追施氮肥对沙地生境下不同饲用燕麦品种叶片氮代谢酶活性的影响
2021-07-01张玉霞张庆昕斯日古楞朱爱民
张玉霞,王 鑫,张庆昕,斯日古楞,郭 园,朱爱民
(1.内蒙古民族大学,内蒙古 通辽 028041;2.内蒙古农业大学,内蒙古 呼和浩特 010018)
氮代谢在植物的生命活动中起至关重要的作用,直接影响作物的发育[1]。在氮素同化过程中,硝酸还原酶(Nitrate reductase,NR)、谷氨酰胺合成酶(Glutamine synthetase,GS)、谷氨酸合成酶(Glutamate synthetase,GOGAT)、谷氨酸草酰乙酸转氨酶(Glutamate oxaloacetate transaminase,GOT)和谷氨酸丙酮酸转氨酶(Glutamate pyruvate transaminase,GPT)酶活性影响植物氮代谢过程[2]。研究表明,NR活性的高低和氮同化能力密切相关,GS活性等关键氮代谢指标均随施氮水平的增加而提高,NR、GS和GOGAT氮代谢关键酶活性越高,氮素同化的能力就越强。氮效率不同的作物品种之间在施氮量处理下氮代谢酶差异明显[3]。
燕麦(AvenasativaL.)是禾本科燕麦属一年生粮饲兼用作物,广泛分布于世界各地,种植面积在全球范围内位居第6位,普通栽培燕麦类型分为皮燕麦和裸燕麦,其中皮燕麦起源于伊朗和俄罗斯等地区,裸燕麦则起源于中国和蒙古[4]。皮燕麦具有耐寒、抗逆性强、适口性好、产量高、易于栽培等优点,是一种重要的一年生饲草[5]。近年来在内蒙古地区以喷灌为主的沙地苜蓿草产业发展迅猛,燕麦作为苜蓿倒茬轮作的首选牧草种类,种植规模也在不断扩大,在沙化草地草产业发展中具有重要地位[6-7]。增施氮肥是提高科尔沁沙地生境下饲用燕麦的产量和品质的关键栽培技术措施[8],目前,关于科尔沁沙地生境下增施氮肥开展了燕麦光合特性、氮肥利用率、衰老特性等方面的研究[9-11],而关于氮代谢酶与施氮量及品种之间差异研究较少[12]。研究不同饲用燕麦品种氮同化过程中相关酶活性的差异对于科尔沁沙地饲用燕麦的规模化种植具有重要意义。有研究表明,氮效率不同的作物品种之间氮代谢酶差异明显[13-14]。为此,在科尔沁沙地选择主栽饲用燕麦品种进行施氮量处理,检测不同部位叶片的氮代谢酶活性,分析氮代谢酶活性在施氮量和饲用燕麦品种之间的差异,为筛选氮高效饲用燕麦品种及合理施用氮肥提供参考依据。
1 材料和方法
1.1 试验地概况
试验地位于内蒙古自治区内蒙古民族大学科技园区(43°30′N,122°27E),属于温带大陆性气候。土壤为风沙土,土壤有机质含量7.28 g/kg,全氮含量1.87 g/kg,碱解氮含量11.24 mg/kg,速效钾含量95.12 mg/kg,速效磷含量10.59 mg/kg。年平均气温0~6 ℃,≥10 ℃积温3 000~3 200 ℃,无霜期140~150 d,年平均降水量340~400 mm,年平均风速3.0~4.5 m/s。
