性质和质量状况[19]。遗憾的是,喀斯特山区由土地利用方式改变引起的土壤质量变化过程及驱动机制还不十分清楚。土壤酶作为反映土壤质量高低的重要生物学指标,参与土壤生物化学过程和物质循环,对土壤有机质转化起重要作用[20],因此,研究喀斯特地区土壤酶活性及其化学计量特征随土地利用方式转变的变化规律及其与土壤养分间的内在相关性有利于解决上述问题。
贵州省具有典型的喀斯特地貌特征,本研究以贵州省西南部的紫云县为研究区,选择马尾松林地、柏树林地、灌木林地、梯田菜地、梯田撂荒地、坡面撂荒地和坡耕地等7种典型的土地利用方式为研究对象,通过分析不同土地利用方式土壤养分、酶活性及化学计量比的差异特征,探究环境因子与土壤酶活性之间相关性,以期揭示喀斯特山区土地利用方式改变引起的土壤养分变化特征及内在驱动机制,为当地生态恢复与重建提供参考及理论依据。
1 材料与方法
1.1 研究区概况
研究区位于贵州省安顺市紫云县境内(105°55′—106°29′E,25°21′—26°3′N),平均海拔1000—1300 m。属亚热带温暖湿润气候,四季分明,水热同季,年平均气温约为15.3℃,年均降雨量1337 mm。山多地少,80% 为 25°以上坡地,土壤以黄壤为主,土层浅薄,山地平均土层深度20—40 cm。研究区人地矛盾突出,人为干扰强烈,植被退化严重,森林覆盖率低且以次生林和人工林为主,长年水土流失导致石漠化趋势明显,生态环境十分脆弱。在国家退耕还林(草)政策驱动下,大部分坡耕地转变为其它土地利用方式,现存典型的土地利用方式主要有坡耕地、梯田、林地、撂荒地等。
1.2 样品采集
2019年9月选取7种当地常见的土地利用方式采集土壤样品,包括马尾松林地、柏树林地、灌木林地、梯田菜地、梯田撂荒地、坡面撂荒地和坡耕地。各土地利用方式转换前均为坡耕地,转换时间在2005年前后。马尾松林和柏树林为人工林,灌木林地主要灌木为火棘,梯田菜地种植蔬菜为辣椒,梯田撂荒地和坡面摞荒地优势植物有马尾松(PinusmassonianaLamb.)、亮叶桦(BetulaluminiferaH. Winkl.)和火棘(Pyracanthafortuneana(Maxim.) Li)等,坡耕地的种植作物为玉米。
采集土壤样品时,7种土地利用方式的坡度和坡向等立地条件尽量保持一致,每种土地利用类型设置3个重复样地(样地面积为 100 m×100 m 左右),每个样地相距 100 m 左右,样地内“S”型设置 9个采样点,每个采样点间距10 m以上,取样深度为 0—20 cm。对每个样地内采集的土壤样品去除枯枝落叶后进行充分混匀,采用四分法留取 1 kg 土壤,代表该样地的土壤样品,本研究共得到21个土壤样品。将采集到的鲜土立即带回实验室,过2 mm筛后分两部分,一部分立即放4℃冰箱保存,用于土壤酶活性的测定,另一部分风干研磨过0.25 mm筛后用于养分和pH的测定。
1.3 土壤样品分析
1.3.1土壤酶活性测定
