森林草莓FvCAX 基因家族的鉴定及生物学功能分析
2021-06-29闫慧清刘艳晶孙贵连黄小龙
闫慧清,刘艳晶,2,黄 绒,孙贵连,黄小龙,2*
(1. 贵州师范大学生命科学学院, 贵阳 550025;2. 贵州师范大学, 植物生理与发育调控贵州省重点实验室, 贵阳 550025)
Ca2+是植物生长发育过程中非常重要的营养元素,是构成细胞壁和细胞膜等结构的必需物质[1],能够维护细胞膜与膜结合蛋白的稳定性,减少或延缓膜的损伤[2]。 CAX (Ca2+/H+exchanger antiporter)是一类重要的跨膜转运蛋白,在调节植物钙含量的变化和植物内阳离子转运上起着极其重要的作用[3]。利用生物信息学方法鉴定森林草莓CAX基因家族的成员,研究其染色体分布、进化关系和基因结构、不同发育时期的表达以及启动子的元件分析等,对于研究蔷薇科物种中CAX基因的功能具有重要的意义。
CAX是存在于细胞质膜或液泡膜上的多基因家族成员[4]。前人利用免疫杂交和烟草免疫显色得到AtCAX1~AtCAX4 均定位于拟南芥的液泡膜上[5]。在拟南芥中已通过实验手段验证了 AtCAX1~AtCAX4 的功能,其中,AtCAX1 参与许多生物学过程,如钙离子跨膜转运、对盐胁迫的反应,钙离子转运和细胞内钙离子稳态等[5]。AtCAX2 和AtCAX4 能够增强植物向液泡区储存重金属的能力,具有转运多种阳离子的功能,增强植物体内Ca2+、Cd2+和Mn2+的累积能力,从而提高植物对重金属胁迫的耐性[4]。AtCAX3 能维持Na+和Li+稳态并增强对低pH 的高度敏感性。AtCAX3 与AtCAX1的氨基酸序列相似度高达77%,但是不同成员之间的表达量存在组织特异性,其中AtCAX1在拟南芥的根、茎、叶和花的组织中均有表达,但在叶中高度表达;AtCAX2在维管组织的薄壁细胞、花以及幼嫩的顶端分生组织中表达量高[6];AtCAX3在根和花的组织中表达量高,而AtCAX4只有在根中表达[7]。添加Ca2+可诱导拟南芥AtCAX1、水稻OsCAX1a和棉花GhCAX3的基因表达[8-9]。苹果MdCAX共有8个成员,其基因结构特点表现出转运蛋白的活性,定量分析结果表明MdCAX对不同盐离子胁迫具有不同的表达模式[10]。油菜BnaCAXs家族共含有17个成员,每个成员包含8 ~ 11 个跨膜结构域,进化树表明其与十字花科的拟南芥结构相似。启动子分析表明,油菜BnaCAXs的启动子中,Dof、MYB 以及W-BOX 是主要顺式作用元件,能够参与胁迫反应;定量分析结果表明BnaCAXs主要在地下部表达,其核心家族成员为BnaC04g07160D和BnaC03g37860D[11]。
森林草莓Fragaria vesca(2n= 2x= 14)为多年生草本植物,其基因组约为206 Mb,易于遗传转化,是重要的蔷薇科模式植物。CAX 是重要的植物转化蛋白,同时还能增强对重金属胁迫的耐性,对于植物生长发育和抗逆性具有重要功能,但迄今为止,其在森林草莓基因组中的成员、表达及其调控尚鲜见报道。本研究采用生物信息学分析方法,分析森林草莓CAX基因家族的成员、理化特征、染色体定位、进化、表达及其调控,旨在进一步研究FvCAX基因家族的生物功能及了解其在森林草莓对营养元素吸收中的作用,为研究森林草莓CAX基因家族的功能提供基础信息。
1 材料与方法
1.1 森林草莓FvCAX 基因家族的鉴定
