快中子诱变突变体的表型鉴定及配合力效应分析
2021-06-28刘忠祥杨彦忠王晓娟连晓荣周文期何海军周玉乾寇思荣
刘忠祥, 杨彦忠, 王晓娟, 连晓荣, 周文期, 何海军,周玉乾, 寇思荣
(甘肃省农业科学院作物研究所, 兰州 730070)
突变体是进行基因功能研究的优异材料。辐射诱变育种技术是常规育种的重要补充,利用其可以创制出新材料、新种质。因此,利用物理及化学诱变,创制新的变异材料,既可以为作物遗传育种提供基础材料,也对开展突变基因定位、克隆、功能解析、抗逆性机制研究及配合力效应分析等具有重要意义。目前,国内外利用辐射诱变技术取得成效的主要有X射线、γ射线、中子、离子束及多种因素结合的空间诱变[1]。根据FAO/ IAEA(联合国粮农组织/国际原子能机构)的突变品种数据库(http://mvgs.Iaea.org/)统计,截至2016年5月,世界上60多个国家在214种植物上利用诱发突变技术育成和推广了超过3 200个品种,一批有影响力的农作物突变品种和一批优异种质资源在农业生产和品种改良方面得到利用[1-2]。20世纪70年代以来,我国利用γ射线等诱变技术在玉米上育成了原武02、原辐17、丹340、丹360、辐746、鲁原92等自交系,组配了掖单13、鲁单5号、鲁单50等鲁单系列杂交种,推动了玉米生产发展[3-5]。中子不带电荷,通过与物质相互作用产生次级电子使物质电离。中子穿透能力较强,与X射线、γ射线、紫外线相比,有更高的辐射诱变率和更宽的变异谱[6-7]。近年来,国内外学者利用快中子辐照已在豌豆[8]、玉米[9-10]、小豆[11]、紫花苜蓿[12]、花生[13]、棉花[14]、水稻[15]和小麦[16]等多种植物中获得了各具特色的突变体。
课题组前期研究中通过252CF裂变快中子源辐射诱变鉴定筛选得到黄绿叶突变体fu17138、fu18398和fu18362,叶片皱缩、叶夹角减小突变体fu1603,矮秆、低穗位突变体fu18312和fu18321,这些突变体的获得为种质创新、品种改良、基因克隆及其功能分析创造了良好的基础材料,也为育种应用提供了有价值的种质资源。本研究以昌7-2、Mo17、B73、LY0986和LY1621为测验种父本(P1),以6个突变体fu18312、fu1603、fu17138、fu18321、fu18398和fu18362为被测系母本(P2),采用NCII设计(不完全双列杂交组合设计),对其11个农艺性状的配合力进行分析,评价6个突变体的一般配合力、特殊配合力效应及育种利用价值,为品种选育奠定了种质基础,为快中子诱变突变体的进一步改良和利用提供了理论依据。
1 材料与方法
1.1 试验材料
以昌7-2(引进郑单958父本、黄改系)、Mo17(美国引进、Lancaster种群)、B73(美国引进、Reid种群)、LY0986(自育Lancaster种群)和LY1621(自育Reid种群)为测验种父本(P1),快中子诱变创制的突变体fu18312、fu1603、fu17138、fu18321、fu18398和fu18362为被测系母本(P2)。其中fu18312、fu18321、fu1603和fu18398是甘肃农业科学院作物研究所利用252CF裂变快中子源辐射诱变自交系LY8405选择培育而成,fu17138和fu18362是利用252CF裂变快中子源辐射诱变玉米自交系PH6WC分离得到。fu18312和fu18321表现为低穗位、矮秆特性;fu1603表现为叶片皱缩、叶夹角减小特性;fu18398、fu17138和fu18362表现为黄绿叶特性。