1.2 试验设计
试验采用燕麦品种和氮肥施用量二因素随机区组试验设计,共8个处理,4 次重复,小区面积4 m×5 m=20 m2,四周设保护行。供试饲用燕麦品种为牧王和甜燕1号,分别来源于北京正道生态科技有限公司和北京佰青源畜牧科技发展有限公司,原产地均为加拿大。于2019年4月1日采用条播方式种植,条播行距15 cm,播种量150 kg/hm2,播种深度3 cm,播种时施用磷肥(P2O5)和钾肥(K2O)均为150 kg/hm2,磷肥为重过磷酸钙(P2O546%)、钾肥为氯化钾(K2O 50%),在燕麦生长期追施0(CK),100,200,300 kg/hm2氮肥(纯氮),分别用N0、N100、N200、N300表示,肥料为尿素(含N 46%),按照15%,40%,25%,20%比例在分蘖期、拔节期、孕穗期、开花期4次追施,撒施后及时灌水,灌溉方式为喷灌。于灌浆期取燕麦的旗叶、倒二叶、倒三叶测定氮代谢酶活性。
1.3 测定指标及方法
硝酸还原酶(NR)测定采用活体磺胺比色法[15];谷氨酰胺合成酶(GS)酶活性参考赵世杰等[16]的方法测定;谷氨酸合成酶(GOGAT)参照赵权志等[17]的方法测定;谷氨酸草酰乙酸转氨酶(GOT)和谷氨酸丙酮酸转氨酶(GPT)参照马新明等[18]和王静等[19]的方法测定。
1.4 数据处理
采用Excel 2003和SPSS 26.0进行数据处理和方差分析。
2 结果与分析
2.1 追施氮肥对饲用燕麦GOGAT活性的影响
由表1可知,随着施氮量的增加饲用燕麦叶片的GOGAT活性呈先增加后降低的变化趋势,且均在N200处理下GOGAT活性最强,牧王旗叶、倒二叶、倒三叶的GOGAT活性分别为0.25,1.14,0.22 μmol/(g·min),甜燕1号旗叶、倒二叶、倒三叶的GOGAT活性分别为0.16,0.80,0.18 μmol/(g·min),显著高于N300和N0处理(P<0.05),牧王的旗叶、倒二叶、倒三叶GOGAT活性在N200和N100处理之间差异不显著(P>0.05),甜燕1号的倒二叶和倒三叶的GOGAT活性则是N200显著高于N100(P<0.05);由此说明,追施氮肥有利于提高GOGAT的活性,但过高亦会导致GOGAT活性下降。在不同氮肥处理下,除N300的旗叶和N100的倒三叶外,牧王的旗叶、倒二叶、倒三叶的GOGAT活性均显著高于甜燕1号饲用燕麦品种(P<0.05),说明在相同氮素水平下不同饲用燕麦品种氮素同化水平存在明显差异。由叶片不同部位得知,倒二叶的GOGAT活性最强,牧王在N0、N100、N200、N300处理下GOGAT活性分别为0.34,1.04,1.14,0.61 μmol/(g·min),甜燕1号在N0、N100、N200、N300处理下GOGAT活性分别为0.23,0.59,0.80,0.43 μmol/(g·min),其次是旗叶,倒三叶的GOGAT活性最弱,因此,倒二叶是评价氮代谢酶的最佳叶片。
