5 种北美栎树秋冬季叶色变化机理分析
2021-07-08代泽娅孙海菁王树凤胡冀珍
代泽娅 谭 准 孙海菁 施 翔 王树凤 胡冀珍
(1.西南林业大学地理与生态旅游学院,云南 昆明 650233;2.中国林业科学研究院亚热带林业研究所,浙江 杭州 311400)
栎树(Quercusspp.)在世界上分布十分广泛,是北半球森林群落的重要建群种。栎树多为落叶或常绿乔木,少数为灌木,根深叶茂,有的秋季叶色五彩缤纷,常用于水源保护,遮荫和美化环境等,因此,栎树具有重要的生态服务功能和环境保护价值。中国本土栎树有60 余种,多用于山区造林。然而乡土栎树中的色叶品种并不多,秋冬季叶片呈色不理想,景观价值并不高[1]。2001 年,中国林业科学研究院亚热带林业研究所从美国东南部地区引进了20 多种栎树,其中很多树种秋季呈色明显,五彩缤纷,如纳塔栎(Quercus nuttallii)、舒玛栎(Q.shumardii)等,具有极高的观赏价值;并且具有速生、耐水湿能力,是长三角地区重要的彩叶景观树种[2]。通过多年的观测发现,引种的北美栎树秋季叶色变化主要有3 种类型:叶片呈现红色、叶片呈现黄色或褐色和叶片不变色或基本保持绿色;而在不同立地条件和气候条件下,即使是同一种栎树,其叶片呈色也会发生很大变化。目前有关引种栎树叶片呈色机理的研究报道较少,姜琳等[1]研究了栎属4 个树种秋冬转色期叶片的生理生化变化,吕秀立等研究了纳塔栎和舒玛栎不同年龄单株叶片转色期花青素、叶绿素以及可溶性糖等生理指标的变化[3],但由于近年来引种的北美栎树品种较多,除纳塔栎、舒玛栎以外,引种的其他栎树如水栎(Q.nigra)、柳叶栎(Q.phellos)、沼生栎(Q.palustris)等,在秋冬季也会出现明显的叶色变化,而因此,迫切需要对这些引种栎树的叶色变化及其影响因素进行评价和研究。
目前,国内外对彩叶植物叶片呈色机制的研究集中在色素的形成机制方面,已有大量研究表明,花青素的积累是彩叶植物叶片显色的主要原因,叶绿素、类胡萝卜素等作为色素的重要组成部分,对植物叶片的显色也起一定作用[4-5]。可溶性糖是花青素形成的前体物质,其含量与花青素的合成密切相关[6-7]。国内在三角枫(Acer buer-gerianum)、元宝枫(Acer truncatum)[8]、红花檵木(Loropetalum chinense var.rubrum)[9]、榉树(Zelkova serrata)[10]、檫木(Sassafras tsumu)[11]、乌桕(Sapium sebiferum)[12]等彩叶树种叶色变化的生理机制方面都做了一些研究。