土壤酶活性采用96微孔板荧光法进行测定[21],共6种水解酶,分别是β- 1,4-葡糖苷酶(BG)、β- 1,4-木糖苷酶(BX)和纤维素二糖水解酶(CBH)、β- 1,4-乙酰-葡糖胺糖苷酶(NAG)、亮氨酸氨基肽酶(LAP)和酸性磷酸酶(AP)。各种酶的反应底物、缩写和功能如表1所示。具体测定方法为:称取4℃保存的鲜土1 g于250 mL 广口瓶中, 加入125 mL醋酸缓冲液(50 mmol/L, pH约5.0), 震荡 1 h制成土壤悬浮液。然后,使用八通道移液器吸 200 μL土壤悬浮液于96微孔板内,每个样品孔中加 50 μL的底物(200 μmol/L, 超纯水配制)。同时做样品和基质控制,样品控制孔加200 μL 土壤悬浮液和 50 μL 醋酸钠缓冲液,基质控制孔中加200 μL醋酸钠缓冲液和50 μL底物。所有样品和空白加完后,将微孔板放在25 ℃的黑暗条件下培养4 h,之后分别在每个孔内加10 μL NaOH溶液(1 mol/L)终止培养。1 min后, 用多功能酶标仪(Synergy H4, BioTek, Winooski, USA)在365 nm波长处激发, 450 nm波长处进行荧光测定。酶底物均为Sigma公司生产, 其他试剂为国产分析纯。
1.3.2土壤化学性质测定
土壤养分和pH采用传统方法测定[22]:土壤有机碳(SOC)采用重铬酸钾氧化外加热法测定;土壤全氮采用半微量凯氏定氮法测定;全磷采用 NaOH 碱溶-钼锑抗比色法测定;有效磷采用 NaHCO3浸提-钼锑抗比色法测定;土壤pH值采用电位法测定,土壤溶液采用水(去CO2蒸馏水)浸提法调制(水土比 2.5∶1)。
1.4 数据分析
利用土壤酶化学计量的向量分析预测土壤养分循环限制因素,计算公式如下[23]:
X=(BG+CBH+BX)/[(BG+BX+CBH)+AP]
Y= (BG+BX+ CBH)/[(BG+CBH+BX)+(NAG+LAP)]
矢量长度=SQRT(X2+Y2)
矢量角度=DEGREES((ATAN2(X;Y))
其中,矢量线段的长度用以表示能量相对于养分的限制,矢量线段与X轴的夹角(矢量角度)用以表示磷元素相对于N元素的限制程度,矢量角度大于45°表示磷限制,小于45°表示氮限制。
文中数据以平均值加标准差的形式进行呈现。采用单因素方差分析法和多重比较法(Duncan)分析不同土地利用方式土壤养分、酶活性及化学计量特征的差异。采用冗余分析(RDA)探索土壤养分和酶活性及化学计量比之间的关系,在进行RDA分析时,首先通过方差膨胀因子(VIF)判断解释变量间的共线性程度,若VIF大于10证明存在共线性问题,需要剔除存在共线性关系的变量,再进行RDA分析,并对分析结果进行蒙特卡罗检验。另外,对不同土地利用方式的土壤养分含量和胞外酶活性进行聚类分析,将土地利用方式进行归类。聚类分析、RDA和VIF计算分别基于R语言中的vegan和packfor程序包,作图基于ggplot2完成。其余统计分析在SPSS 18.0中完成。
2 结果与分析
2.1 不同土地利用方式土壤养分含量及化学计量比特征
不同土地利用方式土壤养分含量和化学计量比差异显著(表2)。土壤有机碳含量表现为梯田菜地最高,柏树林地、马尾松林地、梯田撂荒地和坡面撂荒地次之,灌木林地和坡耕地最低;土壤全氮含量表现为柏树林地最高,坡耕地最低,其它土地利用方式的全氮含量居中;土壤全磷含量表现为柏树林地>马尾松林地和梯田撂荒地>灌木林地和梯田菜地>坡面撂荒地和坡耕地;只有马尾松林地和柏树林地的速效磷含量显著高于坡耕地。与坡耕地相比,梯田菜地的C∶N值显著增大而柏树林地和马尾松林地的C∶N值显著减小,而所有土地利用方式C∶P和N∶P值较坡耕地都显著减小。