从拟南芥基因组 (https://www.arabidopsis.org/)中获得6 个AtCAX 的蛋白序列。利用TBtools 将拟南芥中获得CAX 家族成员序列与森林草莓基因组的序列NCBI (https://www.ncbi.nlm.nih.gov/genome/)进行同源比较(e=1×10-5)。去除冗余序列后,获得初步的FvCAX家族的核酸和蛋白质序列,并再通过NCBI 中保守结构域的数据库(Conserved domain database, CDD)去除不含有CAX 保守结构域的基因,最终确定FvCAX的家族成员。Gene ID 来源于Fragaria_vesca_v4.0.a2. transcripts 文件,转录本来源于 NCBI。利用 ProtParam (http://expasy.org/tools/protparam.html)获得FvCAX家族成员的氨基酸理化性质,如分子量、等电点和平均亲水系数。通过数据库(http://www.csbio.sjtu.edu.cn/bioinf/plant-multi)获得FvCAX家族成员的亚细胞定位信息。
1.2 FvCAX 基因家族的系统树构建及蛋白序列分析
利用Clustal W 对森林草莓FvCAX 蛋白序列与拟南芥(Arabidopsis thaliana)以及NCBI 中收录的水稻(Oryza sativa)、月季(Rosa cultivars)、苹果(Malus domestica)的CAX 蛋白序列进行多重比较,通过Mega7 软件中邻接算法Neighbor-Joining Tree 构建进化树,其中Bootstrap 重复值为1 000 次。MEME(http://meme-suite.org/tools/meme)在线分析Motif 结构。利用TBtools 软件(Redraw Gene Structure)构建基因结构图。
1.3 FvCAX 基因家族的染色体定位分析
利用NCBI 中森林草莓基因组提供的染色体注释信息,得到并利用R 软件对FvCAX 家族成员的染色体定位进行可视化。根据家族成员在染色体上的位置由低到高进行命名。
1.4 FvCAX 在野草莓发育过程中的表达分析
通过森林草莓转录数据分析FvCAX家族成员在植株发育的整个时期表达模式[12],包括有种子(子房壁、胚、胚乳和种皮组织)、幼苗(种子萌发后10 d的植株)、成熟叶片和花中不同组织(花粉、心皮、花托内部的髓、瘦果下面的肉质组织),并将不同的组织划分为不同的阶段[13]。该数据已提交到SRA数据库(SRA065786),使用RPKM 法表示FvCAX家族成员的基因表达量。最后通过R 软件对FvCAX家族成员的表达量进行聚类。
1.5 FvCAX 的启动子分析
为了阐明调控FvCAX基因家族可能的分子机制,对其启动子进行顺式作用元件的分析。首先通过NCBI 获得FvCAX基因起始密码子(ATG)上游2 000 bp 基因组DNA 序列,并用NEW PLACE(https://sogo.dna.affrc.go.jp/cgi-bin/sogo.cgi)数据库鉴定启动子区域中的顺式作用元件。
2 结果与分析
2.1 森林草莓FvCAX 家族的鉴定及特性分析