尽管目前国内已有研究关注引种北美栎树叶片呈色的生理机制,但大多数研究仅限于对叶片转色期间生理指标变化的讨论,针对影响北美栎树叶片呈色的环境因子和内在机制的研究还很少[13],根据引种栎树苗期叶色变化的观测,引种栎树中,秋季变色最早的是纳塔栎,一般在霜降之后开始变红,舒玛栎转色较迟,叶片呈色也为红色。不少水栎苗木叶片在秋冬季节变为红色[2]。姜琳等研究也发现,舒玛栎属于秋季变色较迟的栎树,但叶片红色最深[1]。本研究选取5 种秋季呈色不同的北美栎树,分别为黄色叶类型的柳叶栎,红色叶类型的纳塔栎、舒玛栎、沼生栎、水栎,且具有速生、耐水湿能力、叶片颜色鲜艳等特点,通过研究5 种北美栎树在秋冬转色期叶色变化以及叶片中色素含量、可溶性糖含量等生理指标的动态变化,分析其与环境因子的相关性,旨在揭示不同引种栎树在秋冬转色期叶片呈色的生理基础及其对环境因子的响应,以期为不同栎树在不同立地条件科学选择栽植以及栎树色叶品种的选育提供参考。
1 材料与方法
1.1 试验材料
5 种美国引进的栎树:水栎、纳塔栎、舒玛栎、柳叶栎和沼生栎,采样地为浙江省杭州市富阳区树种对比试验林,均为4 年生苗木,选取生长一致、健康植株树冠中上部叶片进行测定。
1.2 试验方法
1.2.1 取样
于2015 年10 月12 日至12 月16 日对5 种北美栎树取样,每个处理设3 次重复,单株为1 个重复,7~10 d 采样1 次,取每个重复每株树冠中部向阳面叶片3~7 片,每个处理一共取10~20 片叶片。同时在试验期间,在样地放置温湿度记录仪(DL−WS211,杭州尽享科技公司),每30 min 记录温湿度数据。
1.2.2 指标测定
叶绿素、类胡萝卜素采用丙酮浸提法[14],花青素采用盐酸甲醇浸提法[15],可溶性糖采用蒽酮比色法[16]。
1.2.3 数据分析
应用Microsoft Excel、Origin 7.5 等软件进行数据分析和图表处理,DPS 13.0 统计软件对数据进行单因素方差分析及叶片色素含量与温度及可溶性糖之间的相关性分析。
2 结果与分析
2.1 5 种栎树叶片转色期色素含量动态变化
2.1.1 花青素含量变化
5 种栎树叶片转色期花青素含量动态变化情况见图1。

图1 5 种栎树叶片转色期花青素含量动态变化Fig.1 Dynamic changes of anthocyanin content in leaves of 5 oak species during coloring period