1.2 试验方法
试验按 NCII设计方法[17-18],2017年将6个母本和5个父本组配成30个(6×5)杂交组合,于2018—2019年播种于甘肃省农业科学院张掖试验基地。采用随机区组排列,重复3次,行长5 m,行距0.5 m,株距0.2 m,种植密度为10×104株·hm-2,单行区,每小区取样10株,调查株高、穗位、穗长、秃尖、穗粗、轴粗、雄穗分枝数、穗行数、行粒数、出籽率和单株粒重等11个性状,以小区平均数为单位按不完全双列杂交设计进行配合力效应统计分析。
NCII设计以固定模型计算亲本的一般配合力(general combining ability,GCA)及组合的特殊配合力(special combining ability,SCA)的相对效应;随机模型估算群体的遗传力。计算公式如下。
(1)
(2)
(3)
(4)

1.3 数据处理与分析
采用Microsoft Excel 2010软件进行试验数据整理,DPSC数据处理系统进行统计分析和遗传效应分析。
2 结果与分析
2.1 玉米快中子诱变突变体的表型鉴定
2.1.1玉米黄绿叶突变体fu17138的表型鉴定
与野生型PH6WC相比(图1),玉米黄绿叶突变体fu17138在第5叶期开始出现叶片或叶缘黄化的现象;株高和穗位高显著降低;雄穗分枝数增多,由原来的0~1个增加为5~7个;植株能够正常生长并结实,但穗长和穗粒数显著减少。基因定位将叶片或叶缘黄化基因定位于玉米第3染色体(chr3: 173 087 201~182 203 992)约9.2 Mb的区间内[19]。
2.1.2玉米叶夹角突变体fu1603的表型鉴定
与野生型LY8405相比(图2),叶夹角突变体fu1603株型更加紧凑,叶片在5叶期以后出现皱缩卷曲,叶夹角减小,籽粒由马齿型变为硬粒型。基因定位将控制叶夹角的基因定位于玉米第1染色体1.02 bin标记C1-18和C1-2之间(17 966 407~26 885 514)约9 Mb范围内[20-21]。
2.1.3矮秆低穗位突变体fu18312的表型鉴定
野生型LY8405的株高为256.6 cm,穗位高70.7 cm;而矮秆低穗位突变体fu18312的株高仅140.6 cm,穗位高27.1 cm,显著低于野生型(图3A、B和C);果穗变小、籽粒由马齿型变为硬粒型(图3D和E)。

A:拔节期;B:籽粒;C:成株期;D:雄穗;E:果穗。A: Jointing stage; B: Grain; C: Adult stage; D: Male spike; E: Ear.图1 玉米黄绿叶突变体fu17138的表型Fig.1 Phenotype of maize yellow green leaf mutant fu17138

A: 拔节期;B: 成株期;C: 籽粒。A: Jointing stage; B: Adult stage; C: Grain.图2 玉米叶夹角突变体fu1603表型Fig.2 Phenotype of maize leaf angle mutant fu1603
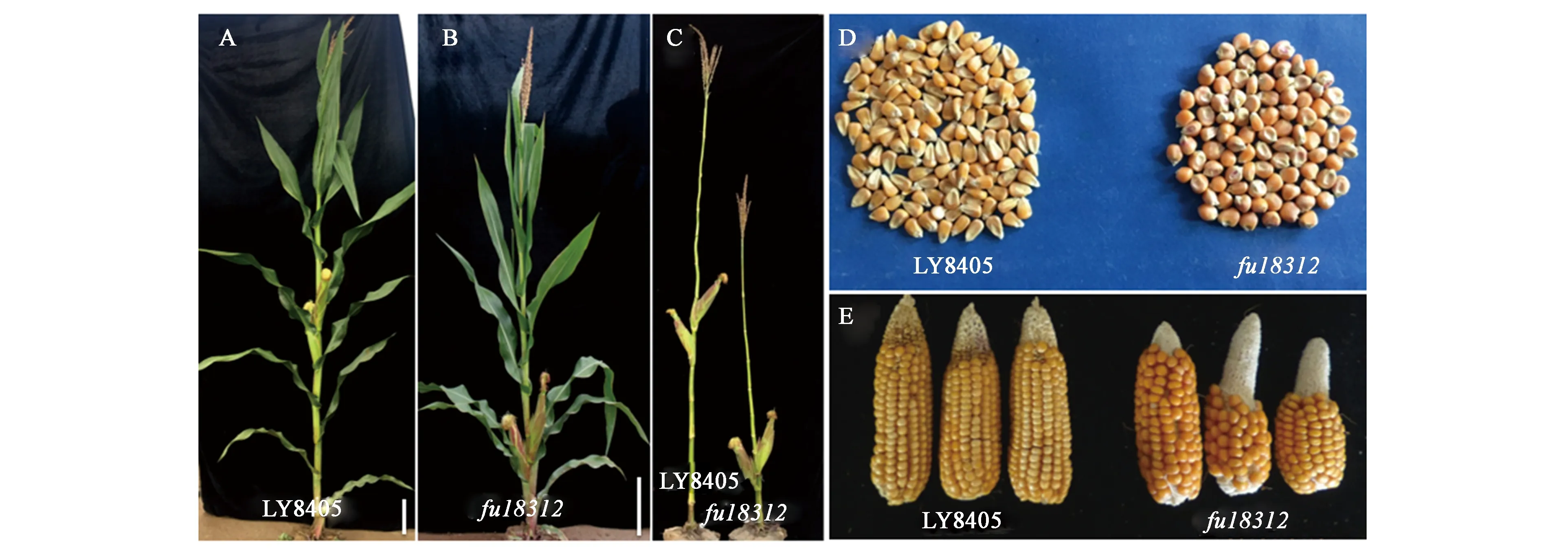