通过对森林草莓的基因组注释序列和CDD 数据库的筛选分析,共获得了森林草莓FvCAX家族包含有7 个成员(表1)。根据其在染色体上的位置(图1),对其成员进行编号为FvCAX1~FvCAX7。由图1 得到森林草莓共有7 条染色体,其中FvCAX1(Chr3:12773449—12777622)、FvCAX2(Chr3:22613312 — 22617539) 位于3 号染色体;FvCAX3(Chr6:7635361—7644208)、FvCAX4(Chr6:7649624-7652926) 、FvCAX5(Chr6:26474172—26479381)、FvCAX6(Chr6:26479432—26484248)和FvCAX7(Chr6:27461045—27466466)位于第6 号染色体上,形成了基因簇。
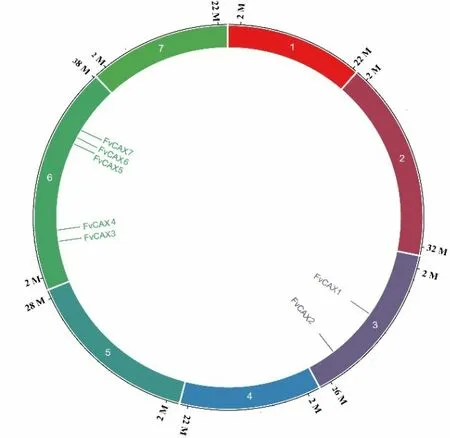
图1 森林草莓FvCAX 基因家族中7 个成员的染色体定位Figure 1 Chromosome location of seven FvCAX genes
通过对FvCAX家族中蛋白质成员的理化性质分析得到,森林草莓FvCAX基因家族蛋白质的相对分子质量差异较小,位于47.02 ~ 59.41 kDa 之间,其中FvCAX6 的分子质量最大,为59.41 kDa,而FvCAX3最小,为47.02 kDa,等电点在4.90 ~ 5.81 之间,表明森林草莓中Ca2+/H+跨膜蛋白均为酸性。由表1 可知,FvCAX基因家族的蛋白质的平均亲水系数(GRAVY, grand average of hydropathicity)值为0.345 ~0.645,表明所有的成员均为疏水蛋白,且疏水程度存在差异。亚细胞定位预测表明森林草莓FvCAX基因家族中有4 个成员定位在细胞质膜上,其余3 个基因定位在叶绿体类囊体膜上,表明不同成员对细胞质和叶绿体中Ca2+的运输有不同的分工。
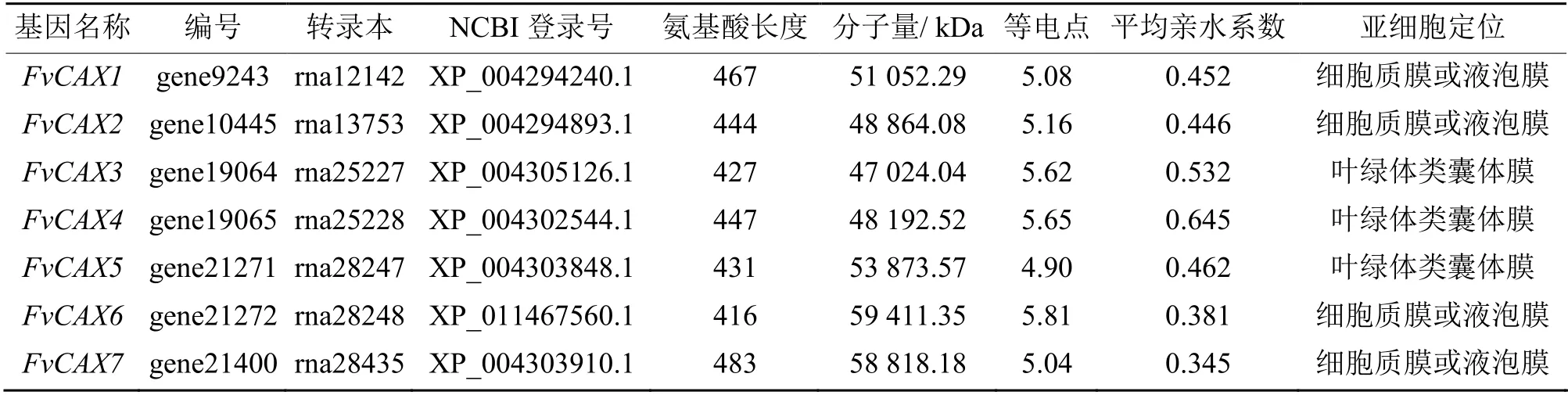
表1 森林草莓FvCAX 基因的理化分析Table 1 The physical and chemical analysis of FvCAX gene family in Fragaria vesca
2.2 森林草莓FvCAX 家族的进化关系及模体分析
通过搜索得到拟南芥(AtCAX1~AtCAX6)、月季(RcCAX1~RcCAX9)和苹果(MdCAX1~MdCAX8)的CAX序列,并将得到的序列与森林草莓构建系统发育进化树(图2)。由图2 可知,7 个FvCAX 蛋白与拟南芥、月季和苹果的CAX 形成独立的四大分支,而且草莓的每个FvCAX 蛋白全部都与月季的亲缘关系最近,其次是苹果和拟南芥。这些结果表明森林草莓中不同的FvCAX 之间存在差异,且每个FvCAX 都能与月季中的RcCAX 功能相似。
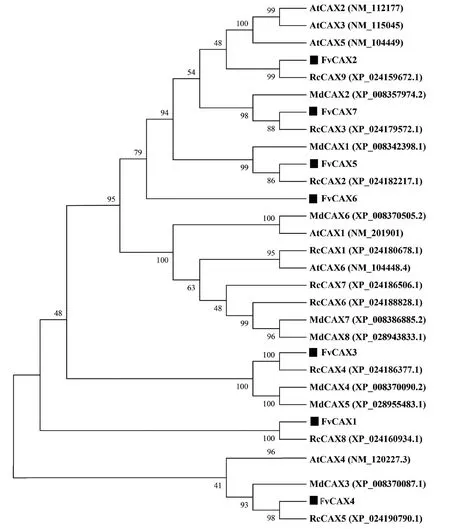
图2 森林草莓FvCAXs 与其他物种CAXs 的系统进化树Figure 2 Phylogenetic tree of the amino acid sequences of FvCAXs and CAXs in other species

图3 森林草莓FvCAX 基因中7 个成员的基因结构和蛋白保守结构域分析Figure 3 Gene structure and protein conserved domains of seven FvCAX genes
利用森林草莓FvCAX的基因组序列和开放阅读框(Open reading frame),确定了森林草莓FvCAX基因的UTR (Untranslated region)、CDS (Coding sequence)和内含子的分布,FvCAX基因家族各成员之间的进化关系如图3A,内含子数量在10 ~ 11 个且内含子在0、1 和2 相位均有出现(图3B)。利用MEME(http://meme-suite.org/tools/meme)对森林草莓CAX基因家族的motif 序列进行分析。结果表明:森林草莓7 个FvCAX基因家族共含有5 个保守的motif(图3C);将motif 的一致序列列出,结果如表2 所示。