从图1 可以看出,从10 月12 日开始,随着气温的下降,5 种栎树花青素含量逐渐上升,其中纳塔栎、水栎叶片变色较早,从11 月中旬开始叶片有部分变红,并在12 月7 日花青素含量出现峰值,此时气温降至5 ℃以下;但此后花青素含量便下降,原因可能是经历低温以后,叶片开始枯萎、脱落。其他3 种栎树花青素含量在12 月16 日出现最大值,而此时气温降至10 ℃以下。根据观察,在整个试验期间,柳叶栎叶色基本未出现颜色变化,其花青素含量在5 种栎树中最低。从11 月底开始,大部分栎树叶片开始呈现不同程度的颜色变化,花青素含量出现大幅增加,在变色期间,5 种北美栎树花青素含量最大值从大到小依次为:纳塔栎>水栎>舒玛栎>沼生栎>柳叶栎。
2.1.2 叶绿素含量变化
由 图2 可以看出,从10 月12 日 至12 月16 日,5 种北美栎树叶绿素a、叶绿素b 和总叶绿素含量均呈下降趋势。但在试验不同时期,2 种色素的变化规律表现不同:试验前期(10 月12 日到11 月5 日),大部分树种叶绿素a 下降趋势较为平缓(图2a),而叶绿素b 则出现了较为明显的下降(图2b),其中纳塔栎、舒玛栎和柳叶栎降幅最大。试验后期(从11 月11 日开始),5 种栎树叶绿素a 含量出现较为明显的下降,此后一直呈下降趋势(图2a),截至12 月16 日,5 种栎树叶绿素a 含量从大到小依次为:沼生栎>水栎>舒玛栎>纳塔栎>柳叶栎;而叶绿素b 在试验后期呈现较为平缓的下降趋势(图2b)。根据对温湿度的记录的发现,虽然温度的下降在一定程度上促进了栎树叶绿素含量的下降,但2 种叶绿素(叶绿素a 和叶绿素b)对温度的响应不同,在11 月5 日之前,气温依然在15 ℃以上,大部分树种叶绿素a 含量变化较为平缓,而叶绿素b 含量却出现大幅下降(图2a,2b),推测叶绿素b 含量的变化可能更多的依赖于树种自身的代谢过程。
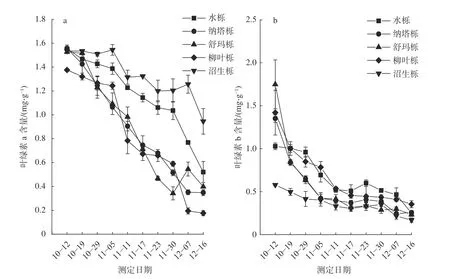
图2 5 种栎树叶片转色期叶绿素a 和叶绿素b 含量动态变化Fig.2 Dynamic changes of chlorophyll a,chlorophyll b content in leaves of 5 oak species during coloring period
2.1.3 类胡萝卜素含量变化
由图3 可知,同一时期内测定的类胡萝卜素含量在不同栎树之间存在明显差异,初秋季节(10 月12 日),舒玛栎叶片类胡萝卜素明显高于其他栎树,说明随着秋季气温下降,叶绿素合成的减少,舒玛栎较早的合成类胡萝卜素,以弥补叶绿素含量下降导致的光合速率下降。随着气温的进一步下降,到12 月份,柳叶栎较其他栎树叶片类胡萝卜素含量出现明显的增加,而根据观察,此时柳叶栎叶片明显开始变黄。在整个测定期内,大部分供试栎树叶片类胡萝卜素含量整体呈波动趋势,而柳叶栎类胡萝卜素含量在整个测定期呈波动上升趋势。

