全基因组关联研究在多囊卵巢综合征中的研究进展
2021-06-29马丽娜马红丽高敬书吴效科张多加王珑
马丽娜,马红丽,高敬书,吴效科,张多加,王珑
(1.黑龙江中医药大学,哈尔滨150040;2.黑龙江中医药大学附属第一医院a.妇科,b.针灸科,哈尔滨150040)
多囊卵巢综合征(polycystic ovary syndrome,PCOS)是一种复杂的生殖内分泌紊乱疾病,影响全球6%~10%女性的健康[1],以亚洲和欧洲人群更常见[2]。PCOS病因复杂、分子发生机制不明。多数研究认为,遗传和环境因素协同作用是导致PCOS发生发展的主要原因[3-6]。因此,寻找PCOS发病的遗传和环境机制是近年的研究热点。家系研究和双胞胎研究均指出,PCOS具有遗传倾向,大量的候选基因研究也证实了200余个PCOS易感基因[7-8],提示PCOS遗传背景的复杂性。但由于PCOS复杂的病理生理过程涉及多种路径和蛋白质,以上单一的基因诊断试验无法阐明其遗传病因及发病机制,故需要寻找一种更全面高效、客观性强的遗传学研究方法。全基因组关联研究(genome-wide association study,GWAS)通过连锁不平衡分析捕获大部分基因组变异,然后利用一组具有代表性的单核苷酸多态性(single nucleotide ploymorphism,SNP)评估全基因组变异,这为复杂疾病的遗传学研究提供了更全面、客观的分析策略。GWAS在卵巢癌、子宫内膜异位症、宫颈癌、子宫肌瘤等女性生殖系统损害性疾病的易感基因座识别上取得了突破性进展[9-12]。近年GWAS已被用于不同种族人群的PCOS致病基因研究。现就GWAS在PCOS中的研究进展予以综述,以期为PCOS的遗传生物学机制研究、精准治疗提供新的见解和方向。
1 PCOS的遗传学研究基础
临床上,PCOS发病呈家族聚集性,被认为具有一定的遗传学基础。一项PCOS家系研究的荟萃分析发现,与正常对照女性的一级亲属(父母、兄弟姐妹)相比,PCOS患者的一级亲属(父母、兄弟姐妹)出现代谢综合征、高血压和血脂异常的风险显著增加[13]。另一项针对3 100多对双胞胎进行的大规模调查问卷结果显示[14],PCOS患病率为2.97%(92/3 100),其中单卵双胎的PCOS患病率是双卵双胎的2倍,提示遗传因素在PCOS发生中起重要作用。以上研究表明,遗传因素与PCOS发病关系密切,但基于复杂疾病的固有特点及PCOS本身的特点家系连锁分析等遗传学方法作用受限,故研究者开始采用GWAS探索PCOS致病基因,即从家系、双生子研究逐步过渡到候选基因关联研究。
PCOS候选基因关联研究主要是选择可能与PCOS病理、生理相关的基因进行关联性分析。至今,研究者已对200余个PCOS的候选基因进行了关联研究[7-8],包括雄激素生物合成、储备及作用相关基因(17α-羟化酶基因、雄激素受体基因、性激素结合球蛋白基因等)、性腺功能失调相关基因(黄体生成素/人绒毛膜促性腺激素受体基因、卵泡刺激素受体基因等)、代谢相关基因(胰岛素基因、胰岛素受体底物基因、胰岛素受体基因、体脂含量和肥胖相关基因等)及慢性炎症相关基因(白细胞介素-6基因、肿瘤坏死因子-α基因等)。由于研究的样本量偏小(受试者少于300名)以及仅评估基因中个别变异的影响,并非调查整个基因的变异等不足和局限,大多数功能候选基因关联性结果缺乏一致的复制。因此,对PCOS致病基因的探索不能仅局限于单基因筛查研究模式。
