尾叶桉不同类型愈伤组织APX类基因转录水平及酶活性的差异1)
2021-06-26黄真池廖春晓赖少娟林文双袁长春
黄真池 廖春晓 赖少娟 林文双 袁长春
(岭南师范学院,湛江,524048)
桉树(Eucalyptus)具有生长快、轮伐期短、适应性强等优点,是我国南方速生丰产林战略树种,在缓解木材等林产品短缺方面发挥着重要的作用[1]。遗传控制、培育优良的抗性品种有利于桉树产业的可持续发展[2]。但是,桉树杂合度高、生长周期长,常规育种困难[3]。通过转基因技术可定向快速改良物种性状。应用转基因技术的前提是携带外源基因的愈伤组织细胞能够启动细胞分裂分化,得到转基因植株[4]。桉树再生的关键是愈伤组织启动分化形成不定芽。因此,研究愈伤分化机理,建立桉树高效再生和遗传转化体系是对其进行遗传改良的基础[5]。
植物细胞中ROS的代谢平衡与其细胞分裂、分化及发育等有重要关系[6]。细胞中的保护酶和抗氧化剂共同调节ROS的平衡,保护酶包括超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、过氧化物酶等,抗氧化剂有维生素C、维生素E、谷胱甘肽等[7]。细胞ROS代谢失去平衡,ROS积累、发生膜质过氧化、引起细胞结构损伤和生物大分子功能丧失。外植体在切伤刺激和离体培养时产生ROS的过度积累,会引起植物细胞膜系统等细胞结构的氧化损伤,是外植体褐化的重要因素之一[8]。
抗坏血酸过氧化物酶(APX)是一种主要的活性氧清除酶,分布于叶绿体、胞浆和过氧化物酶体等细胞器中,以抗坏血酸为电子供体,催化H2O2转化为H2O,在调节植物细胞内ROS代谢平衡中起着重要作用[9]。能正常发育的愈伤组织具有一套高效的抗氧化系统或氧化应激机制来保持自身较强的分化能力[10]。APX酶活性变化及APX基因表达变化可以反映细胞在分化和发育过程中的生理状态[11]。侯大强等[12]发现APX在春石斛兰类原球茎分化芽的过程中存在明显的变化,表明可通过有针对性地在培养基中添加该酶的促进或抑制剂来调控类原球茎的生长或分化。王凤华等[13]采用mRNA差异显示技术在龙眼体胚发生早期发现APX基因的表达与体胚发生有密切联系。郭玉琼等[14]研究表明胞质型APX基因在茶树体胚发生过程中的萌发胚阶段可能发挥着重要作用。过表达APX的转基因拟南芥组织培养过程中外植体细胞积累H2O2减少,促进了愈伤组织诱导和不定芽再生[15]。
目前,有关桉树愈伤分化与APX表达关系的研究鲜有报道。愈伤分化过程中过氧化物酶体、胞浆和叶绿体中APX转录如何变化尚需探讨。本研究分析不同愈伤组织APX酶活性,比较过氧化物酶体、细胞质和叶绿体中APX基因的转录变化,为研究桉树愈伤组织不定芽分化的分子机理提供参考。
1 材料与方法
尾叶桉种子用自来水浸泡2 h,0.2% Triton-100试剂浸泡25 min,并用蒸馏水冲洗后消毒:用体积分数70%酒精消毒30 s,再用体积分数10% NaClO溶液消毒2次,每次7 min,消毒期间用无菌水漂洗,洗净NaClO的残留后,将种子播种到播种培养基(1/2MS+蔗糖20 g/L+琼脂7.5 g/L,pH值5.8~6.0),置于25 ℃暗处萌发。
取8 d苗龄的幼苗,切取0.8~1.0 cm下胚轴作为外植体,接种在添加3.99 mmol/L PBU、0.57 mmol/L 6-BA、0.57 mmol/L IAA和100 mg·L-1Vc的SPCa培养基(679.32 mg·L-1CaCl2、1/4MS铁盐、1/2MS其他无机盐和15 g/L蔗糖)中。(25±2)℃条件下黑暗处理2周后,转移至16 h光/8 h暗光周期、光照度50 μmol/m2·s、温度(25±2)℃的人工气候箱培养12周,统计愈伤组织生长情况。
叶绿素质量分数测定:称取4种类型愈伤组织各0.1 g置于具塞试管中,加入10 mL体积分数95%乙醇于黑暗条件下浸泡24 h。测A665和A649,将测得的值代入公式Cchl(mg/L)=6.63A665+18.08A649,计算叶绿素总质量浓度和叶绿素质量分数。