图3 5 种栎树叶片转色期类胡萝卜素含量动态变化Fig.3 Dynamic changes of carotenoids content in leaves of 5 oak species during coloring period
2.2 转色期叶片可溶性糖含量变化
由图4 可知,5 种栎树可溶性糖含量在转色期因树种的不同,变化趋势也有所不同。在整个测定期间,5 种栎树叶片可溶性糖含量呈波动变化:其中水栎和舒玛栎可溶性糖含量在整个转色期间变化不大,纳塔栎呈下降—上升—下降趋势,并在11 月30 日达到峰值;柳叶栎呈下降—上升—下降趋势,并分别在11 月17 日和12 月7 日出现峰值。而在不同的测定时间,各树种之间的可溶性糖含量差异并不明显,但在11 月底,纳塔栎可溶性糖含量明显高于其他栎树。
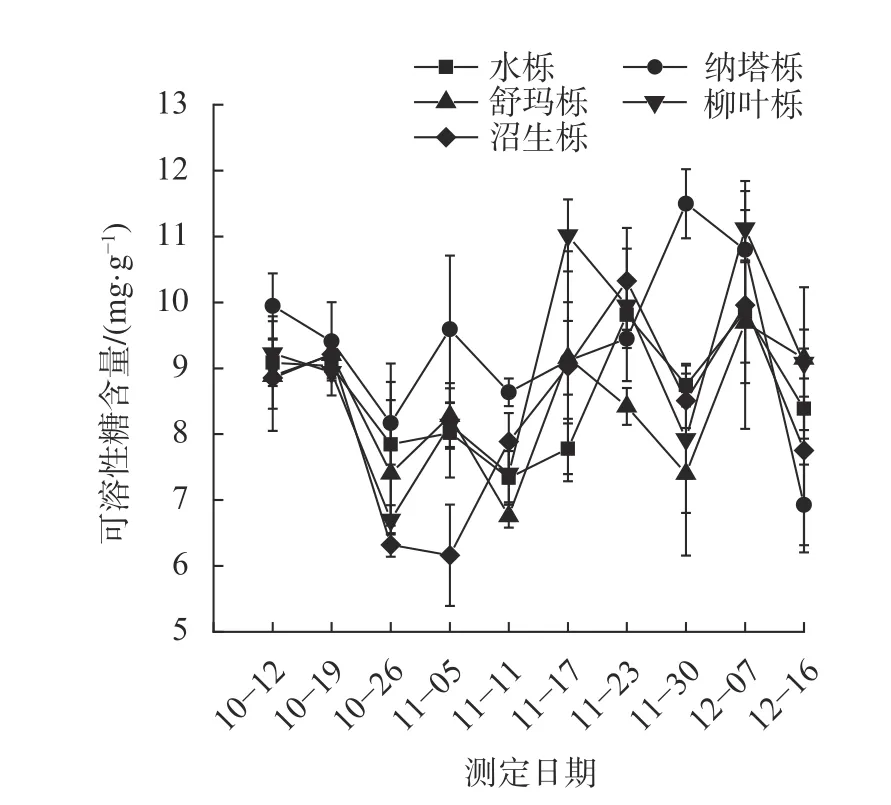
图4 5 种栎树叶片转色期可溶性糖含量动态变化Fig.4 Dynamic changes of soluble sugar content in leaves of 5 oak species during coloring period
2.3 叶片色素与可溶性糖和环境因子的相关性分析
通过对叶片叶绿素、类胡萝卜素和花青素含量与温度、平均湿度和可溶性糖含量的相关性分析(表1)可发现,供试5 种栎树中,只有水栎和柳叶栎叶片花青素含量与可溶性糖含量之间呈显著相关关系(P<0.05),其余树种花青素与可溶性糖含量的相关性并不显著,说明栎树花青素的合成对细胞内可溶性糖的依赖性存在物种间差异。而5 种栎树叶绿素和类胡萝卜素与可溶性糖之间的相关性均不显著,说明可溶性糖对这2 种色素的合成影响极小。
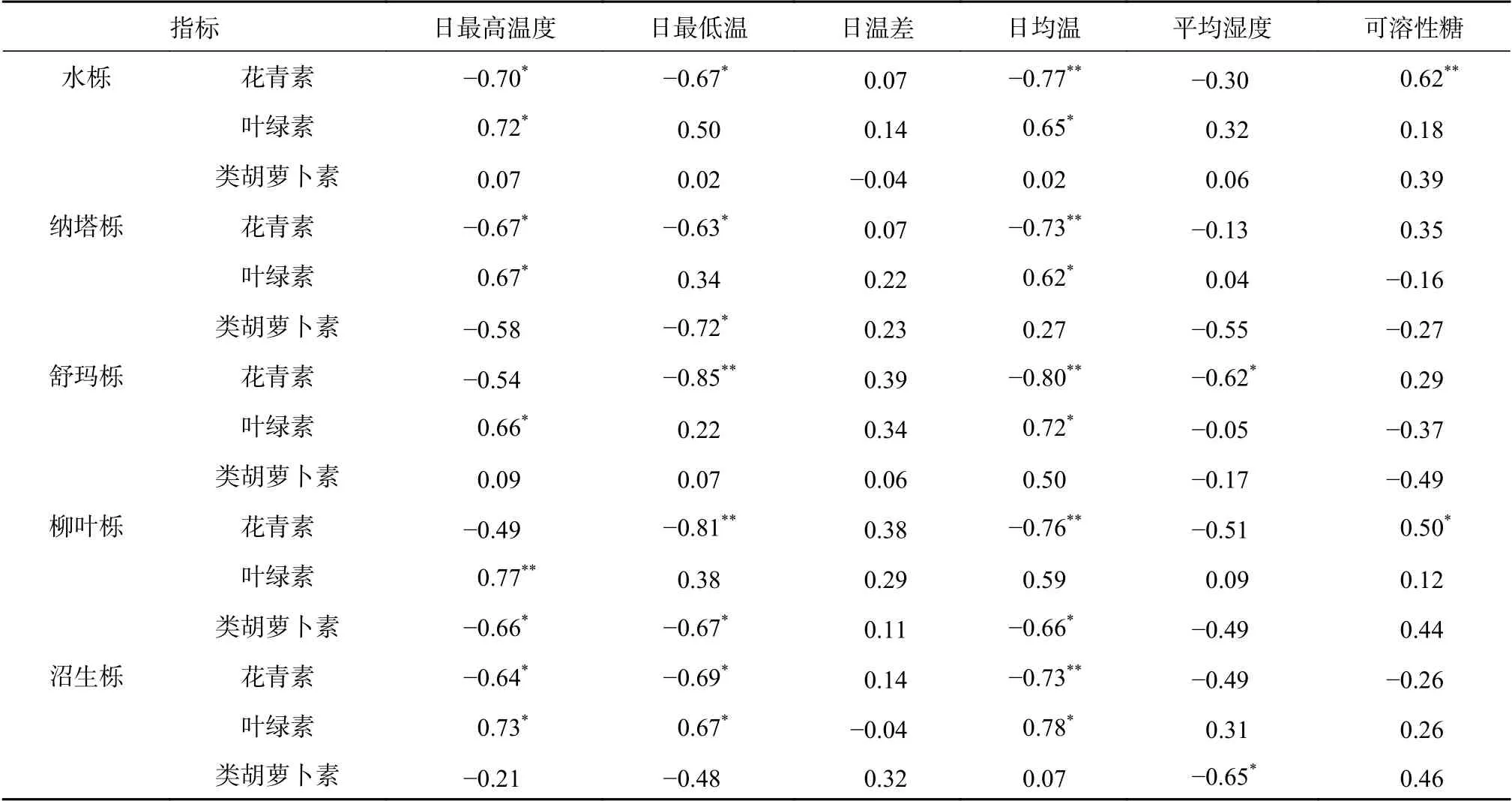
表1 叶片色素含量与温度和可溶性糖之间的相关性Table 1 The relevance of leaf pigment content between temperature and soluble sugar