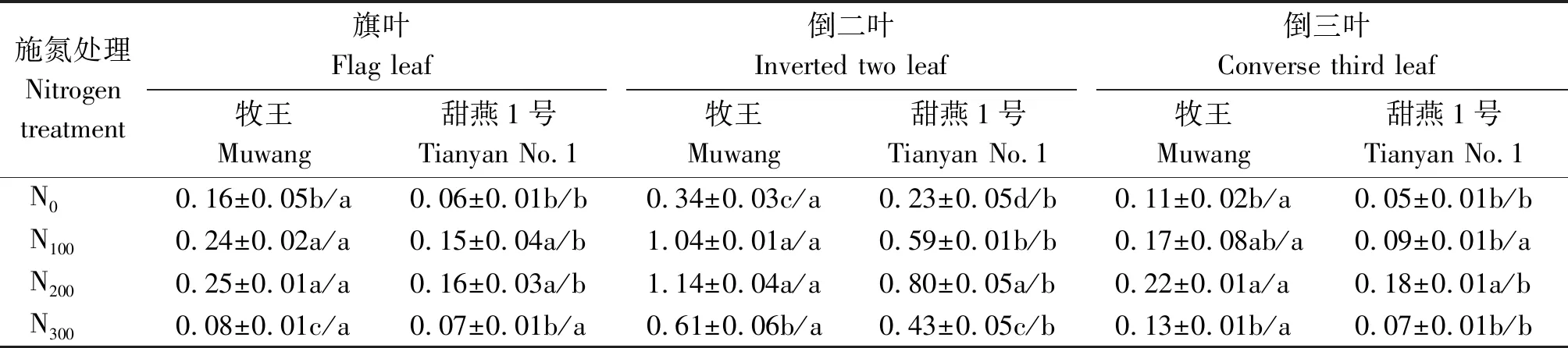
表1 不同施氮量下沙地饲用燕麦叶片GOGAT活性的变化Tab.1 Changes of GOGAT activity in leaves of forage oat in sandy land under different nitrogen application rates μmol/(g·min)
2.2 追施氮肥对饲用燕麦GS活性的影响
由表2可知,随着施氮量的增加,饲用燕麦叶片的GS活性呈先增加后降低的变化趋势,均在N200处理下GS活性最强,牧王旗叶、倒二叶、倒三叶的GS活性分别为10.68,14.10,11.83 μmol/(g·min),甜燕1号旗叶、倒二叶、倒三叶的GS活性分别为7.43,10.02,9.03 μmol/(g·min),其中牧王饲用燕麦品种N200处理叶片的GS活性显著高于N0、N100、N300处理(P<0.05),甜燕1号饲用燕麦品种的旗叶GS活性在不同处理之间差异不显著(P>0.05),倒二叶的GS活性则是N200处理显著高于其他氮肥处理(P<0.05),倒三叶的GS活性则是N200和N100显著高于N0(P<0.05),但与N300差异不显著(P>0.05)。2个饲用燕麦品种之间则是牧王显著高于甜燕1号(倒三叶的N0和N100处理除外)(P<0.05)。不同叶片之间则是倒二叶的GS活性较强。
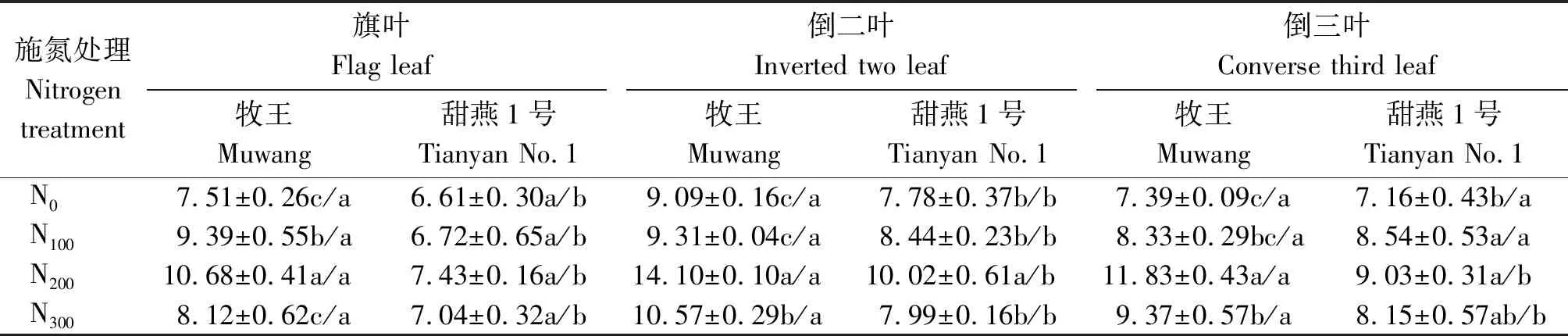
表2 不同施氮量下沙地饲用燕麦叶片GS活性的变化Tab.2 Changes of GS activity in leaves of forage oat in sandy land under different nitrogen application rates μmol/(g·min)
2.3 追施氮肥对饲用燕麦NR活性的影响
由表3可知,牧王饲用燕麦品种叶片的NR活性均显著高于甜燕1号(P<0.05),随着施氮量的增加饲用燕麦叶片的NR活性呈先增加后降低的变化趋势,均为N200处理下燕麦叶片的NR活性最强,牧王旗叶、倒二叶、倒三叶的NR活性分别为75.62,55.64,59.73 μg/(g·h),甜燕1号旗叶、倒二叶、倒三叶的NR活性分别为52.44,37.33,51.86 μg/(g·h),且与其他处理差异显著(除牧王燕麦品种的倒二叶)(P<0.05),牧王不同叶片之间NR活性为旗叶>倒二叶>倒三叶,甜燕1号则规律性不明显。
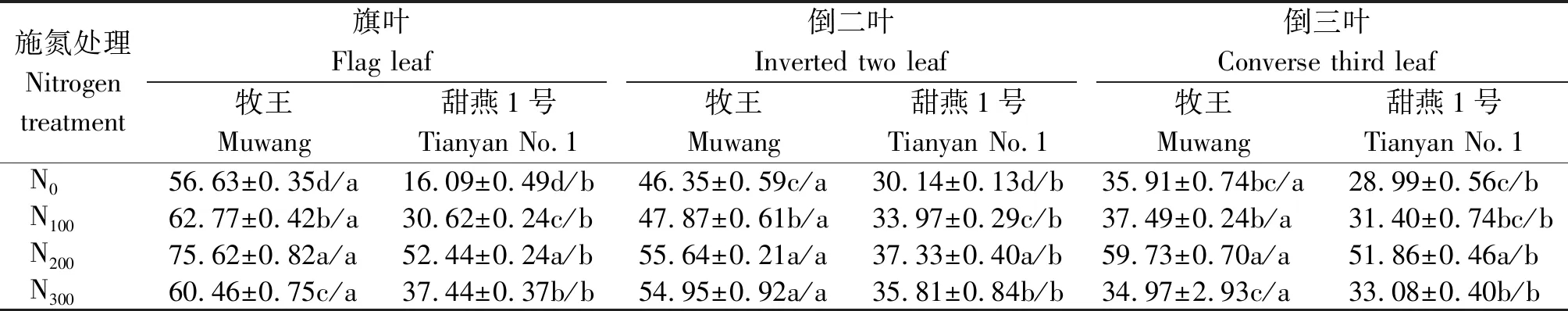