2 GWAS识别PCOS常见基因多态性
GWAS以应用基因组中的SNP作为分子遗传标记,进行全基因组水平上的对照或相关性分析,并通过比较发现影响复杂性状的基因变异、确定疾病遗传风险因素的一种新策略。GWAS可用于2型糖尿病、肥胖、哮喘、精神疾病等具有复杂性状疾病的研究[15-17],并可用于易感基因座的确定。目前,通过GWAS对包括中国汉族女性、韩国人和欧洲血统人群PCOS患者进行检测的8项相关实验检测出20个PCOS易感位点,易感区域相关基因主要参与卵泡成熟和排卵(LHCGR、FSHR、FSHB、MAPRE1、ERBB4、SOD2)、高雄激素相关(DENND1A、ZBTB16)、胰岛素信号通路(INSR、PLGRKT)、1型糖尿病(RAB5B)、2型糖尿病(THADA和HMGA2)、男性勃起功能障碍(C9orf3)、器官大小和细胞增殖控制(YAP1和SUMO1P1)、染色质重组(TOX3)、细胞周期调控(KHDRBS3)、免疫和炎症反应(GATA4/NEIL2)、胚胎发育(KRR1)。
GWAS发现的20个易感基因中,许多在PCOS既往遗传学研究中并未受到关注,如DENND1A、KHDRBS3、YAP1基因。DENND1A基因含有22个外显子和DENND1A.V1、DENND1A.V2两个转录体,编码一种含DENND结构域和clathrin结合结构域的蛋白——Connecdenn1蛋白,可作为Rab35的鸟嘌呤交换因子,与clathrin及适应蛋白2结合,介导细胞内吞,参与调控蛋白质和脂质代谢。有研究表明,PCOS女性卵巢卵泡膜细胞中DENND1A.V2基因过表达,导致17α-羟化酶基因启动子活性改变,促进雄激素合成和分泌,进而介导PCOS患者的高雄表型[18],故推测DENND1A基因对PCOS女性雄激素升高有重要影响。因此,了解上述基因及大量遗传位点SNP的生物学功能对PCOS分子机制的研究具有重要意义,同时也是未来GWAS研究的重点。
3 基于不同种族人群的PCOS-GWAS
3.1 基于汉族人群队列的PCOS-GWAS Chen等[19]首次将GWAS应用于PCOS遗传学研究开展了第一个基于中国汉族女性的PCOS-GWAS,发现了3个与PCOS发病相关的易感区域:2p16.3、2p21和9q33.3,相应位点附近的风险候选基因有LHCGR、THADA和DENND1A,此研究被认为是PCOS遗传学研究领域里程碑式的发现,且相关位点在随后研究中亦得到了证实。一项对2 082例PCOS患者基因型-表型的研究发现,THADA、DENND1A基因区SNP可能通过代谢紊乱相关途径在PCOS发生发展中起作用[20]。Dallel等[21]在突尼斯妇女中发现DENND1A基因rs10818854位点和rs10986105位点与PCOS发病相关。此外,Zou等[22]通过荟萃分析进一步证实,LHCGR基因变异与PCOS特异相关,推测LHCGR可用作预测PCOS风险的潜在生物标志物。