酶活性测定:称取4种类型愈伤组织各0.2 g(每种样品重复3次),加100 mg PVPP,4 mL酶提取液(50 mmol/L pH值7.0 Tris-HCl缓冲液,1 mmol/L EDTA,5 mmol/L MgCl2,1 mmol/Lβ—巯基乙醇),冰浴匀浆后,将匀浆液于4 ℃,8 000 r/min离心10 min,所得上清即为粗酶液。酶液蛋白质含量采用考马斯亮蓝法测定[16];APX酶活性检测参考文献[11]的方法。
RNA提取及反转录:3种类型愈伤组织各称取100 mg材料于液氮中研磨成细粉,加入1 mL RNAiso Plant Tissue匀浆。按RNAiso Plus试剂说明书步骤提取总RNA,并用Nanodrop 2000c检测总RNA的质量和浓度。反转录按文献[17]的方法进行,得到初始cDNA,备用。所有试剂均是宝生物工程(大连)有限公司产品。
目的基因选择及引物设计:通过检索NCBI数据库和Phytozome 12数据库,寻找已报道的桉树APX基因,并将能在尾叶桉中有效扩增的9个APX基因编号为APX1至APX9。其中编码过氧化物酶体APX(Peroxisomal APX,pAPX)的3个基因编号为APX1—3,编码细胞质APX(cytosolic APX,cAPX)的3个基因编号为APX4—6,编码叶绿体APX(chloroplastic APX,chAPX)的3个基因编号为APX7—9。用primer3plus软件设计qPCR引物。内参基因RTEF和9个APX基因的编号和引物序列见表1。

表1 内参基因和9个APX基因的编号、位置和引物序列
qPCR扩增:qPCR反应仪器为CFX-96Touch(Bio-Rad);25 μL反应体系:SYBR PremixTaqTMⅡ 12.5 μL,10 μmol/L Primer F和Primer R各1 μL,ddH2O 8.5 μL,cDNA模板2 μL。扩增程序:94 ℃,10 s;55 ℃,10 s;72 ℃,15 s。用2-ΔΔCt法[18]计算基因相对表达量。
2 结果与分析
2.1 尾叶桉不同类型愈伤组织特征及APX酶活性变化
下胚轴接种到愈伤诱导培养基中培养8周后,尾叶桉愈伤组织分化形成白色、红色、绿色和带芽点绿色愈伤组织等类型。白色愈伤组织表面较为光滑、质地疏松,红色愈伤组织呈粉红色、质地稍致密,绿色愈伤组织呈绿色,有光泽、细胞团致密,表面有颗粒状突起。绿色愈伤组织表面可分化出绿色芽点、顶芽和幼叶清晰可见,后续培养中芽点伸长长出不定芽(图1)。4种愈伤组织叶绿素质量分数从低到高依次为白色、红色、绿色、带芽点绿色愈伤组织,白色愈伤组织的叶绿素质量分数为0.019 mg/g,带芽点绿色愈伤组织的叶绿素质量分数为0.208 mg/g(表2)。结合愈伤组织颜色差别,可推知绿色愈伤组织的叶绿体已启动发育。12周后,白色和红色愈伤组织开始褐变坏死,而80%以上绿色愈伤组织可分化出不定芽。可见8周前诱导形成绿色愈伤组织是桉树再生的关键。

A.白色愈伤组织;B.红色愈伤组织;C.绿色愈伤组织;D.带芽点绿色愈伤组织;E.不定芽。
白色愈伤组织中APX酶活性最低。从白色愈伤组织分化成红色、绿色或带芽点绿色愈伤组织的过程伴随着APX酶活性的明显升高(表2)。后续培养中APX酶活性最高的红色愈伤组织不能形成不定芽,APX酶活性稍低的绿色愈伤组织有80%以上可分化出不定芽。推测愈伤组织的分化除了受APX酶活性的影响,还与APX酶在细胞中的定位有关。
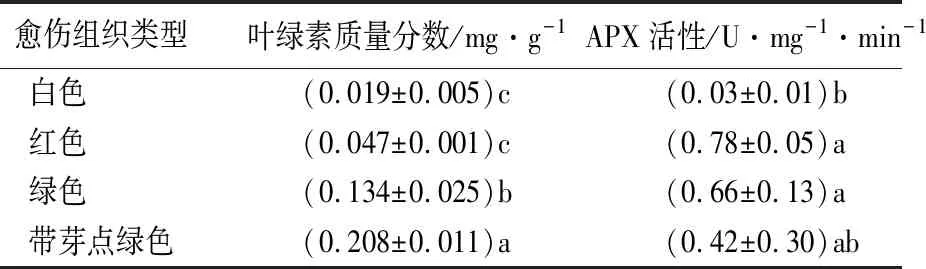
表2 尾叶桉不同类型愈伤组织APX酶活性
2.2 尾叶桉不同类型愈伤组织9个APX基因转录水平变化
2.2.1过氧化物酶体中3个APX基因转录水平变化
设红色愈伤组织基因的相对表达量为1,3种不同类型愈伤组织的基因表达量存在显著差异(表3)。与红色愈伤组织相比,绿色愈伤组织和带芽点绿色愈伤组织中位于过氧化物酶体的3个APX基因的表达量均上调,且带芽点绿色愈伤组织APX相对表达量显著高于绿色愈伤组织。过氧化物酶体中3个APX基因表达上调促进了不定芽的分化。