表2 不同土地利用方式土壤理化性质
2.2 不同土地利用方式土壤胞外酶活性差异
图1结果显示,不同土地利用方式BG活性表现为柏树林地、梯田菜地和梯田撂荒地显著高于马尾松林地、灌木林地和坡面撂荒地显著高于坡耕地;所有土地利用方式的CBH活性都显著高于坡耕地且柏树林地CBH活性最高;只有柏树林地、灌木林地和梯田菜地的BX活性显著高于坡耕地。柏树林地和梯田撂荒地的NAG活性显著高于坡耕地;除坡面撂荒地外,所有土地利用方式的LAP活性显著高于坡耕地。与坡耕地AP活性相比,所有土地利用方式AP活性显著增加,且最大值出现在马尾松林地和柏树林地。

图1 不同土地利用方式土壤胞外酶活性差异
由于BG、CBH和BX都属于代谢碳的酶,NAG和LAP属于代谢氮的酶,AP属于代谢磷的酶,将其归类后结果显示,柏树林地、梯田菜地和梯田摞荒地的碳酶活性显著高于坡耕地,其它土地利用方式的碳酶活性较坡耕地差异不显著。氮酶活性表现为只有柏树林地、灌木林地和梯田摞荒地显著高于坡耕地。所有土地利用方式的磷酶活性都显著高于坡耕地,且最大值出现在马尾松林地和柏树林地。
2.3 不同土地利用方式土壤胞外酶化学计量比和矢量特征
不同土地利用方式土壤酶化学计量比和矢量特征差异显著(表3)。梯田菜地和坡面撂荒地的酶C∶N显著高于坡耕地,其它土地利用方式的酶C∶N较坡耕地差异不显著。酶N∶P表现为坡耕地>梯田菜地>灌木林地、柏树林地和梯田撂荒地>马尾松林地和坡面撂荒地。除灌木林地外,其它土地利用方式的酶N∶P均显著小于坡耕地。酶活性矢量长度介于0.547—0.797之间,所有土地利用方式的酶活性矢量角度均大于45℃。此外,不同土地利用方式碳氮磷酶活性比的平均值分别为1.251、0.505和0.459,而矢量长度和矢量角度的平均值分别为0.614和58.507。用ln(BG+CBH+BX)∶ln(NAG+LAP)∶lnAP表示土壤酶C∶N∶P化学计量比,则不同土地利用方式土壤酶化学计量C∶N∶P总体上为1∶1∶1.3。
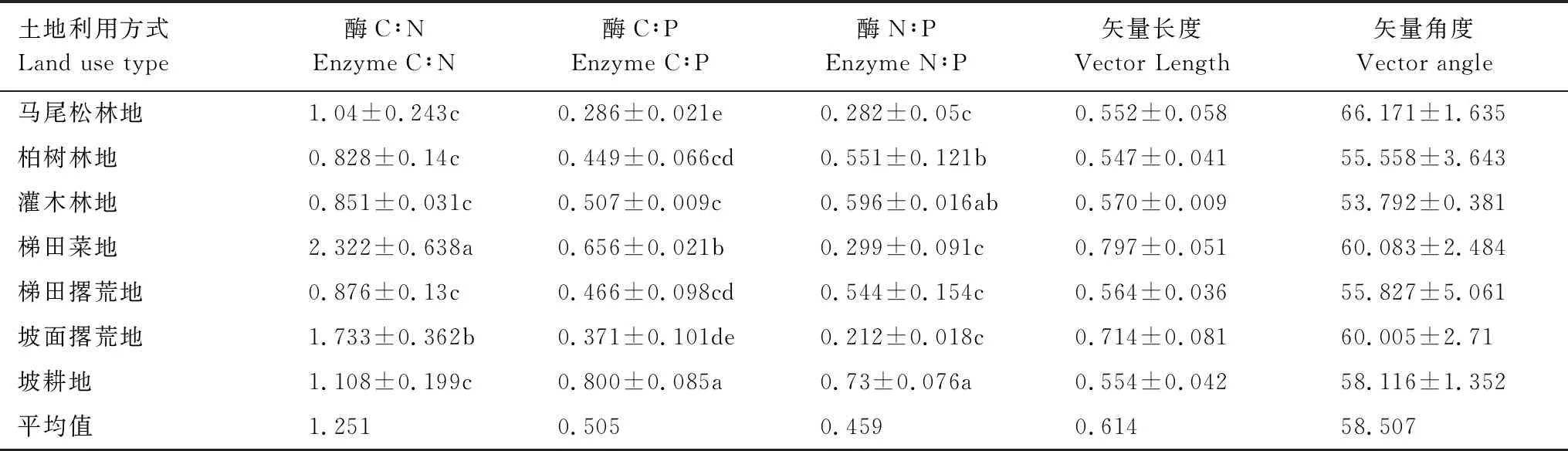
表3 不同土地利用方式土壤酶活性的化学计量比和矢量特征

图2 土壤酶活性及计量比与土壤理化性质关系的冗余分析(RDA)
2.4 土壤环境因子与胞外酶活性之间的相关性
对8种土壤环境因子与酶活性之间的相关性进行冗余分析(RDA),结果显示前两个排序轴共解释了土壤酶活性及计量比变异的55.72%(结果未呈现)。剔除存在共线性关系的变量后,剩余全氮、速效磷、C∶N、N∶P和pH共5个因素,再进行RDA分析,结果显示前两个排序轴共解释了变异的50.22%(RDA1:29.91%和RDA2:20.31%)(图2)。蒙特卡罗检验结果显示,在被检验的5个土壤化学指标中,除速效磷外,都对土壤酶活性及其化学计量比具有显著影响。从图中可以看出,RDA第一轴与氮磷代谢酶(NAG、LAP和AP)关系密切,且沿RDA1增大的方向土壤中全氮含量增加,pH和N∶P减小,RDA第二轴与碳代谢酶(BG、CBH、BX)和酶化学计量比关系密切,且沿RDA2增大的方向土壤中C∶N减小。