为进一步完善对PCOS遗传因素的认识,Shi等[23]扩大样本量开展了第二个基于汉族人群的PCOS-GWAS。此次研究在证实了第一个GWAS识别的3个位点外,又发现8个新的PCOS的易感区域,即2p16.3(FSHR基因)、9q22.32、11q22.1、12q13.2、12q14.3、16q12.1、19p13.3和20q3.2。上述研究结果为探索PCOS的遗传因素(致病基因)提供了线索,引起学术界的广泛关注,随后该研究方法不断被应用于相关研究,但并非GWAS发现的所有SNP均得到了证实。在欧洲、沙特和印度人群中已经验证2p21(THADA)、12q14.3(HMGA2)、2p16.3(LHCGR)和9q33.3(DENND1A)与PCOS发病相关[24-26]。
3.2 基于韩国人群队列的PCOS-GWAS 另一项基于亚洲血统的PCOS-GWAS在韩国女性中开展。Hwang等[27]的研究显示,并未识别达到GWAS显著性(P<10-8)的易感位点。分析原因可能是该研究样本量相对较小,且健康对照组女性并非完全健康,其中10.5%的女性伴高雄激素血症,导致研究结果与中国PCOSGWAS[19,23]研究结果存在差异。随后,Lee等[28]扩大样本量再次进行了基于韩国女性的PCOS-GWAS,结果发现了1个新的PCOS易感位点8q24.2(rs10505648,OR=0.52,P=5.46×10-8),其他微相关易感区域位于4q35.2、16p13.3、4p12、3q26.33、9q21.32、11p13和1p22[P=(5.72~6.43)×10-5]上。rs10505648位于含KH的RNA结合结构域信号转导相关分子3(KH domain containing,RNA binding,signal transduction associated 3,KHDRBS3)基因上游,KHDRBS3基因与端粒酶活性密切相关。端粒酶是一种维持端粒长度的酶,其长度与PCOS发病、临床表现密切相关[29-30]。故推测,KHDRBS3基因可能通过影响端粒酶活性调控端粒酶长度,进而导致PCOS发生及临床表现的异质性。但之前基于中国PCOS人群[19,23]发现的一些位点并未在Lee等[28]研究中得到验证,其原因可能与中国和韩国的PCOS-GWAS使用的基因分型芯片(中国研究:Affymetrix SNP 6.0芯片,韩国研究:Illumina HumanOmni1-Quar v1阵列)不同有关。
3.3 基于高加索/白种人血统人群队列的PCOS-GWAS
为探求既往被识别的与PCOS发病相关位点[19,23,27-28]能否在欧洲人群中得到验证,并试图发现新的位点,欧洲学者在高加索人群和白种人中展开了PCOS-GWAS。Hayes等[31]在欧洲后裔女性中进行了大型的三阶段GWAS发现并确定了与PCOS相关的3个易感基因区域:8p32.1、11p14.1(FSHB)以及9q22.32,其中,8p32.1和11p14.1(FSHB)两个位点为新发现的位点。随后,Hong等[32]在韩国人群队列研究中验证了位于FSHB基因上的SNP与PCOS发生发展密切相关,该研究进一步证实作为FSHR基因补充的FSH信号配体——FSHB基因与PCOS的发病机制相关,且与种族无关。配体(FSHB基因)和受体(FSHR基因)均增加了PCOS的患病风险,体现了神经内分泌在PCOS发病机制中的重要性。
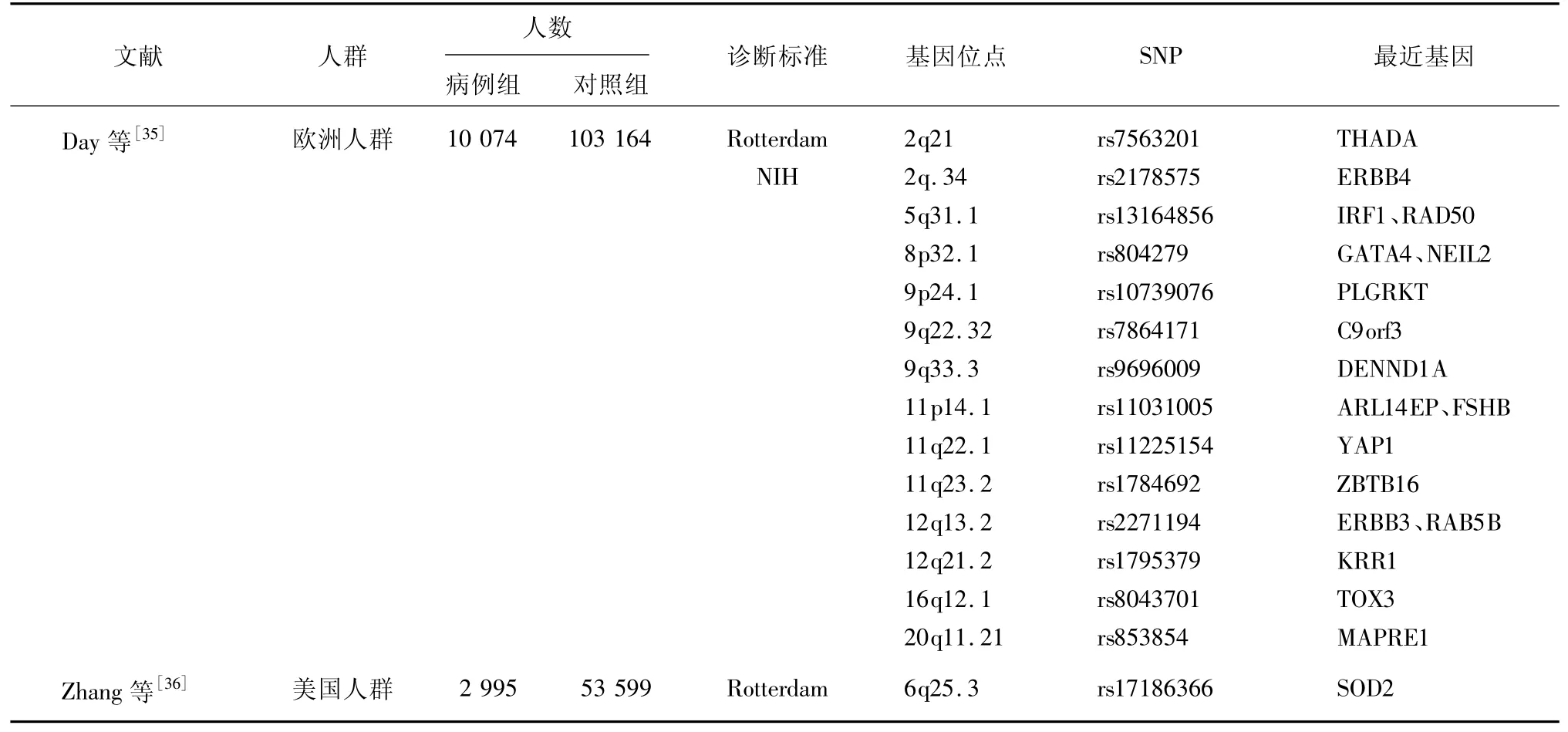