此分析结果可以说明FvCAX基因家族中7个基因都是FvCAX基因家族具有差异功能的基因,有助于预测FvCAX基因家族的生物功能。

表2 森林草莓FvCAX 蛋白的预测motifTable 2 The sequences of motif consensus of FvCAX protein

图4 森林草莓FvCAX 基因家族在不同组织和发育时期的表达模式(括号中的数字表示发育阶段)Figure 4 Expression patterns of seven FvCAX genes in various tissues and development (The number in the bracket indicates the developmental stages)

图5 森林草莓FvCAX 基因启动子区域结合的与胁迫相关顺式作用元件分析Figure 5 Identification of cis-acting regulatory elements related to stresses in the promoter region of the FvCAX gene family
根据保守motif 的结构,所得的结果如表2 所示。其中motif 1 (MFAMKDKLDITLGVAIGSSTQISMFV IPFCVVVGWIMGIPMDLNFQLFET)和motif 3 (FGWL CIMTIWISWLSEYVVDTIEGASDSWNIPVAFISVIL LPIVGNAAEH)都具有运输Ca2+的功能。motif 5 的蛋白具有跨膜功能。
2.3 森林草莓FvCAX 家族的表达分析
FvCAX家族中5 个成员在不同的组织及同一组织的不同发育阶段具有不同的表达量,表明这些成员发挥不同的功能,具体如图4 所示。根据其表达模式的变化,可以将基因表达情况分为两大类,第一类基因包括FvCAX1~3 和FvCAX5,在整个植株发育时期表达量较低,仅有部分基因在花发育的部分时期有表达量;第二类基因在所有组织及其发育阶段具有显著的表达量,尤其是FvCAX6和FvCAX7,在苗期、叶片、花的发育过程和种子的形成过程中表达量都显著性增加,表明这两个基因对森林草莓发育过程中Ca2+的吸收具有重要的作用。同时发现FvCAX7在同一个组织的不同发育时期表达量存在差异,FvCAX7在花药发育的第10 阶段含量最高,在第11 和12 阶段表达量降低,表明森林草莓不同发育阶段对Ca2+的需求存在差异。
2.4 森林草莓FvCAX 家族的启动子分析
转录因子能够结合到FvCAX启动子的顺式作用元件上,对调控转录水平的表达起到重要作用。为了研究森林草莓CAX家族基因启动子区域所能结合的顺式作用元件,我们利用PLACE (Plant cis-acting regulatory DNA elements)数据库对FvCAXs基因启动子进行了分析。有34 个顺式作用元件能结合到FvCAX家族基因的启动子上,其中有22 个元件均能结合到7 个家族成员中,除了常见的启动子,如TATA box 和CAAT box 之外,其中丰度较大的主要有MYB、Dof (AAAG)、WRKY、W-box 和MBS (图5A)。这些元件在调控植物基因响应外界胁迫发挥了重要作用,结果如图5B 所示。其中最大的为MYB (31%),其次为DOF (24%)和WRKY (23%)。