2.5 不同土地利用方式土壤化学性质和酶活性的聚类分析
对不同土地利用方式的土壤养分含量、pH和胞外酶活性进行聚类分析,结果显示:不同的土地利用方式的土壤可清晰地分为4组,灌木林地、梯田撂荒地和坡面摞荒地为一类,马尾松林地和柏树林地为一类,梯田菜地和坡耕地各为一类(图3)。

图3 不同土地利用方式土壤养分和酶活性的聚类分析
3 讨论
3.1 土地利用方式转变对土壤养分含量和酶活性的影响
土地利用是自然条件和人为活动的综合反映,转变土地利用方式是人类干预土壤质量最直接的活动[24],通过影响土壤有机物的输入、水分和养分迁移和微生物活动等[25],进而改变土壤养分循环强度、总量和路径[26],最终导致土壤质量发生变化[3,17]。本研究发现喀斯特坡耕地转变为林地、梯田或撂荒地后,土壤有机碳、全氮和全磷含量明显增加,且马尾松林地和柏树林地的速效磷含量显著增加。这与前人研究结果基本一致[19,27- 28],表明对喀斯特坡耕地进行植被恢复和生态重建可有效改善土壤的养分状况。主要原因在于坡耕地主要种植玉米等粮食作物,在人为干预下土壤养分消耗过多但地表枯落物归还量小,导致土壤养分相对贫瘠[3]。然而,坡耕地撂荒或转变为林地和梯田后地上植被得以恢复,群落结构发生巨大变化[29],直接增加了土壤表层枯枝落叶输入量和地下根系生物量,土壤微生物将输入的有机物分解为无机养分归还土壤,从而增加了养分含量,改善了土壤质量状况[25,30]。
土壤酶通过催化、降解、转化及合成作用参与有机质的转化和生物地球化学循环,是反映土壤肥力的重要生物学指标[20]。本研究结果表明对喀斯特坡耕地进行土地利用转换后土壤BG、CBH、LAP和AP活性普遍提高,而BX只在转化为柏树林地、灌木林地和梯田菜地后显著提高,NAG只在转化为柏树林和梯田摞荒地后得到显著提高。对土壤酶进行归类后,发现坡耕地转变为柏树林地、梯田菜地和梯田撂荒地之后碳代谢酶活性显著提高,转换为柏树林地、灌木林地和梯田摞荒地后氮代谢酶活性显著提高,转换为林地、撂荒地和梯田后磷代谢酶活性普遍增加,且马尾松林地和柏树林地磷代谢酶活性的增加幅度最大。土壤酶主要来源于土壤微生物、植物根系、土壤动物的分泌物及其残体的分解物等,并且一般认为水解酶主要负责分解土壤中的活性有机质[12,31],因此,本研究中土地利用方式改变后土壤胞外酶活性的提高与土壤中活性有机物的增加关系密切,比如枯落物和根系分泌物等,它们作为土壤微生物养分的主要来源,在很在程度上刺激土壤微生物胞外酶的分泌[32- 33]。此外,土壤理化属性也会对土壤酶活性产生影响,已有研究表明,温度、水分、pH、养分有效性和真菌/细菌比都会在一定程度上影响土壤酶活性[34- 35]。本研究中,坡耕地转变为林地、梯田和撂荒地后,土壤有机碳、全氮和全磷含量明显增加,马尾松林地和柏树林地的速效磷含量显著增加。并且,马尾松林、柏树林地、灌木林地和撂荒地的地上植被覆盖率较坡耕地有很大提高,植物根系的发达程度和凋落物归还量也有很大改善。梯田一方面因为截留作用土壤的水肥条件较比坡耕地优越,另一方面梯田菜地因为人为施入农家肥土壤有机质含量会明显提高。这些条件导致土壤活性有机物输入量和土壤理化属性较坡耕地明显好转,进而引起土壤酶活性显著提高。