表3 尾叶桉不同类型愈伤组织过氧化物酶体中3个APX基因转录水平变化
2.2.2细胞质中3个APX基因转录水平变化
3种愈伤组织胞质APX4基因转录变化不明显。与红色愈伤组织相比,绿色愈伤组织和带芽点绿色愈伤组织胞质APX5和APX6基因表达量均上升(表4)。总体来看,胞质APX表达增强有利于不定芽芽点的分化。但胞质APX同工酶基因的变化不尽相同。

表4 尾叶桉不同类型愈伤组织细胞质中3个APX基因转录水平变化
2.2.3叶绿体中3个APX基因转录水平变化
3种不同类型愈伤组织叶绿体3个APX基因表达量均存在显著性差异(表5)。绿色愈伤组织和带芽点绿色愈伤组织的APX7和APX8的基因表达量显著高于红色愈伤组织。两者APX8的相对表达量分别为红色愈伤组织的21.0和29.4倍。APX7和APX8表达上调促进了绿色愈伤组织分化和芽点形成。APX8定位于叶绿体,相对表达量剧增与绿色愈伤组织和带芽点绿色愈伤组织中叶绿体的发育和数量倍增有关。绿色愈伤组织和带芽点绿色愈伤组织的APX9的表达下调,说明同一细胞器中的APX同工酶的功能不同或影响愈伤分化的作用不同。
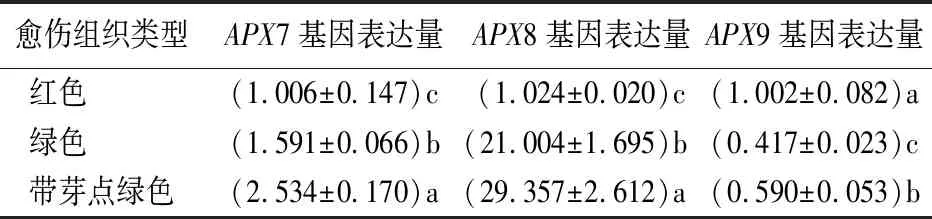
表5 尾叶桉不同类型愈伤组织叶绿体中3个APX基因转录水平变化
3 结论与讨论
根据亚细胞定位,高等植物APX分为4个亚家族,分别是细胞质APX、线粒体APX、叶绿体APX和过氧化物酶体APX[19-20]。在过氧化物酶体中,ROS是光呼吸、脂肪酸β氧化和酰脲代谢等的副产物。虽然CAT能够清除高浓度的H2O2,但其与H2O2亲和力低。相反,APX与H2O2亲和力高,可显著降低H2O2的浓度,防止H2O2从过氧化物酶体中泄漏出去[21]。pAPX的N端活性结构域面向胞浆,C端结构域起着锚定作用并促进酶功能的实现[22]。过表达pAPX可增强花生的耐盐性[23]。目前未见pAPX表达与愈伤组织分化相关的报道。试验中,3个过氧化物酶体APX基因(APX1、APX2、APX3)的相对表达量均为带芽点绿色愈伤组织最高。表明定位于过氧化物酶体的APX类基因表达量上调能提高pAPX同工酶活性,清除愈伤组织过量的ROS,减轻ROS对细胞的毒害,有利于绿色愈伤组织形成和芽点的分化。
胞质APX可清除H2O2,减少H2O2向叶绿体流入。当缺乏细胞质APX时,将导致叶绿体H2O2清除系统崩溃、H2O2浓度升高、蛋白质被氧化[24]。与下胚轴相比,2周龄尾叶桉愈伤组织胞质APX5和APX6基因的转录下调,而胞质APX4基因的转录则显著上调[11]。本试验中,具有芽分化能力的绿色和带芽点绿色愈伤组织胞质APX5和APX6基因转录显著上调。不同愈伤组织间胞质APX4基因表达变化不明显。说明细胞质APX基因转录上调有助于保护叶绿体H2O2清除系统,促进愈伤组织叶绿体的发育。
叶绿体作为光合作用的场所与其他细胞器相比更易遭受氧化胁迫的伤害[25]。由于叶绿体中无CAT,APX在叶绿体中起着关键作用[9]。叶绿体有两种APX同工酶,即类囊体结合型和基质结合型APX,在光氧化胁迫下的光保护和基因表达调控方面发挥双重作用[26]。实验发现带芽点绿色愈伤组织叶绿体APX7、APX8的表达量显著高于其它类型愈伤组织。APX8在绿色愈伤组织和带芽点绿色愈伤组织中显著上调,分别为红色愈伤组织的21.0倍和29.4倍。推测叶绿体中APX8的显著上调与愈伤组织叶绿体发育、叶绿体数量倍增和芽原基形成密切相关。
综上所述,尾叶桉愈伤组织在切伤刺激和离体培养时,通过提高APX酶活性以降低活性氧伤害。过氧化物酶体、细胞质和叶绿体中的APX基因总体表达上调有利于清除体内过量的ROS,促进芽原基的分化和不定芽的形成。绿色愈伤和带芽点绿色愈伤中叶绿体APX8转录水平显著升高,说明APX8表达与叶绿体发育密切相关。本研究可为提高尾叶桉不定芽诱导率积累资料,为深入研究桉树愈伤组织分化的分子机理提供参考。