A:LY8405成株期;B:fu18312成株期;C:fu18312和LY8405成株期节间及穗位;D:籽粒;E:果穗;图A/B中标尺=20 cm。A: Adult stage of LY8405; B: Adult stage of fu18312; C: Internodes and ear height of LY8405 and fu18312; D: Grain; E: Ear; Bars in A and B are 20 cm.图3 矮秆低穗位突变体fu18312表型Fig.3 Phenotype of low spike mutant fu18312
2.2 突变体的配合力效应分析
2.2.1主要农艺性状的方差分析 对组配的30个杂交组合11个农艺性状进行方差分析,结果(表1)表明,区组间各性状差异均未达到显著水平;但组合间11个性状差异均达到极显著水平。亲本一般配合力方差和亲本间特殊配合力方差分析结果显示,P1(父本)除穗轴粗差异不显著外,其余性状均达显著或极显著差异;P2(母本)除雄穗分枝数和秃尖长外,其余性状均达到显著或极显著差异;而P1×P2除穗轴粗外,其余性状均达显著或极显著差异。由此可见,这10个性状的遗传均受加性效应和非加性效应共同作用。

表1 各性状的方差分析(均方值)Table 1 Analysis of variance for every trait
2.2.2供试材料11个性状的一般配合力效应分析 不同亲本11个性状的GCA效应值有很大的差异(表2)。自交系昌7-2雄穗分枝数、穗位高、秃尖长和出籽率的GCA效应值分别为25.0、7.3、-50.67和2.76,利用其能增加F1的雄穗分枝数,增高穗位,减小秃尖长,提高封顶性和出籽率;自交系LY0986除穗位高和雄穗分枝数外,其余10个性状的GCA效应值均为正值,其中穗长、穗行数、行粒数和单株产量的GCA效应值分别为15.44、5.04、12.78和21.16,利用其能够提高F1的产量;Mo17和B73两自交系多数性状的GCA效应值为负效应;突变体fu18312仅雄穗分枝数、秃尖长和穗轴粗的GCA效应值为正向效应,其余性状均为负向效应,且株高和穗位高的GCA效应值分别为-10.30和-18.20,表明fu18312具有显著的降秆作用,抗倒伏能力强;突变体fu18321的GCA效应与fu18312类似;突变体fu1603仅穗长、轴粗和秃尖长的GCA为负效应,其余性状均为正效应,利用其能够使F1植株的株高和穗位高增加,穗粗、穗行数、行粒数、单株产量、雄穗分枝数、出籽率增多;突变体fu18398和fu18362的GCA效应与突变体fu1603的GCA效应基本一致;突变体fu17138在单株产量、穗长、行粒数、出籽率和穗位高的GCA效应值分别为15.29、3.48、22.64、4.48和2.84,能够提高F1产量,利用其配制的杂交种丰产性较好。

表2 11个材料11个性状的一般配合力效应值Table 2 Effect values of general combining ability for 11 characters of 11 materials
同一性状不同亲本的GCA值差异较大。如株高和穗位高,突变体fu18312和fu18321的GCA效应值表现为较高的负效应,能够有效降低F1的株高和穗位高,提高杂交组合的抗倒伏性;而突变体fu1603和fu18398表现为较高的正效应。穗长方面,LY0986、fu17138和fu18398的GCA效应值较高,利用其能够增加F1的果穗长度。行粒数方面,LY0986、fu1603、fu18398、fu18362和fu17138的GCA效应值有较高的正效应,利用其能够增加F1的行粒数。单株产量方面,LY0986、fu17138、fu1603和fu18398的GCA效应值较高,在今后的高产育种中可重点加以利用。
