发酵剂对发酵香肠挥发性风味物质形成的作用及影响机制研究进展
2021-07-01刘英丽杨梓妍于青林曹雅婷蒋逸萱李洪岩
刘英丽,杨梓妍,万 真,于青林,曹雅婷,蒋逸萱,李洪岩,刘 洁,王 静
(北京工商大学食品与健康学院,北京食品营养与人类健康高精尖创新中心,中国-加拿大食品营养与健康联合实验室(北京),北京市食品添加剂工程技术研究中心,北京 100048)
发酵香肠是指将搅碎的猪肉(或牛、羊肉等)、动物脂肪、盐、糖、发酵剂和香辛料等混合后灌进肠衣,经过微生物发酵而成的香肠制品[1]。由于原料肉、香辛料的种类和比例的变化以及发酵和干燥条件不同,发酵香肠产品呈现出感官多样性,几乎每个国家都有自己的传统特色发酵香肠,如意大利的萨拉米香肠、德国的耐贮香肠、西班牙的肉干香肠、葡萄牙的阿萨多香肠、我国的腊肠等。发酵香肠品种多样,至今仍无正式且统一的分类标准和方法,常见按照脱水程度和发酵程度进行分类[2]。按照脱水程度进行分类可分为半干发酵香肠、干发酵香肠。半干发酵香肠又称快速发酵香肠,水分质量分数在40%以上。干发酵香肠水分质量分数小于40%,由于干发酵香肠的水分活度低,因此不用冷藏也可长久保存。按发酵程度(以pH值表示)可分为低酸发酵香肠(pH≥5.5)和高酸发酵香肠(pH<5.4)。发酵香肠的稳定性主要是由发酵和干燥过程中乳酸菌酸化和水分活度的降低综合作用决定的。在微生物发酵过程中主要的理化性质的变化为:pH值降低、脱水、硝酸盐及亚硝酸盐被还原、亚硝基肌红蛋白的形成、蛋白质的水解及胶凝化、脂肪水解和氧化现象[3]。
发酵香肠独特的风味与乳酸菌、革兰氏阳性过氧化氢酶阳性球菌(Gram-positive catalase-positive cocci,GCC+)及酵母菌等微生物的发酵作用密切相关。乳酸菌的主要功能是通过糖发酵产生乳酸降低香肠的pH值以保证其微生物安全性,同时使纤维蛋白胶凝化,从而提高最终产品的硬度和内聚性。GCC+有助于发酵香肠风味和色泽等典型特征的形成,其普遍具有较强的蛋白酶和酯酶活性,能促进肽、氨基酸、醛、游离脂肪酸的释放,从而有效促进发酵香肠特殊风味的形成。过氧化氢酶可以除去部分乳酸菌产生的H2O2,防止产品中形成高铁血红素和胆绿素,从而稳定产品的色泽并能够防止脂肪的过度氧化[4]。酵母菌主要生长在香肠表面或者肉馅靠外部分,可以保护香肠不受湿度波动的干扰,促进香肠的干燥过程并防止过度氧化[5]。此外,酵母菌能够促进蛋白水解和脂肪分解,从而影响香肠风味物质的形成。传统或自然干燥发酵香肠的生产不使用发酵剂,产品品质高度依赖自然环境,为了提高最终产品的品质和安全性,并使生产过程标准化,常以外源微生物作为发酵剂投入生产。肉类发酵剂是一类含有活性或休眠微生物的制剂,能在肉类中发挥理想的代谢活动[6]。大多数商业肉用发酵剂为乳酸菌和GCC+(葡萄球菌(Staphylococcus)和微球菌)的混合物。
风味是发酵香肠最重要的品质属性之一。与热加工产生的风味不同,发酵香肠的风味是由发酵和成熟过程中的化学反应和微生物代谢反应形成的[7]。发酵香肠的主要成分(蛋白质和脂类)经酶水解产生芳香化合物的前体物质——游离脂肪酸和氨基酸。这些前体物质作为化学反应和微生物反应的底物,产生挥发性风味物质[8]。近20 年来,许多研究报道了发酵菌株对于发酵香肠风味的贡献。Hugas等[9]指出接种GCC+菌株对于提升发酵香肠感官品质是十分必要的。此外,Stahnke[10]、Beck[11]和Olesen[12]等描述了木糖葡萄球菌(Staphylococcus xylosus)和肉葡萄球菌(Staphylococcus carnosus)通过氨基酸和游离脂肪酸转化为挥发性风味化合物来调节香气的能力。本文根据现有研究对发酵香肠发酵、成熟过程中挥发性风味物质的形成和发展以及与微生物代谢相关的反应进行概述,并总结了几种主要微生物对发酵香肠挥发性的风味物质的作用及影响机制,以期为发酵香肠风味的研究提供一定的参考。
1 发酵香肠挥发性风味的形成
1.1 挥发性风味化合物前体物质的生成