表3 不同施氮量下沙地饲用燕麦叶片NR活性的变化Tab.3 Changes of NR activity in leaves of forage oat in sandy land under different nitrogen application rates μg/(g·h)
2.4 追施氮肥对饲用燕麦GOT活性的影响
由表4可知,牧王品种叶片的GOT活性均高于甜燕1号,但只有旗叶的N0、倒二叶和倒三叶的N300处理差异显著(P<0.05);旗叶和倒二叶的GOT活性随着施氮量的增加呈先增加后降低的变化趋势,倒三叶的GOT则随着施氮量的增加不断增加,除牧王的旗叶和甜燕1号的倒三叶外,其他施氮处理燕麦叶片的GOT活性均显著高于N0(P<0.05),但甜燕1号的旗叶、牧王的倒二叶在不同施肥处理之间差异不显著(P> 0.05),牧王的旗叶和甜燕1号的倒二叶则是N200与N100之间差异不显著(P>0.05),牧王的倒三叶GOT活性为N300显著高于N200和N100处理(P<0.05),甜燕1号的N200处理与N100和N300处理差异不显著(P>0.05),但N300处理显著高于N100处理(P<0.05)。
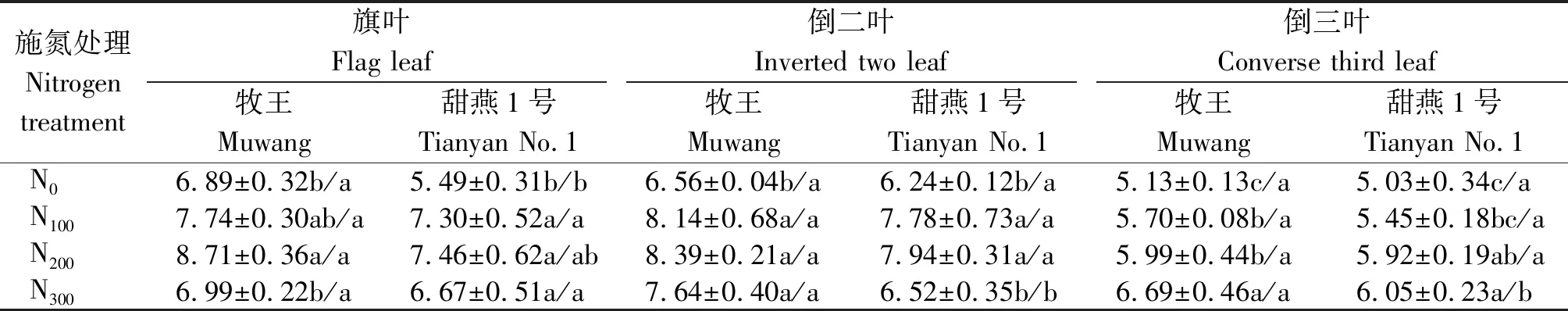
表4 不同施氮量下沙地饲用燕麦叶片GOT活性的变化Tab.4 Changes of GOT activity in leaves of forage oat in sandy land under different nitrogen application rates μmol/(g·h)
2.5 追施氮肥对饲用燕麦GPT活性的影响
由表5可知,牧王旗叶的GPT活性显著高于甜燕1号(P<0.05),倒二叶和倒三叶则差异不显著(P>0.05);随着施氮量的增加,饲用燕麦叶片的GPT活性呈先增加后降低的变化趋势,且均在N200处理下GPT活性最强,牧王旗叶、倒二叶、倒三叶的GPT活性分别为14.85,14.34,12.43 μmol/(g·h),甜燕1号旗叶、倒二叶、倒三叶的GPT活性分别为14.14,15.34,12.06 μmol/(g·h),不同品种在不同氮肥处理下的旗叶、倒二叶、倒三叶的GPT活性均为N200>N300>N100>N0;甜燕1号的旗叶、倒二叶的N200与N300差异不显著(P>0.05),但显著高于N0(P<0.05);牧王旗叶的N200处理显著高于N300和N0处理(P<0.05),倒二叶的N200、N300、N100之间差异不显著(P>0.05),但显著高于N0(P<0.05)。

表5 不同施氮量下沙地饲用燕麦叶片GPT活性的变化Tab.5 Changes of GPT activity in leaves of forage oat in sandy land under different nitrogen application rates μmol/(g.h)