2.2.3供试材料11个性状的SCA效应分析 不同组合在某一性状上或某一组合在不同性状间的特殊配合力(SCA)存在很大的差异(表3)。30个组合中,单株产量SCA相对效应值较高的组合包括:LY0986×fu17138、昌7-2×fu18312、LY1621×fu18362、Mo17×fu18321、昌7-2×fu1603、B73×fu18321等。11个亲本中,LY0986、昌7-2、LY1621、fu17138、fu18362、fu18398和fu1603的单株产量GCA效应值较高,利用它们组配杂交组合的SCA效应值也相对较高。穗行数SCA相对效应为正效应、且效应值较高的组合包括LY1621×fu1603、LY1621×fu18362、Mo17×fu18321和LY0986×fu17138;行粒数SCA相对效应为正向效应且效应值较高的组合有LY0986×fu17138、Mo17×fu18321和Mo17×fu1603;亲本中,LY0986、fu18398和fu1603的株高GCA效应值较高,所配组合中以B73×fu1603、LY1621×fu18398和LY0986×fu18398的SCA相对效应值较高;穗位高SCA相对效应值低的包括:LY1621×fu18312、昌7-2×fu18321和LY0986×fu17138,表明突变体fu18312、fu18321和fu17138具有降低株高和穗位高的作用;穗长SCA相对效应值较高的组合包括B73×fu18398、Mo17×fu18362、LY1621×fu18362和Mo17×fu18312,杂交增加了F1穗长;秃尖长SCA相对效应值应越小越好,组合LY0986×fu17138、LY0986×fu18398、B73×fu18362、B73×fu1603和昌7-2×fu18362的秃尖长SCA相对效应较低。根据每一组合株高、穗位高、秃尖长特殊配合力的负向效应和其余各性状特殊配合力的正向效应综合评判,杂交组合LY0986×fu17138、昌7-2×fu18312、LY1621×fu18362、昌7-2×fu1603、B73×fu18321和LY1621×fu18398是比较优良的杂交组合。尤其组合LY0986×fu17138表现最为突出,可进一步扩大试验规模,确定其在品种选育中的应用价值。

表3 30个组合的11个性状特殊配合力效应值Table 3 Effect values of specific combining ability for 11 characters of 30 combinations
2.2.4杂交组合各性状的遗传参数估计 从表4看出,11个性状的配合力方差除雄穗分枝数和秃尖长外,其余9个性状的加性方差均在80%以上,说明F1杂种优势主要由亲本配子的加性效应决定。雄穗分枝数和秃尖长由于亲本相互作用而产生的非加性基因型方差与加性基因型方差相接近,由非加性基因型方差与加性基因型方差共同作用。
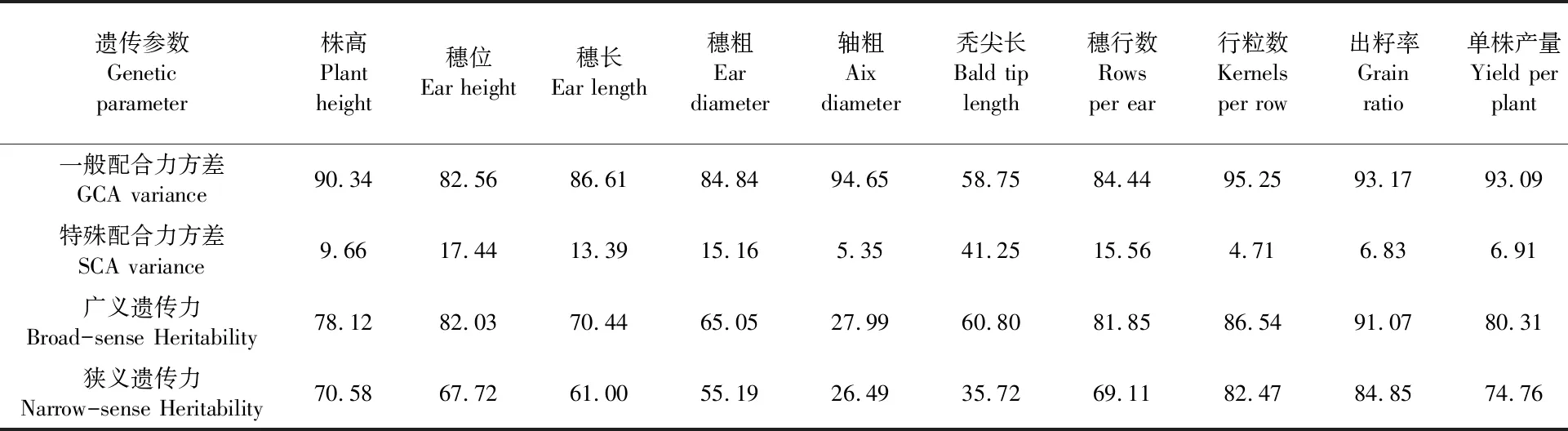
表4 组合各性状遗传参数估计Table 4 Analysis of genetic parameters for all characters of crossed combinations (%)