脂肪和蛋白质分解是发酵香肠风味形成的基本生化反应[13],为芳香化合物的产生提供前体物质(游离脂肪酸和氨基酸)[14-15]。脂肪分解是指存在于肌肉和皮下组织中的脂质部分(主要是甘油三酯和磷脂)的酶解。脂肪经原料肉中内源酶和微生物酶水解产生游离脂肪酸。与磷脂相比,甘油三酯水解产生更多的脂肪酸,酶解过程结束时检测到的大部分游离脂肪酸来自甘油三酯部分[16]。中链(碳数为6~12)、长链(碳数为14~18)游离脂肪酸的浓度通常低于其感知阈值,因此对发酵香肠风味的贡献很小,而短链脂肪酸(碳数小于6)会产生强烈的奶酪味或刺激性气味[17]。蛋白质水解包括内源酶和微生物蛋白酶(外蛋白酶和内蛋白酶)酶解肌浆和肌原纤维蛋白。由于酸性条件对微生物酶的抑制作用,仅有约40%的蛋白水解活性是由微生物酶产生[18]。内肽酶酶解作用下产生多肽,多肽通过外肽酶的活性进一步降解为多肽和游离氨基酸[19],这些多肽和游离氨基酸可产生苦味、甜味和鲜味。
1.2 挥发性风味化合物的生成
如图1所示,发酵香肠中产生挥发性化合物的反应可以分为化学反应和微生物代谢反应两类。化学反应包括脂肪酸自动氧化反应和美拉德反应等。微生物代谢反应可进一步分为碳水化合物发酵反应、氨基酸降解反应、β-氧化反应和葡萄球菌酯酶酶解[20]。每种反应对香味的贡献取决于发酵香肠实际的加工参数。
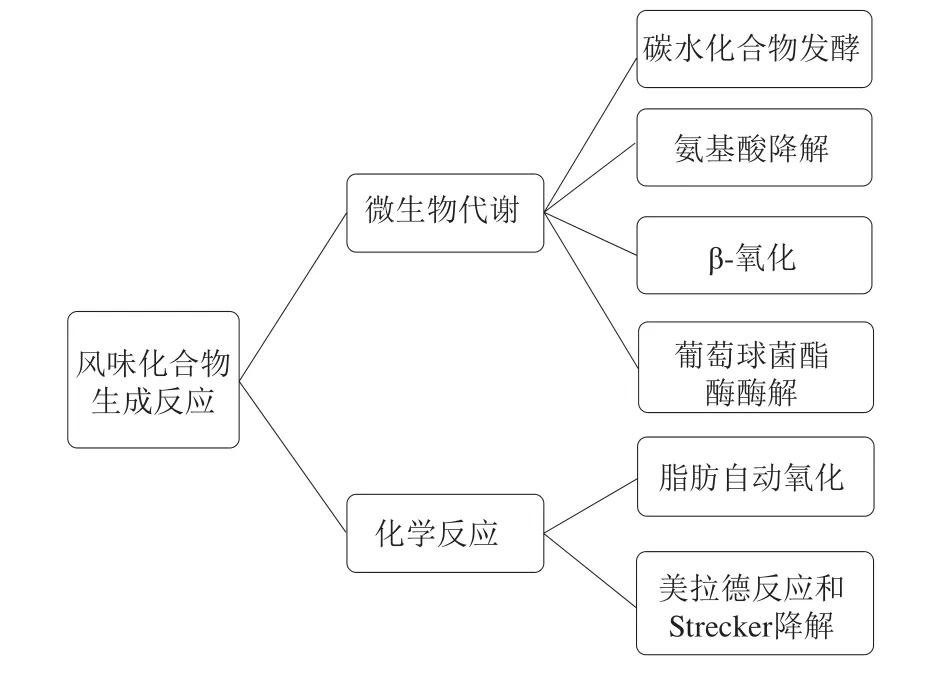
图1 发酵香肠中挥发性风味化合物的生成途径Fig.1 Chemical and microbial reactions involved in volatile compound generation in fermented sausages
1.2.1 产生挥发性风味化合物的化学反应
发酵香肠中最重要的化学反应是脂肪的自动氧化反应,特别是多不饱和脂肪酸的氧化反应[20]。脂肪的自动氧化与脂肪酸的不饱和程度密切相关,这个过程产生氢过氧化物,氢过氧化物进一步分解为挥发性化合物(烷烃、醛、醇、酯和羧酸)[21]。由于醇的气味阈值与其他羰基化合物相比较高,通常不被认为是发酵香肠风味的主要贡献者。很多醛是由不饱和脂肪酸氧化形成的,多于6 个碳原子的醛来自游离脂肪酸的氧化。由于醛的气味阈值低且在脂质氧化中生成率很快,因此是产生特有香气的成分之一[22]。虽然过度的脂肪自动氧化是发酵香肠品质劣变的主要原因之一,但适度的脂肪氧化对发酵香肠的风味形成也是必不可少的[20]。发酵香肠中的美拉德反应是由还原糖和氨基化合物反应引发的。在肌肉转化过程中,核糖核苷酸降解产生的核糖可作为参与美拉德反应的还原糖[23],肽和蛋白质水解产生的游离氨基酸作为参与美拉德反应的氨基化合物。美拉德反应开始时产生的挥发性化合物包括呋喃酮、糠醛、二羰基化合物等,进一步与胺、氨基酸、硫化氢、硫醇、氨等化合物反应产生不同的杂环化合物(吡啶、吡嗪、恶唑、噻吩、噻唑)[7],其特点是气味阈值低且有强烈的刺激性气味。Strecker降解反应[10]与美拉德反应有关,美拉德反应中碳水化合物的降解产生的二羰基化合物存在下氨基酸的氧化脱氨基和进一步脱羧,最后生成醛类化合物[24],与美拉德反应相比,氨基酸更易通过Stecker降解发生脱氨脱羧反应,这可能是许多羰基化合物,如分支醛2-甲基丁醛、3-甲基丁醛等的来源途径。
1.2.2 产生挥发性化合物的微生物代谢反应
发酵香肠中的碳水化合物发酵主要由乳酸菌完成。乳酸菌经代谢将发酵香肠中的碳水化合物(葡萄糖、蔗糖、乳糖和麦芽糖糊精)转化为乳酸和醋酸等有机酸,使发酵香肠pH值下降,肉蛋白发生胶凝化,从而提高香肠的硬度。乳酸菌通过丙酮酸代谢产生了乙酸、甲酸、乙醇、乙醛、2,3-丁二酮、3-羟基-2-丁酮和2,3-丁二醇等化合物[25],这些化合物与特定的风味有关。由氨基酸降解产生的挥发性风味物质对干香肠的风味特性起着重要作用:一个主要的氨基酸降解反应是支链氨基酸(缬氨酸、亮氨酸和异亮氨酸)的转胺和脱羧,产生各自的支化醛、醇和酸[20],如亮氨酸、异亮氨酸、缬氨酸、苯丙氨酸等分别生成3-甲基丁醛、2-甲基丁醛、2-甲基丙醛、苯甲醛及其醇、酸等衍生物,这些产物是发酵香肠典型风味成分的主要组成之一。酪氨酸和色氨酸的氨基酸侧链降解产生酚类和吲哚,其他氨基酸也可经微生物代谢转化为各自对应的醛类化合物。