3 讨论与结论
CAX 是含有多个基因的基因家族,其中水稻有6 个,苹果含有8 个,油菜有17 个[14]。通过同源搜索,在森林草莓基因组中鉴定出7 个CAX基因。在不同的物种中,FvCAX 的理化性质存在差异,如甘蓝类中CAX 等电点变化范围是4.71~ 6.67,苹果的CAX 等电点为4.97 ~ 6.67[10-11],本研究结果指出草莓的等电点介于4.90 ~ 5.81。此外,作为膜的重要转运蛋白,CAX 定位于质膜、液泡膜、叶绿体类囊膜和线粒体膜上[15],但在不同物种中存在差异。前人报道拟南芥的6 个CAX 成员主要定位在液泡膜和细胞膜上[16],苹果的CAXs 定位于质膜上或者内质网膜上[17]。本研究中,对野草莓的CAXs 定位于质膜、液泡膜和叶绿体类囊膜,表明可能野草莓的植物细胞基质内和叶绿体内Ca 需求较多,且对于基质和叶绿体内阳离子的转运发挥重要的作用。
FvCAXs具有组织特异表达特征,表明家族中不同成员存在功能差异,可能存在不同的阳离子转运功能。FvCAX1~3 仅在花发育时期有表达;FvCAX6和FvCAX7在植株整个发育期都有表达,尤其是在苗期(种子萌发后10 d)和成熟的叶片中。根据其表达模式的高低,我们确定了FvCAXs家族的核心基因为FvCAX6和FvCAX7。苹果MdCAXs在不同的营养器官中,如根、茎和叶中的表达模式也不同,且受相同的外界重金属胁迫处理后,成员表现出不同变化量的改变,表明MdCAXs呈现出特异性应答反应[10]。棉花GhCAX表达丰度和模式均有所差异,其中GhCAX3主要在叶、茎、柱头、花瓣和胚珠中表达,在根和纤维组织中的表达量较少[18]。在烟草中过表达AtCAX4能显著提高植物根部Cd2+累积[19]。同时,在Cd2+胁迫条件下,AtCAX4的表达显著提高[20]。甘蓝型油菜BnaCAX家族中的两个主效基因同样受到Cd2+胁迫的诱导。关于森林草莓FvCAX基因家族在Cd2+胁迫下的表达值得进一步研究。
启动子所包含的顺式调控元件能够在转录水平上调控基因的表达。水稻OsCAX4基因表达量在盐胁迫条件下显著上升[21];在烟草和大豆中过表达CAX1提高了植物对盐胁迫的耐受性[12]。此外,GhCAX3对于棉花适应低温条件必不可少[22]。通过数据库预测得到共有22 个顺式作用元件均结合到FvCAX家族基因的启动子,丰度较大的包括MYB、Dof (AAAG)、WRKY、W-box 和MBS (图5B),这些元件在调控基因响应逆境胁迫中均发挥了重要作用。如水稻和拟南芥的WRKY受到高低温、盐碱、干旱和营养缺素等逆境胁迫因子而被诱导表达[23]。水稻OsDof13受低氮胁迫后呈现先下降再升高的表达变化[24],表明FvCAX基因表达量能够在不同环境胁迫下被诱导表达。因此,前人研究表明CAX家族在调控植物应对胁迫逆境发挥重要作用,与MYB、Dof (AAAG)、WRKY、W-box 和MBS 顺式作用元件有关。本研究基于基因组序列共鉴定出7个森林草莓FvCAX家族成员,并且与月季的亲缘关系最近,通过分析结构域和MEME 蛋白基序表明,FvCAX是保守的,并结合不同组织和发育时期的转录组数据得到该家族成员的基因表达模式。本研究结果可为进一步研究草莓FvCAX 蛋白家族功能提供理论依据,为研究森林草莓中CAX基因家族的功能提供理论基础。