秋冬叶片转色期环境因子变化最明显的是气温的下降,本研究对试验期间日平均气温,日最高、最低气温以及日温差与叶片色素含量相关性分析发现,5 种栎树叶片花青素含量与日平均气温呈极显著负相关(P<0.01),与日最高气温和最低气温也呈显著的负相关(P<0.05),而与日温差的相关性不明显,说明秋冬季气温变化是影响花青素合成的重要环境因子,低温促进花青素的积累,高温促进花青素的分解。大部分供试栎树叶绿素含量与日最高气温和日平均温度呈显著正相关(P<0.05),说明秋冬季气温下降,是导致叶绿素含量下降的关键因子。由表1 还可以看出,5 种栎树类胡萝卜素含量与温度的相关性不显著,推测叶片类胡萝卜素的积累可能有其自身的调节机制,不受温度变化的影响。同时发现,平均湿度对5 种栎树叶片色素含量的影响也不显著(表1)。
3 结论与讨论
2015 年10 月12 日至12 月16 日,整个试验期间,纳塔栎、沼生栎最先变为红色,舒玛栎变色时间稍迟,叶片也呈红色,柳叶栎在转色期以黄色为主,未出现明显的红色;水栎在转色期后期叶片变黄凋落,通过观测发现纳塔栎、舒玛栎2 种栎树叶片红色效果最为明显,与陈益泰等苗期的研究结果基本一致[2]。因此,从观赏角度而言,纳塔栎和舒玛栎属于秋叶赏红类型,根据多年观察,霜后柳叶栎叶片开始变黄,可以持续较长一段时间的色叶期,因此也具有较高的观赏价值。
叶片显色是不同因子综合作用的结果,植物叶片中色素的种类及含量决定了叶色的表达[4,17]。叶片中的色素主要有3 类:叶绿素、类胡萝卜素和花青素,其中叶绿素使叶片呈现绿色,类胡萝卜素在植物光合作用中起辅助作用,可以吸收光能并传递给叶绿素a,同时也是使叶片呈现黄色、橙色的主要色素[17]。花青素是植物体内主要的呈色物质,在酸性条件下呈现红色,在碱性条件下呈现蓝色。研究证实,花青素含量的多少直接体现出植物所呈现的色彩鲜艳程度[18]。植物叶片色素含量的变化是环境因素和内部代谢过程共同作用的结果[19-20],秋冬季彩叶植物叶片的转色主要是由于花青素和类胡萝卜素含量的增加,使叶片呈现由黄到红不同程度的彩色。一般认为,随着秋冬季日照时间的缩短和温度的下降,植物叶片开始自然衰老,从而启动叶绿素分解,而这种不利的环境,反而促进了花青素及类胡萝卜素的合成和积累,促使部分树木的叶色在秋季变成了红色或黄色[6,21-22]。根据试验研究表明,随着秋冬季气温下降,5 种北美栎树中,纳塔栎、舒玛栎、水栎、沼生栎叶片内花青素含量均有明显增加,而柳叶栎叶片内花青素含量增加幅度较小,类胡萝卜素含量增加幅度较大;秋冬转色期,纳塔栎、舒玛栎呈色效果优异,叶色鲜红;沼生栎叶片暗红;水栎叶片在转色后期色叶斑驳,然后枯黄凋落,色叶时间较短;柳叶栎叶片主要呈黄色,这与试验期观测结果一致。