微生物对氨基酸的降解可以由发酵过程中存在的不同微生物种群来完成,葡萄球菌属和酵母菌均可降解氨基酸并产生具有重要香气贡献的挥发性化合物[18]。胱氨酸、半胱氨酸和甲硫氨酸的降解产生挥发性硫化物,甲硫氨酸产生甲硫醇、甲硫和氧化产物,如二甲基二硫和二甲基三硫。这些化合物的特征是气味阈值很低,具有类似熟肉、硫磺、卷心菜和洋葱气味。此外,微生物发酵过程中氨基酸分解代谢产生的酸和醇类化合物在葡萄球菌酯酶催化作用下生成酯类化合物[25]。游离脂肪酸通过微生物酶作用经β-氧化反应被降解,从而产生短链脂肪酸和β-酮酸,β-酮酸通过微生物(主要是葡萄球菌属)脱羧反应降解为甲基酮,然后进一步降解为仲醇[18,25]。仲醇通常呈青草、木材、脂肪、花香和蘑菇味,但其阈值高,因此对整体香气的贡献较小;而甲基酮的气味阈值低,具有果味和脂肪气味,对发酵香肠的典型风味的形成有重要影响。此外,葡萄球菌酯酶还能够水解酯类化合物生成醇和酸[24]。
1.3 挥发性风味化合物的提取与分离
对发酵香肠中挥发性化合物的定性、定量分析是揭示发酵香肠挥发性风味物质组成和含量的必要手段。提取技术的使用是必不可少的,但是实验分析得到的样品挥发性化合物组成高度依赖于所使用的提取技术,对同一样品使用不同的提取技术会产生不同的结果。表1共整理了6 种发酵香肠风味物质的提取技术。固相微萃取技术自20世纪90年代首次出现以来[26],在香气分析领域得到了广泛的应用。这种技术包括将挥发性化合物吸附到涂有不同选择性材料的二氧化硅纤维中,而后挥发性化合物由纤维被解吸到气相色谱仪的进样口。使用固相微萃取提取的香气成分组成取决于挥发性化合物在顶空与基质之间以及顶空与吸附纤维之间的分布系数。然而,纤维在平衡状态下吸附的化合物的数量与样品中总挥发性成分浓度直接相关,但只要知道每种挥发性成分的分布系数,就可以计算出其浓度。固相微萃取技术在发酵香肠风味提取中应用十分广泛,曹辰辰等[27]应用固相微萃取结合气相色谱-质谱(gas chromatography-mass spectrometry,GC-MS)技术提取并检测发酵香肠中的挥发性化合物,共分析出134 种挥发性化合物,其中包括醛类18 种、醇类24 种、酸类10 种、酯类25 种、酮类10 种、碳氢化合物41 种、其他6 种。
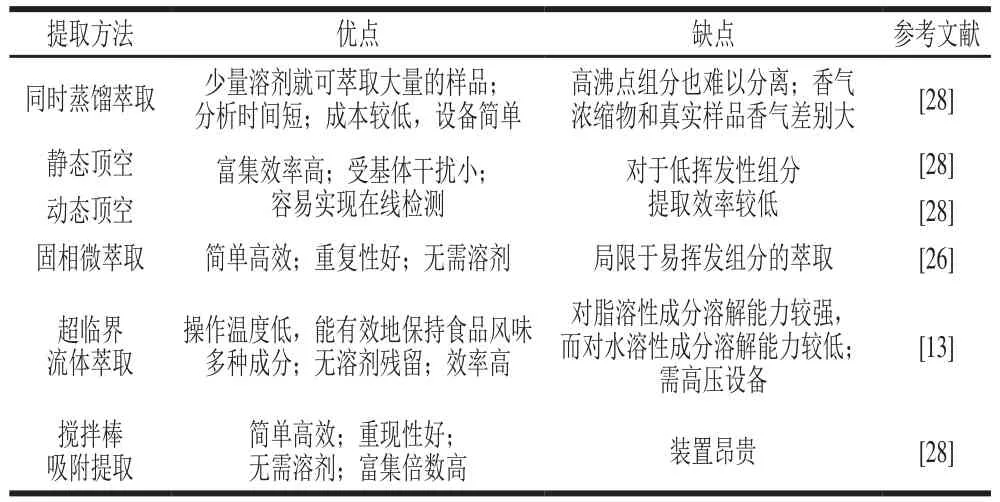
表1 发酵香肠挥发性风味物质提取技术Table 1 Extraction techniques used for analysis of flavor substances in fermented sausages
1.4 挥发性风味化合物的定性与定量分析
风味物质的分离,一般采用气相色谱柱法,可分离的组分多达百种,而对于单一组分的定性分析常采用质谱法、红外光谱、紫外光谱法、核磁共振波谱法。色谱分析也可与上述仪器联用,快速鉴定分离成分。目前常用的检测方法有GC-MS法、气相色谱与嗅觉检测联用(gas chromatography-olfactometry,GC-O)法和电子鼻检测法。GC-MS仪是一种将质谱和气相色谱组合使用的方法,它是目前最广泛使用的风味检测方法之一,其利用气相色谱分离混合物的组分,并且可以使用来自标准光谱的数据来比较质谱对香气化合物定性分析。Olivares等[29]采用顶空固相微萃取-GC-MS联用技术对干发酵香肠发酵、成熟过程产生的香气化合物进行了分析。近年来,风味研究的方向是阐明对食品风味贡献度较大的化合物,香气化合物贡献一般通过气味浓度与其气味阈值的比值计算气味活性值(odor activity value,OAV)得到。在含脂肪肉类中,高脂肪比例对气味阈值计算有很大影响。然而,OAV没有考虑挥发性化合物之间产生的协同效应。GC-O等方法可用于从复杂混合物中选择芳香活性成分,这是通过使用人的鼻子作为检测器的嗅探端口对从气相色谱仪洗脱的化合物进行筛选来完成的。该方法结合不同的分析技术(阈值稀释分析法、检测频率法、强度法)鉴定出食品中大量挥发性成分中真正具有气味活性的成分,以及各气味成分在不同浓度下对整体气味的贡献大小,这些都是用其他仪器检测难以做到的。