株高、穗位、穗长、穗行数、行粒数、出籽率和单株产量的广义遗传力和狭义遗传力分别为70.44%~91.07%和61.0%~84.85%,受环境影响较小,能较稳定地遗传给子代,自交系选育时可在早代进行选择而轴粗和雄穗分枝数的广义遗传力和狭义遗传力较低分别27.99% ~ 35.12%和26.49%~18.56%品种选育时不宜在自交系早代进行选择。
3 讨论
3.1 快中子辐射诱变与种质创新、品种改良效应探讨
种质资源的创制是优良品种选育的重要基础。诱发植物突变技术是种质创新与品种改良的有效途径[3]。美国、加拿大等国家的科学家利用突变技术创制了玉米、大豆和大麦低植酸突变体[22];日本科学家利用高能重离子诱变育成了低镉的稻米品种[23];我国科研工作者利用加速器产生的14Me V中子辐照育成水稻新品种中铁31[24],利用离子注入技术选育出小麦新品种皖麦32[25],利用航天诱变技术育成小麦新品种鲁原502[26]。本研究以快中子辐照玉米干种子后,经鉴定筛选出株型紧凑、叶夹角变小突变体fu1603[20-21]、黄绿叶突变体fu17138[19]和低穗位矮秆突变体fu18312,为种质创新、品种改良和基因克隆创造了良好的基础材料,也为植物诱发突变技术提供了良好的理论支持。
3.2 快中子辐射诱变突变体配合力效应探讨
配合力大小是评价作物基础材料利用价值的重要指标,GCA受基因的加性效应控制,能够稳定遗传。前人对辐射诱变材料配合力的影响已有报道[27]。本研究表明,突变体fu18312、fu1603、fu17138、fu18321、fu18398和fu18362不同农艺性状的配合力效应存在差异。就单株产量、穗长、穗粗、穗行数、行粒数和出籽率 的GCA而言,fu18312和fu18321的GCA效应为负效应,fu1603、fu17138、fu18398和fu18362的GCA效应均为正效应,利用这些突变体有利于培育出大穗、多粒、高产的玉米新品种;其余性状如株高、穗位高、秃尖长和分枝数等,突变体的GCA效应值差异较大,需要有选择的利用。不同杂交组合单株产量的SCA表现差异较大。其中,组合(昌7-2×fu18312)和(LY0986×fu17138)的SCA效应为正效应,且效应值较高,可进一步扩大试验规模,确定其在品种选育中应用价值。
3.3 快中子辐射诱变突变体的应用潜力探讨
快中子辐射诱变是为了创制新种质供新品种选育,因此,对新种质进行配合力测定和综合评价具有重要意义。本研究重点分析了6份突变体的GCA和SCA效应,明确了其不同性状配合力特性和利用方向。黄绿叶突变体fu17138的穗部性状GCA效应值较高,利用其组配的杂交组合果穗较长,穗粒数较多,出籽率较高,是高产育种的良好材料,可用于大穗丰产品种的选育;且其株高、轴粗和秃尖长GCA效应表现为较高的负效应,因此,对降低株高、缩小轴粗、减少秃尖长度也有一定的作用。单株产量SCA表明,组合LY0986×fu17138的产量较高,进一步验证了突变体fu17138用于高产育种的潜力。矮秆、低穗位突变体fu18312降秆作用显著,对改良组合抗倒性具有一定优势;但产量性状的GCA效应多为负效应,不利于大穗丰产品种的选育,有待进一步改良。株型紧凑、叶夹角变小突变体fu1603有利于高秆、大穗品种的选育。玉米叶夹角是影响高密度种植的一个重要性状,与株型及产量高度相关,叶夹角减小是目前密植栽培和机械化收获的理想株型特征之一[28],对该材料进行改良和应用,有利于培育出高产、耐密植的玉米新品种。