2.6 施氮对燕麦产量的影响
由图1可知,施氮对燕麦干草产量影响显著,但不同燕麦品种间表现不同。随施氮量的增加牧王品种干草产量呈先增加后降低的变化,在N200处理水平下产量最高,为11 460.41 kg/hm2,显著大于其他施氮处理(P<0.05),在N0水平下干草产量最低。甜燕1号品种干草产量在N300处理下最高,为11 786.12 kg/hm2,显著大于其他氮素处理(P<0.05),N100和N200处理间干草产量差异不显著(P>0.05),但显著大于未施氮处理(P<0.05)。
2.7 饲用燕麦产量与不同氮代谢酶的相关性分析
由表6可知,燕麦干草产量与氮代谢酶活性均呈正相关关系,但仅与倒三叶GOT活性呈显著正相关关系,相关系数为0.831,燕麦干草产量与倒二叶和倒三叶GPT活性达到极显著相关关系,相关系数分别为0.925和0.908,而燕麦干草产量与旗叶、倒二叶和倒三叶中的GOGAT、GS、NR活性未达到显著相关水平。
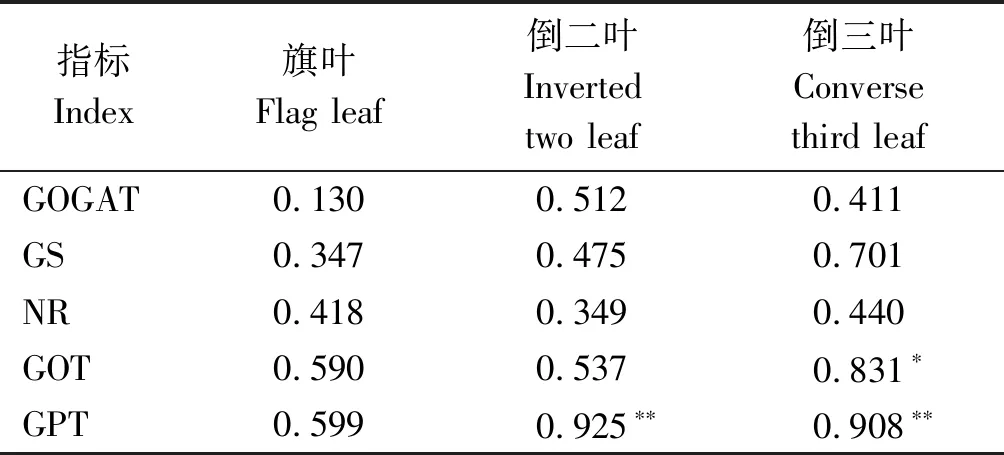
表6 燕麦干草产量与叶片不同部位氮代谢酶活性相关系数Tab.6 Correlation coefficient of oat hay yield and nitrogen metabolism enzyme activities in different parts of leaves
3 讨论与结论
3.1 讨论
GS-GOGAT循环是植物体内铵态氮同化的主要途径[20-23],刘焕[24]研究施氮水平对冬小麦旗叶GS活性的影响表明,GS活性与产量的相关系数达到显著水平;研究施肥对小麦籽实产量及叶片中GS和GOGAT活性影响的文章较多,结果表明,施氮240 kg/hm2时,小麦旗叶的GS、GOGAT等关键酶活性最高[25],籽实产量最高[26],氮高效基因型小麦较高的 GS 活性可促进植株对氮素的吸收与同化,提高氮素利用效率[12];前人研究施氮对玉米叶片中氮代谢相关酶活性影响结果表明,施氮显著提高了玉米叶片GOGAT 和GS氮代谢关键酶活性[27],适宜的施氮水平下玉米功能叶片GS活性较强[28],GOGAT和GS活性随施氮量的增加呈先上升后下降的变化[29]。本研究与其研究结果相似,牧王和甜燕1号饲用燕麦品种不同部位叶片的GOGAT、GS活性随着施氮量的增加均呈先增加后降低的变化趋势。NR是氮素同化的起始酶和限速酶,刘焕[24]研究施氮水平对冬小麦旗叶NR活性的影响,结果表明,在开花期NR酶活性最强,NR活性的变化为N360、N300>N240、N180>N0,但NR活性与产量的相关系数不显著。本研究与其结果相同,相关性分析亦表明,NR与燕麦干草产量相关性未达到显著水平。本研究发现牧王和甜燕1号饲用燕麦品种不同部位叶片的NR活性随着施氮量的增加均呈先增加后降低的变化趋势,且在200 kg/hm2施氮处理下酶活性最强。这与赵吉平等[25]、张弦等[26]、牛巧龙等[30]结果一致,NR随着施氮量的增加呈单峰曲线变化规律。