在GC-O分析中,气相色谱柱的流出物由一名描述洗脱化合物气味的专家进行分析。根据嗅觉测定技术的不同,所得到的香气图将包括香气描述、强度、持续时间等。感官分析中人的鼻子一直是评价气味的重要手段,但是准确性及灵敏度却受到多种因素的影响,一方面此方法的主观性强、重复性差;另一方面,鼻子对气味具有适应性,容易出现疲劳而影响评判结果。随着计算机科学和仿生的发展,电子鼻弥补了这一不足。电子鼻是模拟人的嗅觉形成过程,由传感器阵列、信号预处理单元和模式识别系统3 个系统组成。与其他常规仪器分析方法相比,电子鼻分析具有样品无需前处理、不受人主观因素影响、响应时间短、检测速度快、重复性好等优点,能够很好地应用于在线检测。孙学颖等[30]利用固相微萃取-GC-MS联用结合电子鼻、电子舌技术对不同处理组羊肉发酵香肠样品的风味进行测定,4 组香肠样品中共鉴定出49 种挥发性风味物质,且结果表明基于电子鼻技术可实现对发酵香肠样品快速区分。对风味物质的定量分析,常用方法有归一法、内标法、外标法。内标法实验误差小,结果对比性强,将一定量的纯物质作内标物,再根据相应峰面积或峰高比求出待测组分的含量,是发酵香肠风味分析中常用的定量方法。
1.5 发酵香肠中的挥发性化合物
发酵香肠中挥发性化合物主要包括醛类、醇类、酯类、酮类、酸类、氮类和硫类化合物、呋喃类、萜烯类和内酯。醛类物质的阈值较低,主要来源于油酸和亚油酸等不饱和脂肪酸的氧化以及氨基酸降解,是发酵香肠中最重要的风味化合物[31]。其中己醛是发酵香肠中含量最高、最主要的醛类,被描述为青草味,其含量能够反映脂肪的氧化程度[32],戊醛、庚醛、辛醛和癸醛分别具有脂香、奶香、鲜草香和甜香味。酮类化合物有助于奶香味的产生,3-羟基-2-丁酮和2,3-丁二酮是与发酵香肠风味相关的酮类化合物,具有黄油味、蘑菇和香草等气味。醇类化合物通常对发酵香肠的香气贡献很小。Gianelli等[33]报道称这可能是由于醇类化合物具有较高的检测阈值。发酵香肠中最强烈的酸类风味化合物是乙酸、丁酸和3-甲基丁酸[31,34-36]。许多酯类化合物被认为是发酵香肠中水果味和焦糖味的来源,其中,丁酸乙酯、2-甲基丁酸乙酯和戊酸乙酯对发酵香肠的贡献较大[35]。硫化合物的特征是具有低阈值和特有的香味,这种特有的香味通常被描述为大蒜、洋葱、煮土豆、煮肉味。发酵香肠中气味最强烈的硫化物是二烯丙基硫化物、二烯丙基二硫化物、二甲基二硫化物、3-甲基硫代丙醇和甲硫醇[35-37]。3-甲基丁醛与含硫化合物反应,可产生类似培根的香气,且3-甲基丁醛的阈值很低,其浓度对发酵香肠的典型风味有重要影响,可作为量化菌株产香能力的指标。发酵香肠中的萜烯类化合物能具有植物芳香,如树脂、松树和柠檬味,其主要来源于香辛料,通常在含有辣椒等香辛料的发酵香肠中被检测出来。芳樟醇也被认为是一种气味较强的挥发性风味物质[38]。呋喃化合物有助于产生熟肉的香气,其中,2-戊基呋喃具有的香气最强烈,赋予香肠洋葱味、腐臭的气味。Olivares等[39]发现消费者偏好的高脂肪含量干发酵香肠香气与香味化合物己醛、2-壬烯醛、2,4-壬烯醛、丁酸乙酯和1-辛烯-3-醇有关,这几种化合物有助于青草味、脂肪味、水果味和蘑菇味的形成。表2整理了发酵香肠中的挥发性风味物质及其可能的形成机制。

表2 发酵香肠中检测出的挥发性化合物Table 2 Aroma volatile compounds detected in fermented sausages

续表2
以上通过对风味化合物前体物质的生成、风味化合物生成途径以及定性和定量分析,对发酵香肠风味研究进行了深入的探讨。然而,为了选择合适的原料、发酵剂和其他条件以提升发酵香肠的风味,还需要了解关于挥发性风味化合物的形成途径。
2 发酵菌株对发酵香肠挥发性风味的影响
2.1 乳酸菌对发酵香肠挥发性风味的影响