3.2 不同土地利用方式土壤养分限制因子
土壤C∶N∶P化学计量特征是表征土壤养分平衡、矿化和固持作用的重要指标,反映土壤养分组成及质量状况[36]。本研究中,土壤 C∶N、C∶P和N∶P的平均值分别为9.58、207.174和21.228,并且,与坡耕地相比,梯田菜地C∶N显著增大,柏树林地和灌木林地的C∶N显著减小,其它所有土地利用方式C∶P和N∶P都显著减小。Tian等研究发现我国陆地表层土壤C∶N、C∶P和N∶P分别接近14.4、136 和9.3[8]。相比之下,本研究的C∶N 偏低而C∶P和N∶P偏高,说明研究区表层土壤有机质分解过程中受 P 素限制。田静等[36]和刘立斌等[10]对喀斯特土壤 C/N/P 化学计量特征的研究支持了我们的结果,证明喀斯特表层土壤普遍存在缺磷现象。坡耕地转梯田菜地后C∶N显著增大主要是因为农家肥的施入导致土壤有机碳含量大幅增加,柏树林地和灌木林地C∶N显著减小的主要原因是土壤总氮含量的显著增加。坡耕地转换为林地、摞荒地和梯田后,土壤碳氮磷含量总体呈增加趋势,然而,这些土地利用方式的C∶P和N∶P均显著减小,说明对喀斯特坡耕地进行土地利用方式转换不仅有助于土壤养分含量的提高,还促进了养分的平衡,增加了磷元素的有效性。
喀斯特土壤的营养状况相对贫瘠,根据资源分配理论[37],土壤微生物可通过调节酶的分泌,提高限制元素的利用性,从而缓解土壤养分的限制[38],因此,研究土壤酶的化学计量特征对土地利用变换的响应规律,有助于探究土壤微生物对养分限制的应对策略。本研究结果显示坡耕地转梯田菜地和坡面撂荒地后,酶C∶N显著增加而酶C∶P和酶N∶P显著减小,转马尾松林地、柏树林地和梯田撂荒地后酶C∶P和酶N∶P显著减小,转灌木林地后酶C∶P显著减小。说明坡耕地转其它土地利用方式后土壤N素限制得到了缓解,但加重了土壤P的限制。转变土地利用方式后,植被得以恢复,地上植物的快速生长促使N、P元素由土壤向植物大量转移,由于土壤中的P元素属于沉积型元素,主要来源于矿物岩石的缓慢风化作用[39],而N元素主要来源于微生物的对凋落物分解作用[40],土壤P素比N素更难得到补给,因此植物对养分的吸收更易导致土壤P素的减少,从而加重土壤中P的限制。此外,随土地利用方式转换时间的增加,地上植物因存在对营养元素的再吸收现象,枯落物中磷含量增加导致C∶P降低,当其进入土壤以后,土壤微生物需要提高磷循环相关酶的活性以降解归还的枯落物[41]。
经过对数转化后,本研究土壤酶化学计量比C∶N∶P为1∶1∶1.3,偏离了全球生态系统1∶1∶1的比值[14],表明研究区域内所有土地利用方式表层土壤具有较高的P转化酶活性,意味着研究区内土壤P素相对缺乏。并且,所有土地利用方式土壤酶活性的矢量角度都大于 45°,核实该研究区受 P元素限制[42-43]。值得注意的是,本研究酶C∶N和酶C∶P的平均值分别为1.251和0.505,低于全球尺度水平1.41和0.62,而酶N∶P的平均值为0.459,略高于全球水平0.44[14],说明土壤氮、磷代谢酶活性均高于全球水平,反映研究区主要受磷元素的制约,而氮元素也不富裕。刘立斌等对贵州高原型喀斯特次生林 C、N、P 生态化学计量特征的研究结果与本研究一致[10]。研究区土壤呈酸性,磷元素易被络合成不易被植物和微生物利用的形态,从而导致土壤磷元素受限[4]。此外,研究区属典型的喀斯特岩溶地貌,土壤空隙大,集中的雨季及其所引起的林溶会进一步降低P的有效性[41,44]。