关于转氨酶与施氮量的关系研究较少,赵吉平等[25]研究施氮量对小麦氮素代谢关键酶活性的影响,结果表明,小麦旗叶的GPT活性随施氮量增加而先升后降,施氮240 kg/hm2时GPT酶活性最高,小麦旗叶氮素代谢关键酶活性与籽粒产量呈显著相关。本研究表明,牧王和甜燕1号饲用燕麦品种不同部位叶片的GOT、GPT活性随着施氮量的增加均呈先增加后降低的变化趋势,且在200 kg/hm2施氮处理下酶活性最强(除N300处理倒三叶的GOT)。相关性分析表明,燕麦干草产量与倒二叶和倒三叶中GPT活性极显著相关,与倒三叶中GOT活性显著相关。
前人研究表明,氮高效利用玉米品种叶片中的GS、GOGAT、NR活性高于氮低效利用玉米品种[14],氮高效品种在不同的氮水平下有较高的氮代谢酶活性[29]。在对氮高效和低效型小麦的研究中表明,氮素高效型小麦品种的 NR 活性和 GS 活性显著高于低效型品种,且受到氮素供应水平的正向调控[31]。在田间栽培条件下,品种间比较发现,氮素高效型小麦品种旗叶 NR、GS 活性和籽粒 GS 活性高于低效型品种。本研究表明,除N100处理的倒三叶GS活性,不同氮肥处理下倒二叶GPT活性外,牧王饲用燕麦品种的GOGAT、GS、NR、GOT、GPT酶活性均高于甜燕1号,除倒三叶的GOT活性外,2个燕麦品种叶片中GOGAT、GS、NR、GOT和GPT活性均在N200处理下最高,且牧王品种在N200处理下干草产量最高,而甜燕1号则在N300处理水平下最高,结合相关性分析结果,研究认为,GOT和GPT是饲用燕麦氮素同化的关键酶,牧王饲用燕麦品种的氮效率高于甜燕1号。
3.2 结论
牧王和甜燕1号饲用燕麦品种不同部位叶片的GOGAT、GS、NR、GOT、GPT活性随着施氮量的增加均呈先增加后降低的变化趋势,且在N200施氮处理下酶活性最强(除N300处理倒三叶的GOT);除N100处理的倒三叶GS活性,不同氮肥处理下倒二叶GPT活性外,牧王饲用燕麦品种的GOGAT、GS、NR、GOT、GPT酶活性均高于甜燕1号;牧王和甜燕1号饲用燕麦干草产量分别在200,300 kg/hm2施氮水平下达到最大;GOT、GPT活性是饲用燕麦氮素同化的关键酶,牧王的氮素同化能力强于甜燕1号。因此,在科尔沁沙地生境下种植牧王饲用燕麦品种追施氮肥的适宜用量为200 kg/hm2,而种植甜燕1号饲用燕麦品种追施300 kg/hm2氮肥可获得较高的干草产量,GOT、GPT活性是筛选氮高效利用饲用燕麦品种的关键酶。