乳酸菌在香肠发酵过程中起主导作用。乳酸菌可以发酵碳水化合物产生乳酸、乙酸等,从而降低产品的pH值以提高产品的微生物安全性,同时可以使蛋白质发生凝胶化以加快香肠的干燥速度,终产物的切片性、硬度和内聚性都有所提高[46]。常用作肉品发酵剂的乳酸菌有清酒乳杆菌(Lactobacillus sakei)、弯曲乳杆菌(Lactobacillus curvatus)、植物乳杆菌(Lactiplantibacillus plantarum)等。乳酸菌对发酵香肠的安全性有重大意义,不仅可以通过产生有机酸降低肉品基质的pH值,也可产生细菌素抑制病原微生物的生长。Hebert等[47]公布了从手工发酵香肠中分离出的细菌素生产菌株L.curvatusCRL705的基因组序列(1.83 Mb),该菌株可产生双组分乳杆菌素705细菌素和AL705,AL705是一种具有抗菌活性的细菌素[48]。乳酸菌在发酵香肠中具有强的定植能力,占有绝对的数量优势。Sanz等[49]报道了一种存在于清酒乳杆菌中精氨酸特异性的氨基肽酶,该酶对游离氨基酸的释放起着重要作用,可以进一步进入精氨酸脱亚胺酶途径。L.sakei中精氨酸分解代谢所需的编码蛋白的基因聚集成簇,由精氨酸诱导,转录时受到葡萄糖抑制。Chaillou等[50]在L.sakei基因组中发现了第2种可能的精氨酸脱氨酶途径,该途径涉及两种精氨酸脱氨酶,可促进精氨酸的代谢,从而提高清酒乳杆菌在肉类环境中的竞争力。相较于GCC+,乳酸菌对发酵香肠风味的主要贡献似乎仅限于碳水化合物的分解代谢。近些年来,几种肽酶在乳酸菌中纯化和鉴定[49,51],这些内、外肽酶的作用有助于增加影响风味发展的游离氨基酸的浓度,使得乳酸菌潜在的产良好风味物质的能力逐渐引起关注。Sinz等[51]研究发现清酒乳杆菌TMW 1.1322、TMW 1.1383、TMW 1.1393具有不同程度代谢某些氨基酸、多肽的能力。Demeyer等[52]发现乳酸杆菌中的外肽酶与肌肉中的氨肽酶共同促进了游离氨基酸的产生,具有潜在的促进风味物质形成的能力。Hu Yingying等[53]研究了分离自哈尔滨风干肠的戊糖片球菌、短乳杆菌、弯曲乳杆菌及发酵乳杆菌在猪肌浆蛋白提取物中分解肌浆蛋白形成风味物质的潜力,结果发现这些乳酸菌能够分解肌浆蛋白形成风味物质及其前体,其中戊糖乳杆菌的分解能力最强,能够产生更多可溶肽和游离氨基酸。Papamanoli等[54]从希腊香肠中分离出的L.sakei、L.curvatus和L.plantarum均显示出高的亮氨酸和缬氨酸氨基肽酶活性。然而,对于乳酸菌利用风味前体物质(如支链氨基酸等)进一步分解代谢生成挥发性风味化合物的研究鲜有报道。Larrouture等[55]研究发现清酒乳杆菌、弯曲乳杆菌以及戊糖片球菌对亮氨酸的分解代谢率很低,对于3-甲基丁醇等典型香肠香气化合物的形成中几乎不发挥作用。Díaz等[56]称游离氨基酸含量的增加不足以显著增加香气化合物含量,因为还需要存在氨基酸降解机制以产生更多的风味化合物。综合而言,部分乳酸菌可利用肉类蛋白质产生更多的寡肽和游离氨基酸,为风味化合物的生成提供前体物质。Chen Qian等[57]以乳酸菌水解肌浆蛋白为例解释了风味化合物合成的可能途径,具体如图2所示。首先,肌浆蛋白被乳酸菌分泌的蛋白酶和肽酶降解为寡肽和氨基酸,一旦跨细胞膜转运,寡肽和氨基酸将被细胞内酶进一步降解。随后,氨基酸经氨基转移酶[58]或脱氨酶作用代谢成α-酮酸。作为中心中间体,α-酮酸可直接或间接脱羧形成相应的醛。直接方法是通过α-酮酸脱羧酶的非氧化脱羧,间接方法是α-酮酸脱氢酶和酰基激酶的氧化脱羧作用,导致形成羧酸,再于醛脱氢酶的作用下生成醛。醇脱氢酶可催化醛转化为相应的醇,醇再通过酰基转移酶或酯酶的作用与羧酸反应生成酯。在整个代谢过程中产生的挥发性的醛、醇、羧酸以及酯类化合物将影响发酵香肠整体的风味轮廓。表3整理了各类发酵香肠中不同细菌发酵剂对发酵香肠挥发性风味的主要影响。

图2 乳酸菌分解肌浆蛋白产生挥发性风味物质的可能代谢途径Fig.2 Possible metabolic pathways of lactic acid bacteria decomposing sarcoplasmic proteins to produce volatile flavor compounds
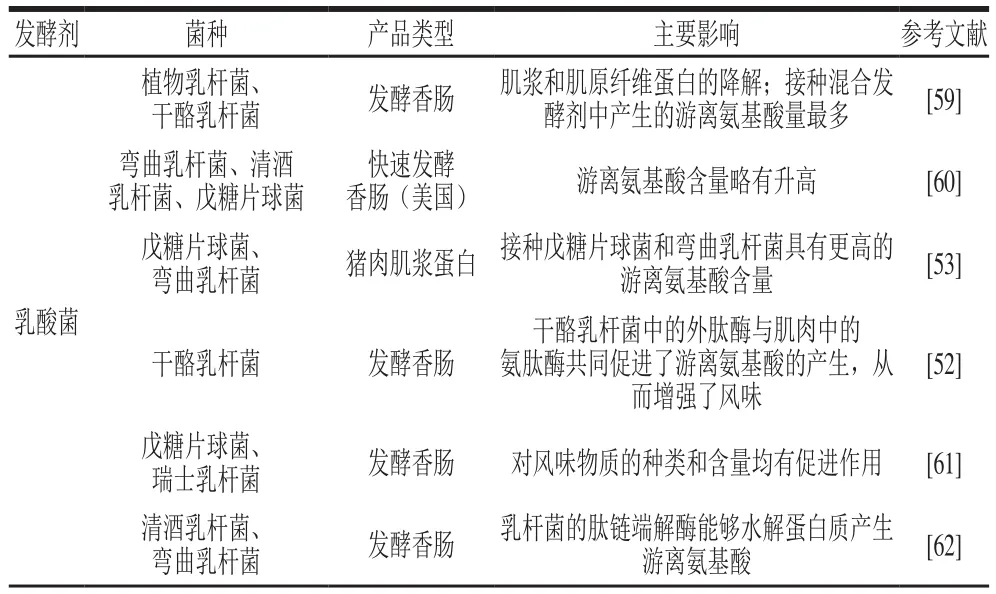