Day等[33]基于欧洲白人鉴定了6个PCOS的易感位点,包括ERBB4/HER4、YAP1、THADA、FSHB、RAD50和KRR1,其中YAP1和THADA在既往GWAS[19,23]中已有报道;FSHB在Hayes等[31]的GWAS中也有报道,并在韩国人群[32]中得到验证;而ERBB4/HER4、RAD50、KRR1三个位点是基于欧洲人群新发现的与PCOS发病相关的易感位点。随后,有学者在中国汉族人群中验证ERBB4(rs1351592)与PCOS发病的相关性,结果证实ERBB4基因与中国汉族女性PCOS发生发展密切相关[34]。故可推测,不同种族PCOS女性可能具有相似的基因结构。
3.4 其他PCOS-GWAS 一项基于欧洲血统队列PCOS-GWAS国际荟萃分析[35],鉴定出3个新的与PCOS发病相关位点(靠近PLGRKT、ZBTB16和MAPRE1)基因。新位点变异可能参与PCOS女性胰岛素信号通路、高雄激素相关和脂肪代谢及卵泡发育过程,但这3个新位点尚需在其他不同种族人群中进一步重复和验证。以上在亚洲和欧洲女性中进行的大规模GWAS[19,23,27-28,31,33,35]已经确定了19个基因座与PCOS相关,但研究使用了不同的诊断标准及自我报告信息。虽然大多数易感位点在不同种族间得到证实,但不同研究中的病例选择偏差和异质性客观存在,为减少这种偏差和异质性,Zhang等[36]通过鹿特丹准则算法对电子健康记录识别的PCOS患者和健康对照女性进行GWAS发现了1个新的PCOS发病相关位点——SOD2 rs17186366。另有研究显示,SOD2中的A16V(rs4880)与中国汉族女性的PCOS、黄体生成素水平以及黄体生成素/卵泡刺激素比值相关[37]。SOD2基因功能及作用机制还有待进一步探索,目前通过PCOS-GWAS已发现至少20个与PCOS发生发展相关的位点,其中11个位点已在不同种族间得到了反复证实和复制,见表1。

表1 PCOS-GWAS鉴定的SNP
4 GWAS的局限性
GWAS运用基因芯片技术筛选出了新的风险位点,为PCOS的生物学机制研究提供了新的方向,但仍存在一定的局限性:①GWAS发现的是易感区域(易感位点),并非致病基因,且多数易感位点并不位于已知基因的开放阅读框,而是位于非编码区或内含子上,给研究带来一定困难。②GWAS能识别大量与PCOS发病相关的常见变异,但常见变异只能解释复杂性疾病的小部分遗传信息,并不能完全解释疾病,进一步探讨罕见变异对疾病的影响十分必要。③目前GWAS过度依赖统计学分析,较容易出现假阳性及假阴性结果,这也是不同种族队列研究结果出现不一致或相反结论的原因之一。所以,识别致病变异和基因并了解它们在PCOS病理机制中的作用是PCOS的GWAS面临的困难之一,即易感基因精细定位和基因功能研究,可能有助于提高人们对PCOS病因和病理机制的认识。
续表1
5 小 结
PCOS是常见的育龄女性生殖内分泌紊乱性疾病,影响人体多个系统和组织。但其确切病因、病理机制尚不清楚,严重影响了PCOS的诊断和精准治疗。家系研究和双胞胎研究明确了PCOS具有遗传基础,且大量候选基因的关联研究也证实了200余个PCOS发病相关的易感基因,提示了PCOS遗传背景的复杂性。GWAS运用基因芯片技术筛选出新的风险位点,至今已有20个与PCOS患病相关的易感区域被识别,为PCOS易感基因的研究奠定了基础,极大地推动了PCOS的遗传学研究。然而,目前有限的信息和证据尚无法全面阐释PCOS确切的遗传方式和分子机制。相信随着生物技术的进步及研究的深入,PCOS病因、病理机制将逐渐清晰,从而为PCOS的精准治疗提供帮助。
