河南省花生白绢病菌群体多样性及对萎锈灵敏感性研究
2021-06-25张忠信杜鹏强董文召
李 敏,李 爽,张忠信,杜鹏强,周 琳,董文召
(1.河南农业大学 植物保护学院/河南省新型农药创制与应用重点实验室/河南省绿色农药创制工程技术研究中心,河南 郑州 450002;2.河南省作物分子育种研究院/河南省农业科学院 经济作物研究所,河南 郑州 450002)
花生白绢病是由齐整小核菌(SclerotiumrolfsiiSacc.)引起的土传真菌病害,亦称茎腐病、脚腐病、南方枯萎病等。该病原菌以菌核和菌丝在土壤中或植株病残体上越冬,在适宜的条件下,菌丝或萌发的菌核从花生植株茎基部侵入,导致受侵染植株变黄、枯萎,根茎部变褐、腐烂[1-3];亦可侵染花生荚果和果仁,影响产量。花生白绢病在世界各花生产区均有发生[1]。近年来,随着我国花生种植面积的增加,连作重茬及秸秆还田等耕作方式日益普遍,导致花生白绢病在我国各花生产区发生危害日趋严重、分布区域逐年扩大,山东、河南和湖北等省[4-10]均有大面积暴发流行的报道,地块发病株率为10%~30%,严重地块达40%以上;发病田块减产10%~20%,病重田块减产50%以上,甚至造成绝收,该病害已成为限制我国花生生产的一个重要因子。
控制花生白绢病菌侵染将是花生生产中持续面对的问题。虽然作物轮作是病害治理的重要基础,但由于白绢病菌有500多种寄主植物以及菌核在土壤中可存活3~5 a,极大地限制了采用与非寄主作物轮作或整地时用土深埋受侵染作物残体等物理和栽培措施[11]。再者,持久的寄主抗性可能是最有效的病害管理手段之一,但目前市面上缺乏抗白绢病的花生品种。尽管有报道称美国花生田白绢病菌种群对五氯硝基苯、氟酰胺和戊唑醇的敏感性下降[12-13],但施用化学杀菌剂仍是目前防治花生白绢病最常用的措施。
利用化学防治和生物防治控制花生白绢病菌,必须了解该病原菌的分布和多样性。菌丝亲和群(MCG)是一种广泛用于研究病原真菌的遗传多样性、地理分布、病害流行规律和病原致病性变异等的重要手段[14-15]。NALIM等[16]和XIE等[17]分别通过MCG分析对美国德克萨斯州和美国南部6州花生白绢病菌田间种群进行了多样性评估;OKABE等[18]亦采用MCG分析了日本茨城县花生白绢病菌田间种群的多样性;CILLIERS等[19]对非洲南部15个地区7种寄主上分离的121株白绢病菌进行MCG分析的结果表明,白绢病菌在MCG、寄主和资源分布上存在巨大的变异和联系。LE等[11]结合MCG和核糖体DNA-ITS序列分析,证实了分离自越南中部花生、番茄和芋头上的103株白绢病菌菌株的菌丝相容性、生长速率和菌核特性等遗传和表型性状上表现出多样性。作为花生种植面积和总产量均居全国前列的河南省,近年来花生白绢病严重威胁着省内南阳、驻马店、商丘、周口、新乡等主产区的花生生产。但目前关于河南省花生白绢病菌群体多样性及分布尚缺乏系统研究。截至2020年12月21日,我国登记用于防治花生白绢病的化学杀菌剂仅有7种(http://www.chinapesticide.gov.cn /),其中萎锈灵(Carboxin)是一种内吸性较强的琥珀酸脱氢酶抑制剂类杀菌剂(SDHIs),国外亦早已登记用于防治由齐整小核菌引起的多种植物白绢病[2,20-21]。