表3 细菌发酵剂对发酵香肠挥发性风味的影响Table 3 Effect of lactic acid bacteria on volatile flavor of fermented sausages

续表3
2.2 葡萄球菌对发酵香肠挥发性风味的影响
肉品发酵过程中的GCC+主要包括葡萄球菌和微球菌,其主要作用为提高色泽稳定性,缩短加工时间以及促进风味的形成。从发酵香肠中分离的GCG+主要是非致病性凝固酶阴性葡萄球菌(coagulase-negative staphylococci,CNS),如木糖葡萄球菌、马胃葡萄球菌(Staphylococcus equorum)、肉葡萄球菌以及腐生葡萄球菌(Staphylococcus saprophyticu)[72-74]。木糖葡萄球菌和肉葡萄球菌是常被用作发酵剂的GCC+,它们在肉类发酵过程中具有很强的竞争力,能够在高盐、低温等环境胁迫下生存。葡萄球菌主要通过其硝酸盐还原酶活性使发酵香肠呈现稳定的红色[75]。此外,葡萄球菌还有助于其他感官品质的形成,如质地和风味[76]是在蛋白质和脂类代谢相关酶的作用下形成的。之前的研究已经证明,发酵香肠的香味可由不同的葡萄球菌调节[18,77]。
Hugas等[9]强调了选用GCC+菌株作为肉用发酵剂以确保产品感官质量的必要性。其对风味的贡献主要源于以下几种反应类型:碳水化合物发酵、氨基酸代谢、脂质氧化及酯酶催化反应。碳水化合物经细菌发酵产生乳酸等有机酸,降低pH值从而使得肉蛋白凝固。这一过程中的主要角色由乳酸菌承担,而葡萄球菌也可缓慢地利用碳水化合物(图3),并将其转换为有机酸以及挥发性香气化合物,如2,3-丁二酮、乙醛、乙偶姻可以为发酵香肠带来黄油味、奶酪味等风味。
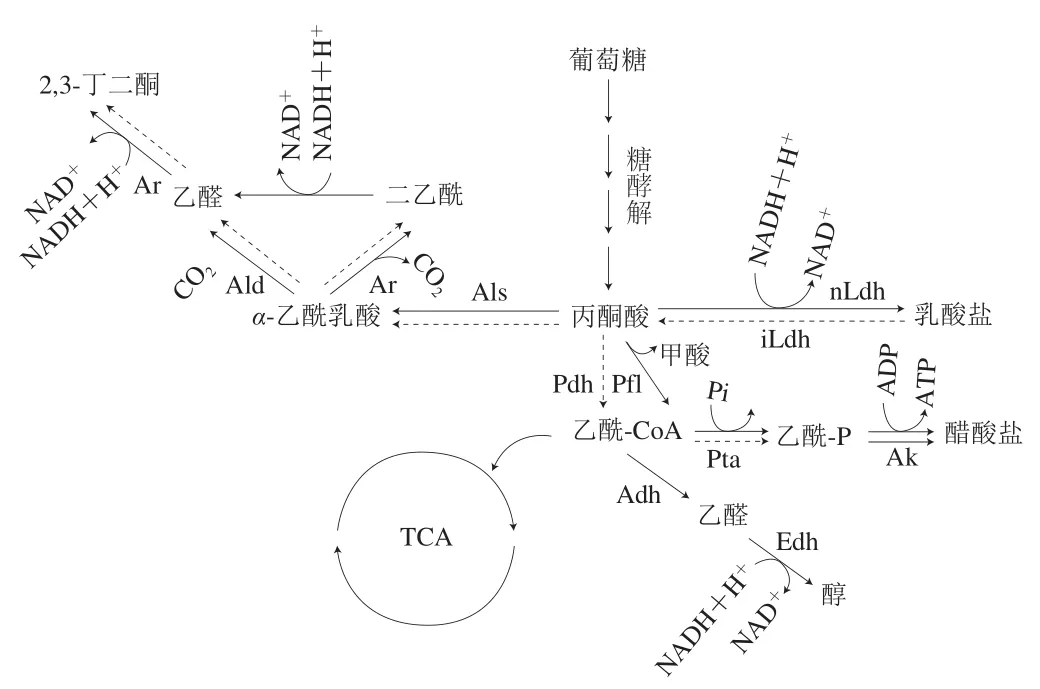
图3 葡萄球菌的葡萄糖代谢途径[78-80]Fig.3 Glucose metabolism in Staphylococcus[78-80]
Stahnke[10]、Beck[11]和Olesen[12]等描述了木糖葡萄球菌和肉葡萄球菌菌株通过将氨基酸和游离脂肪酸转化为挥发性风味化合物调节香气的能力。与碳水化合物代谢相比,氨基酸代谢在发酵香肠芳香化合物的产生中起着更为重要作用,其产物被认为与干发酵香肠的独特风味形成有关[62]。木糖葡萄球菌和肉葡萄球菌通过将氨基酸(特别是支链氨基酸:亮氨酸、异亮氨酸和缬氨酸)转化为挥发性风味化合物来调节香气,在此过程中支链氨基酸降解为甲基支化醛、醇和酸[11],图4是亮氨酸经微生物代谢向芳香化合物的转化途径,所产生的3-甲基丁醇、3-甲基丁醛和3-甲基丁酸被分别描述为果香、奶酪味、脚臭味,通常与发酵香肠的香气有关。Ravyts等[63,81]报道了3-甲基-1-丁醇对香肠风味复杂性的重要贡献,该化合物同样来源于葡萄球菌的亮氨酸分解代谢,如木糖葡萄球菌和肉葡萄球菌。此外,以S.carnosus833作为发酵剂的发酵香肠已经被证明可以产生更多的甲基酮、支链醛及醇类[10],可将意大利干发酵香肠的成熟时间缩短两周以上。
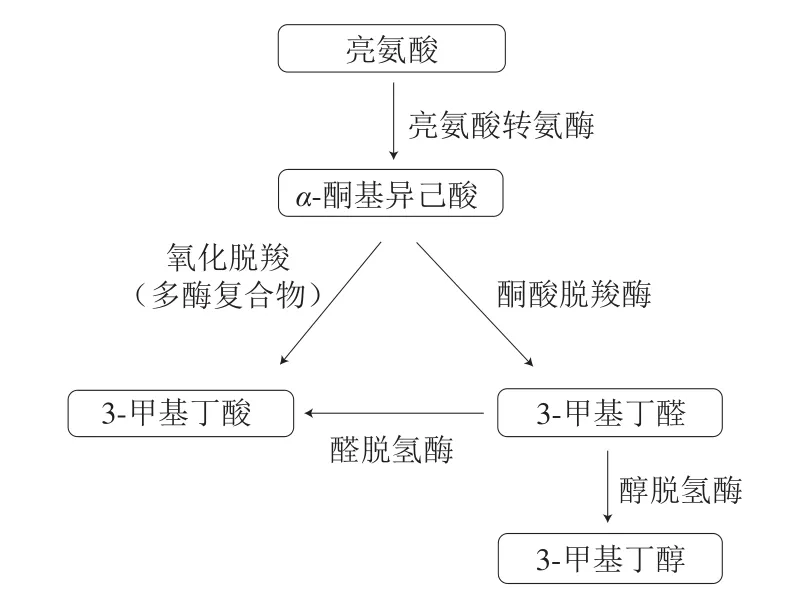
图4 亮氨酸转化为挥发性化合物的途径[62,82]Fig.4 Pathway for the conversion of leucine into aroma compounds[62,82]
发酵过程中脂肪分解主要由内源性脂肪酶负责[83],但葡萄球菌也被认为起到了重要作用。脂肪经酶解产生游离脂肪酸,而后游离脂肪酸通过不完全的β-氧化作用被葡萄球菌酶降解。游离脂肪酸首先被氧化为烯酰-辅酶A(coenzyme A,CoA),烯酰-CoA经水合作用生成羟酰基-CoA,再进一步氧化为酮酰-CoA。酮酰基-CoA酯通过硫酯酶作用被脱酰基为β-酮酸、短链游离脂肪酸和CoA[66]。β-酮酸由葡萄球菌通过氨基酸脱羧反应产生甲基酮,例如2-戊酮、2-己酮和2-庚酮,这有助于发酵香肠风味的形成[84]。甲基酮可以进一步转化为仲醇。Søndergaard等[77]报道添加木糖链球菌产生3-甲基-1-丁醇、二乙酰基、2-丁酮、乙酰丁酮、苯甲醛、苯乙酮和甲基酮(2-戊酮和2-庚酮)等重要风味化合物,可以为发酵香肠带来更“圆润”的风味,在南欧常被用于生产芳香型香肠。