3.3 土地利用方式转变过程中土壤酶活性变化的驱动因素
本研究RDA分析去共线性后留下了土壤全氮、速效磷、C∶N、N∶P和pH共5个因素,但经蒙特卡罗检验后,只筛选出土壤全氮、C∶N、N∶P和pH共4个显著影响土壤酶活性及计量比的因素,说明在土地利用变化过程中,这4个因素对土壤酶活性及其化学计量比的变化起主要驱动作用。速效磷被剔除的原因是土壤P素矿化程度低且速效磷对土地利用方式的转变相对不敏感,相比之下,不同土地利用方式间土壤全氮、C∶N、N∶P和pH差异明显,这些指标又影响着土壤微生物的生长发育,因而影响土壤酶活性的消长[45]。RDA结果显示,氮磷相关代谢酶与土壤全氮、N∶P和pH关系密切,并且与全氮含量正相关,与N∶P和pH负相关,碳代谢酶(BG、CBH、BX)和酶化学计量与土壤中C∶N正相关。从本质上来讲,该研究结果表明土壤酶活性与土壤养分具有趋同性,并受土壤pH的调控。土壤氮素含量的增加可通过提高植物生物量等方式增加土壤碳的输入,土壤微生物获取养分和能量后,根据资源分配理论,调节酶的分泌,改变酶的生化特性,进而提高酶的活性[41]。Sinsabaugh 等研究发现土壤NAG和AP活性与土壤pH负相关,这与本研究一致[14]。主要原因是大部分土壤水解酶的最适pH在5左右,土壤pH的升高或降低都会对酶活性产生不利影响[46]。本研究坡耕地变更土地利用方式后pH呈降低趋势,而土壤酶活性呈增加趋势,因此二者呈明显负相关关系。
对研究区7种土地利用方式的土壤养分含量和胞外酶活性进行聚类分析,可将其大致分为4类:灌木林地、梯田撂荒地和坡面摞荒地为一类,马尾松林地和柏树林地为一类,梯田菜地和坡耕地各为一类。由于每一类的土壤性状相似,因此,同一类的不同土地利用方式可互相替代,也可对同一类的不同土地利用方式采取相同管理措施。该研究结果具有一定实践指导意义,在进行植被恢复或土地管理过程中,可根据实际情况对同一类土地利用方式进行灵活调整,有助于减轻基层工作者的工作量并提高植被恢复或土地管理的成功率。
4 结论
土壤碳氮磷化学计量和酶活性化学计量特征显示研究区主要存在缺磷现象,但氮素也不富裕。对坡耕地转变土地利用方式后,土壤养分状况得到明显改善,不仅显著提高了养分含量,而且土壤微生物通过调节胞外酶的分泌和活性提高了磷素的有效性,促进了土壤养分的平衡。总体来看,坡耕地转马尾松林地和柏树林地后,土壤质量改善的效果相似,且较其它土地利用方式效果更加显著。值得注意的是,坡耕地转马尾松林地、柏树林地和梯田撂荒地后加剧了土壤磷素的限制,因此在造林过程中需要注意磷肥的补充。此外,土壤全氮含量、C∶N、N∶P和pH是土壤酶活性变化的主要驱动因素,土壤酶活性与养分含量的变化具有趋同性并受土壤pH的调控。