由于其作用靶标单一,有关萎锈灵及结构相似化合物的抗药性在多种致病真菌,如灰霉病菌(Botrytiscinerea)、瓜类白粉病菌(Podosphaeraxanthii)和西瓜蔓枯病菌(Didymellabryoniae)[22-24]等上已有报道。但花生白绢病菌对萎锈灵的敏感性变化在我国尚未见研究报道。因此,在对河南省5个花生主产区花生白绢病系统调查的基础上,对采集的28株代表性菌株进行形态学特征研究和MCG分析,测定萎锈灵对28株菌株的毒力,旨在明确河南省主要花生产区白绢病菌群体的多样性、分布及对萎锈灵的敏感性,为科学制定和成功实施花生白绢病的生物防控和化学防控策略提供依据。
1 材料和方法
1.1 试验材料
培养基为PDA培养基:马铃薯200 g、葡萄糖20 g、琼脂20 g,蒸馏水定容至1 L。
试剂与仪器:95%萎锈灵(Carboxin),广西田园生化有限公司生产;二甲基亚砜(分析纯),北京索莱宝科技有限公司生产;吐温-80(分析纯),北京索莱宝科技有限公司生产;RXZ型智能人工气候箱,宁波江南仪器厂生产;GXZ型智能光照培养箱,宁波江南仪器厂生产;SW-CJ-2FD洁净工作台,苏净集团苏州安泰空气技术有限公司生产;ME104E/02电子天平,梅特勒-托利多仪器(上海)有限公司生产;游标卡尺,香港杭泰(国际)集团有限公司生产。
1.2 试验方法
1.2.1 田间发病率调查和菌株采集 2019年7—8月,在河南省南阳、驻马店、商丘、郑州和新乡5个花生主产区,随机选取23块花生田对花生白绢病的发病率进行调查。每个田块之间至少相隔10 km上。每个田块采用五点取样法进行调查,每点调查50株,记录病株数,并计算发病率。发病率=病株数/调查总株数×100%。在23个田块中,每个病株上的菌核视为1份样本,每个田块均采集15份样本,并将其分别放于塑料袋中,带回实验室进行菌株分离。
1.2.2 菌株的分离和保存 将每份菌核样本先用30%次氯酸钠溶液浸泡30 s,再用75%乙醇浸泡30 s,之后用无菌水冲洗3遍,最后用无菌滤纸吸干菌核表面的水分后转接到PDA平板上,置于30 ℃培养箱中黑暗培养。2 d后挑取单菌核萌发的单菌丝进行纯化培养,此过程重复3次。收集已纯化菌株产生的菌核,一份置于灭菌的2 mL 离心管中于4 ℃冰箱中保存备用;另一份置于灭菌的20%甘油管中于-80 ℃长期保存。
1.2.3 形态学特征观察 将已活化的28株花生白绢病菌菌株分别置于PDA平板上,于30 ℃培养箱中黑暗培养,2 d后,在菌落边缘打取直径为6 mm的菌饼,转接于另一PDA平板(直径90 mm)中央,在25 ℃培养箱中培养2 d后,用十字交叉法测量菌株的菌落直径,并计算每株菌株的菌丝生长速率。菌落直径测量后,各PDA平板继续置于相同条件下培养,期间观察并记录菌落特征。28 d后,记录每个培养皿中的菌核数量,收集每个PDA平板中的所有菌核,置于30 ℃培养箱中风干24 h后,随机抽取20个成熟菌核,用游标卡尺测量菌核直径;用电子天平称量每个平板中所有菌核的总干质量,并根据每个平板中菌核的数量计算单个菌核的干质量。每株菌株设置3个重复。