2.3 酵母菌对发酵香肠挥发性风味的影响
在发酵香肠成熟期间,酵母是主要的微生物群之一。其中,假丝酵母属(Candida)、德巴利氏酵母属(Debaryomyces)和耶氏酵母属(Yarrowia)是最常见的属[85]。酵母主要定植在发酵香肠表面或者肉馅靠外部分,可以在香肠外壳表面形成一层膜,防止产品过度脱水,并有利于最终风味的形成。真菌的生长使香肠表面形成有于发酵成熟的微环境,减少香肠表面黏连和脂质氧化[86]。酵母菌具有较强的脂肪和蛋白质水解活性,具有赋予发酵香肠独特风味的潜力。其中,无论是单独接种还是作为混合发酵剂的一部分进行混合接种发酵,德巴利氏酵母属均可广泛参与肌原纤维蛋白的水解[87]。Santos等[88]研究了分离自猪肉中的汉逊德巴利酵母(Debaryomyces hansenii)水解肌浆蛋白的活性,结果表明汉逊德巴利酵母具有水解肌浆蛋白的活性,可以增加猪肉中极性和非极性肽以及游离氨基酸的生成量,其中Tyr、Val、Met、Leu、Ile、Phe和Lys含量增幅均超过50%。Patrignani等[89]报道,耶氏酵母属具有较强的脂质和蛋白水解活性,对于游离脂肪酸释放有积极作用,可进一步转化为理想的风味化合物。
Olesen等[90]研究了产朊假丝酵母(Candida utilis)对发酵香肠香味形成的影响,观察到其具有较高代谢活性,增加了香肠中挥发性化合物,特别是酯类化合物的含量;此外,C.utilis代谢支链氨基酸缬氨酸后产生的挥发性化合物(2-甲基丙酸和2-甲基丙醇等)对香肠香气形成有重要影响。支链氨基酸作为许多挥发性风味化合物的前体物质,其代谢产物是发酵香肠典型风味的主要组成之一。Durá等[91]发现汉逊德巴利酵母可以通过利用支链氨基酸生成挥发性化合物这一途径影响发酵香肠的香气轮廓,但食用盐含量会对其产生负面影响。Flores等[92]通过接种汉逊德巴利酵母和混合细菌发酵剂生产发酵香肠,他们观察到2-甲基-丙酸乙酯、2-甲基丁酸乙酯和3-甲基丁酸乙酯等化合物的含量显著增加,这3 种物质都与香肠的香味有关,此外发酵香肠中由脂质氧化生成的挥发性化合物(如己醛)含量显著减少,同时乙酯生成量显著上升。这一结果表明汉逊德巴利酵母能抑制脂质氧化,并促进酯类化合物的产生。Cano-García等[93]对酵母菌提升风味的潜力进行了深入探讨。他们的研究表明,酵母菌特别是汉逊德巴利酵母具有较好的提升风味潜力,在改善、提升香肠风味亦或使香肠风味多样化上具有较好的应用前景,且主要通过抑制脂质氧化、延缓酸败和产生醇类、酮类、酯类等风味物质而赋予香肠良好风味。表4整理了不同发酵香肠中酵母菌对发酵香肠风味形成的影响。
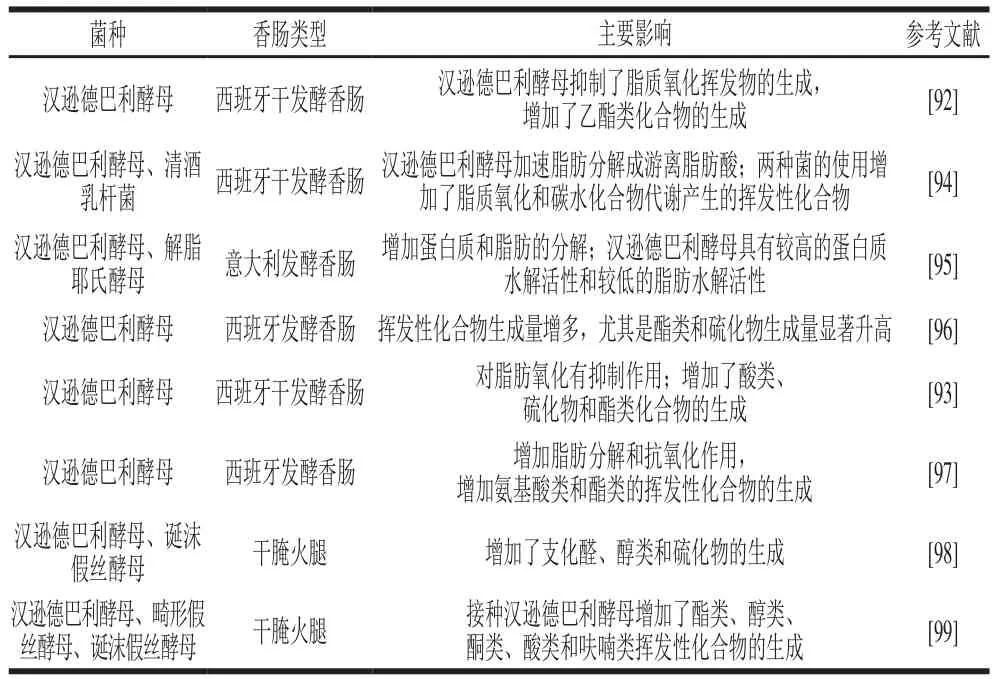
表4 酵母对发酵香肠挥发性风味形成的影响Table 4 Effect of yeast on volatile flavor of fermented sausages
3 基于高通量测序技术对挥发性特征风味与菌群相关性的分析
发酵香肠的发酵过程属于开放式多菌种混合发酵体系,微生物是发酵的主体,也是风味物质的主要生产者。长期以来,研究者主要采用基于纯培养方法对微生物进行分析,依靠形态学、培养特征、生理生化特性的比较对香肠发酵过程中微生物进行研究,然而这种方法只适用于可培养的微生物且效率低下。20世纪90年代初,以聚合酶链式反应为基础的分子生态学的发展带来了描述细菌群落的新方法,例如随机扩增多态性DNA-聚合酶链式反应、实时聚合酶链式反应、温度梯度凝胶电泳、变性梯度凝胶电泳)[100-101]。这些基于聚合酶链式反应的分子生态学方法比传统依赖于培养的方法对发酵香肠生物群落的认识更加全面。Fonseca[100]和Cocolin[102]等使用变性梯度凝胶电泳在发酵香肠中鉴定乳酸菌,包括乳杆菌清酒、弯曲乳杆菌、植物乳杆菌,以及肠球菌、片球菌和金黄色葡萄球菌几个物种。