彩叶树种秋冬季转色期间,温度是影响花青素积累的一个重要因素,研究表明[17,23],温度不仅影响花青素合成相关的酶活性,而且直接影响花青素的稳定性,低温会促进花青素的合成,高温则使植物分解代谢加剧从而导致花青素合成减少和分解[24]。研究表明,低温促进了日本红枫(Acer palmatum)叶片中花青素的积累,使叶片红色指数增加[7]。张敏等研究也表明,低温对榉树叶片花色素苷的生成有较强的促进作用[10]。本文研究也发现,随着气温的下降,5 种栎树花青素含量逐渐增加,而叶绿素含量逐渐降低。许晓波[13]研究了环境因子(包括平均温度、日照时数、最高气温、最低气温等)对秋季纳塔栎叶片色素积累的影响,结果发现,引起纳塔栎秋季叶片变色的主要因素是平均气温,其次是最高气温。本研究通过对5 种栎树叶片色素含量与平均温度的相关性分析也表明,5 种栎树花青素含量与平均温度呈极显著负相关,由此推测,秋冬季合适的低温是促进北美栎树叶片转色的重要环境因子。除了季节交替的原因以外,最近一些研究发现,其他很多环境因子包括紫外线、干旱、光胁迫等均能不同程度影响植物体内色素的合成和积累[25-28],而这些环境因子对栎树叶色变化的影响还有待于进一步研究。
除了环境因子以外,花青素的积累还与碳水化合物的代谢有关[7],糖是植物体内物质能量的来源和结构物质,罗兰[29]对彩叶草的研究表明,高温季节彩叶草叶片会发生褪色,主要原因是由于高温导致彩叶草体内碳水化合物储备不足,使花青素的糖链发生水解为植物体补充能量;而秋季夜晚温度较低时,暗呼吸的速率加速了糖的积累,从而促进了花色素的合成。可溶性糖是花青素形成的前体物质,有研究表明,三角枫、小叶鸡爪槭(Acer palmatum)等叶片内可溶性糖含量与花青素含量呈正相关,而在五角枫(Acer mono)和元宝枫叶片内相关性不明显[8]。吕秀立等研究发现,舒玛栎和纳塔栎叶片中花青素含量与可溶性糖含量呈极显著正相关[3];而姜琳等等研究却发现,舒玛栎和纳塔栎叶片内可溶性糖与花青素并无显著相关性[1]。本研究中水栎和柳叶栎叶片花青素含量与可溶性糖含量呈正相关,而舒玛栎、纳塔栎花青素含量与可溶性糖含量无显著正相关。由于可溶性糖对花青素合成的调控极为复杂,不仅是花青素的前体物质,而且也为其他色素如叶绿素、类胡萝卜素等提供底物和能源,同时也是色素形成过程中的信号物质[6,8],有研究推测,可溶性糖虽然影响北美栎树叶片花青素的合成,但也许并非决定性因素[1,8]。根据本研究结果推断,可溶性糖含量必然在一定程度上影响了北美栎树花青素的积累,但两者之间的相关性可能取决于测定的季节以及供试苗木的年龄等因素。然而,北美栎树作为一类引进树种,在国内的引种驯化不过十几年,相关的研究还很少,特别是针对其叶片呈色机理的研究,目前还处于探索阶段,因此,对于北美栎树叶片呈色的生理生化基础以及花青素合成的分子调控等研究工作,还需要开展进一步的探索。