1.2.4 MCG测定及其多样性分析 MCG测定参照XIE等[17]的方法,略作修改:将所有菌株的菌核置于PDA平板上,于30 ℃培养箱中黑暗活化,2~3 d后,在菌落边缘打取直径为6 mm的菌饼,转接到PDA平板(直径90 mm)中。每个PDA平板中包含2株菌株共3个菌饼,其中,1株菌株转接2个菌饼作为亲和对照,另1株菌株转接1个菌饼,菌饼之间距离为4~5 cm。转接后的PDA平板放置于25 ℃培养箱中黑暗培养,14 d后观察并记录各PDA平板上菌株间菌丝交汇处的反应情况。当2株配对菌株的菌丝交汇处相互融合,则记为亲和,并被划分为相同的亲和群;当2株配对菌株的菌丝交汇处产生1条明显的坏死带或空白带时,则记为不亲和,并被划分为不同的亲和群。本试验中,28株花生白绢病菌菌株在所有可能的情况下两两组合进行配对,每个配对重复3次。
花生白绢病菌MCG的多样性水平高低以香农-威纳多样性指数(H)来表示,香农-威纳多样性指数是考虑到个体数量及种类数量的一个多样性指数,其计算公式为:H=-∑[pi×lnpi],pi为第i个MCG类群中各菌株所占的频率,香农-威纳多样性指数的数值越大,代表多样性水平越高[25]。
1.2.5 菌株对萎锈灵的敏感性测定 采用菌丝生长速率抑制法[26]测定花生白绢病菌对萎锈灵的敏感性。称取一定量的95%萎锈灵原药溶于少量二甲亚砜中,再用含2%吐温-80的无菌水配制成100 mg/L的母液,并稀释成系列浓度的药液,使其中的二甲亚砜和吐温-80的终浓度分别控制在6%和2%。分别取1 mL药液和9 mL PDA培养基于直径9 cm的培养皿中混匀,制成终质量浓度分别为1、0.5、0.25、0.125、0.062 5和0.031 25 mg/L的含药PDA平板。将各供试菌株的菌核置于PDA平板上于30 ℃培养箱中黑暗活化,2~3 d后,在菌落边缘打取直径为6 mm的菌饼,并转接于含有萎锈灵系列质量浓度的PDA平板上,以含0.6%二甲亚砜和0.2%吐温-80的无药PDA平板为相应的溶剂对照,每处理3次重复。置于30 ℃培养箱中黑暗培养,3 d后,用十字交叉法测量菌落直径,计算菌丝生长抑制率,并根据概率值分析法求得毒力回归方程和有效中浓度(EC50值)。菌丝生长抑制率=[(对照菌落直径-菌饼直径)-(处理菌落直径-菌饼直径)]/(对照菌落直径-菌饼直径)×100%。
1.3 数据分析
试验数据采用Excel 2010和SPSS 22.0软件进行统计分析,应用Duncan’s新复极差法进行差异显著性检验,利用Pearson相关系数法进行相关性分析。
2 结果与分析
2.1 田间发病率和代表性菌株
通过对5个花生主产区田间随机调查(图1),发现被调查的23个田块中花生白绢病均有发生。其中,2个田块(5和18)的发病率分别高达50.4%和47.2%;10个田块的发病率为10.4%~28.4%;其余11个田块的发病率相对较低,介于3.6%~8.8%。此外,还发现13个春播花生田块中白绢病的发病率均高于10个夏播花生地块。从河南省5个花生主产区采集的345份样本中成功分离到253株花生白绢病菌株,并分别从5个花生主产区随机选取其中的28株作为代表性菌株进行MCG分析和形态学特征研究(表1)。