但是这些方法也存在其自身的局限性,方法本身需要进行凝胶电泳,图谱的复杂性造成了有限的分辨率和灵敏性且工作量较大费时费力;自2005年以来,以罗氏公司454测序技术、Illumina公司的Solexa测序技术和ABI公司的Solid测序技术为代表的高通量测序技术相继诞生,并迅速广泛应用于传统发酵食品的微生物群落的宏基因组研究中。迄今为止已有部分研究者使用高通量测序技术对发酵香肠的微生物多样性进行分析。Połka等[103]通过高通量测序揭示了典型意大利香肠中不同成熟阶段的细菌多样性。Wang Taojun等[104]通过高通量测序技术对意大利萨拉米香肠、中式腊肠等发酵香肠的细菌群落进行了分析,结果表明中式腊肠比意大利萨拉米香肠的细菌种类更为丰富。腊肠的主要细菌为葡萄球菌和乳酸菌,乳酸菌包括乳酸菌、乳杆菌、片球菌和乳球菌。葡萄球菌属的操作分类单元在意大利腊肠中最多,高达97.45%。
利用高通量测序技术能详细、系统地分析发酵香肠中微生物群落的结构以及群落功能,在此基础上多元统计学的应用,如偏最小二乘法回归分析、偏最小二乘判别分析和正交偏最小二乘法回归分析等,可进一步挖掘数据背后的生物学意义,建立起发酵香肠中微生物菌群与特征风味物质的相关性联系。米瑞芳等[105]对4 种传统自然发酵酸肉中细菌16S rDNA V3~V4区进行高通量测序,并结合电子鼻和固相微萃取-GC-MS分析等手段,检测出挥发性物质包括酸类、醇类、醛类、酯类、酮类、萜烯类等化合物共126 种,结果表明4 种酸肉样品之间的菌属种类和相对丰度均存在一定的差异性,同时酸肉样品中的风味物质在种类和含量上也存在较大差异性;且乳杆菌、魏斯氏菌等微生物可能影响酸肉中醇类、酯类和酸类挥发性风味物质,从而影响酸肉的风味品质。郭壮等[58]采用电子鼻对不同发酵时间腊肠风味品质进行评价,并采用MiSeq高通量测序技术对细菌的多样性进行解析,主坐标分析和肠型分析的结果显示不同发酵时间腊肠样品在空间排布上和基于腊肠风味指标的聚类结果相近。由此说明腊肠样品中的细菌影响腊肠风味品质的形成。此外,通过将优势细菌属相对丰度和风味指标进行关联分析发现,乳杆菌属相对丰度与电子鼻传感器W1S、W2S(对乙醇灵敏)和W3S(对烷烃灵敏)的响应强度呈显著正相关,葡萄球菌属相对丰度与W1S响应强度之间呈显著负相关,而魏斯氏菌属和环丝菌属相对丰度与电子鼻传感器W2W(对含硫化合物灵敏)响应强度之间存在显著负相关关系。Hu Yingying等[106]发现传统干发酵香肠中的主要挥发性化合物与核心细菌(乳酸乳球菌、植物乳杆菌、消化乳杆菌、清酒乳杆菌、魏斯氏菌属)之间存在显著相关性,这些核心菌群促进了主要挥发性化合物的形成。Hu Yingying等[107]采用高通量测序技术分析了东北地区5 种发酵香肠的细菌菌落组成和多样性,并采用顶空固相微萃取-GC-MS法检测出包括醇、酸、醛、酮、酯和萜烯类共120 种挥发性化合物。此外,基于Spearman相关分析,探索了核心细菌与主要挥发性化合物之间的潜在相关性,结果表明:干香肠中的主要挥发性化合物与核心细菌(乳酸乳球菌、植物乳杆菌、消化乳杆菌、清酒乳杆菌、希腊魏斯氏菌)之间存在显著相关性,这些核心菌群促进了主要挥发性化合物的形成。
4 结 语
随着我国改革开放的深入,我国的肉类工业得到了飞速发展,国民对于高档发酵肉制品需求量不断提升是国内发酵香肠发展的机遇。自20世纪80年代起,众多国内肉品企业开始引入西式发酵香肠,如切片萨拉米、即食发酵香肠等,以满足我国消费者对中高档发酵肉制品的需求。但我国消费者和欧美消费者对风味的需求存在较大差异使得发酵香肠未得到我国消费者的充分认可。因此,在引进西式发酵香肠先进技术的同时,开发“中国化”的风味是目前我国肉品企业面对的挑战。发酵香肠风味的形成与发展与发酵过程中微生物群落的代谢活动密切相关,在继续开展优良肉用发酵剂筛选工作的同时还应明确:主要微生物在整个发酵过程中的动态变化以及与风味物质变化之间是否有关联性;主要的风味物质是由哪些微生物产生,通过何种代谢途径产生。只有明确了上述核心问题,对微生物群落结构、微生物与风味物质的相关性进行深度分析与挖掘,搭建起菌群与风味物质的应答关系,才能有针对性地对发酵香肠的风味品质进行调控,保证发酵香肠的产量和质量。