表1 河南省23个采样田块28株花生白绢病菌株信息Tab.1 Information on the 28 isolates of Sclerotium rolfsii from different peanut fields in Henan Province
2.2 形态学特征
2.2.1 菌落特征 对28株花生白绢病菌菌株的菌落特征观察结果表明,所有的菌株均产生白色的菌丝和形成褐色或深褐色的菌核,但这些菌株的菌核在PDA平板上的分布存在差异,其中,8株菌株产生的菌核分布在PDA平板的边缘(图2a),其余20株菌株产生的菌核则随机分布在PDA平板上(图2b)。
2.2.2 菌丝生长速率 从表2可以看出,花生白绢病菌不同菌株的菌丝生长速率存在差异。28株菌株的菌丝生长速率在0.55~1.31 mm/h,其中采自南阳的菌株NYXYWJ-4菌丝生长速率最小(0.55 mm/h),而采自新乡的菌株XXYYJZ-3菌丝生长速率最大(1.31 mm/h)。对于不同采样地的菌株来说(因郑州和驻马店均只有2株菌株,故未做比较),南阳、商丘和新乡的菌株在菌丝生长速率上没有显著差异(表3)。
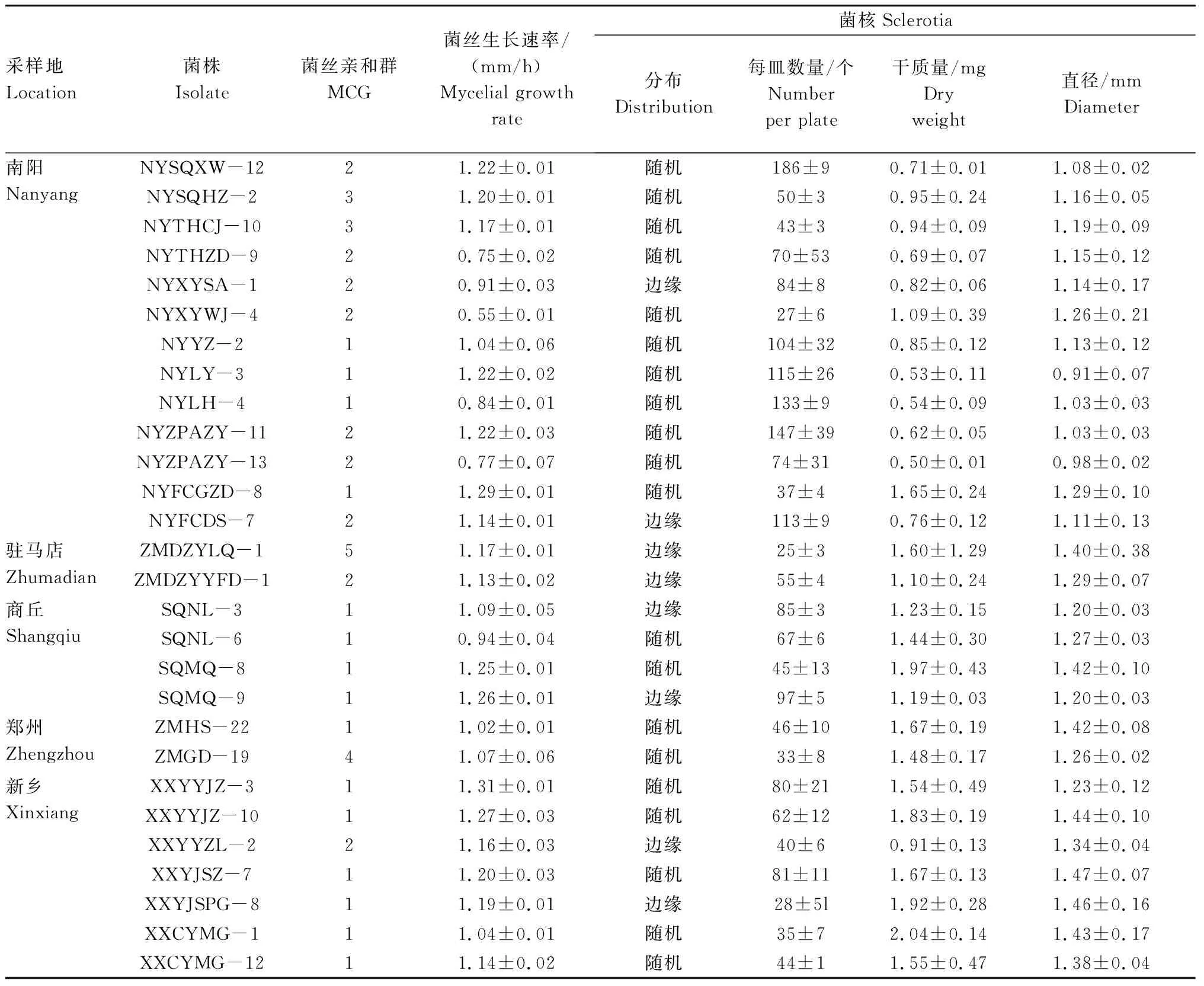
表2 不同采样地的28株花生白绢病菌菌株的表型特征
2.2.3 菌核特征 在离体条件下,28个花生白绢病菌菌株的菌核干质量、菌核直径和菌核数量均存在差异,菌核干质量为0.50~2.04 mg,菌核直径为0.91~1.47 mm,每皿菌核数量为25~186个(表2)。分析不同采样地的菌株可知(因郑州和驻马店均只有2个菌株,故未做比较),南阳、商丘和新乡的菌株在菌核数量上不存在显著差异;新乡和商丘的菌株菌核干质量明显比南阳的大;南阳、新乡和商丘菌株的菌核直径存在显著差异(表3)。

表3 河南省不同采样地花生白绢病菌的形态学特征Tab.3 Morphological characteristics of Sclerotium rolfsii from different locations in Henan Province
2.3 菌株的菌丝生长速率、菌核数量、菌核干质量、菌核直径的相关性分析
对28株菌株的菌丝生长速率、菌核数量、菌核干质量、菌核直径进行Pearson相关性分析的结果(表4)表明,菌丝生长速率与菌核数量(r=0.097,P>0.01)没有相关性,而菌丝生长速率与菌核干质量(r=0.350,P>0.01)、菌核直径(r=0.219,P>0.01)呈正相关,但没有达到显著水平;菌核数量与菌核干质量(r=-0.623,P<0.01)、菌核直径(r=-0.648,P<0.01)呈显著负相关;菌核干质量与菌核直径(r=0.937,P<0.01)呈显著正相关。

表4 28株花生白绢病菌的菌丝生长速率、菌核数量、菌核干质量及菌核直径的相关性分析Tab.4 Correlation analysis of mycelial growth rate,number,dry weight,diameter of sclerotia among 28 Sclerotium rolfsii isolates from peanut
2.4 菌株的MCG及其多样性分析
根据2株配对菌株间菌丝交汇处的反应,28株花生白绢病菌菌株共划分为5个MCG(表5),并随机命名为MCG1、MCG2、MCG3、MCG4、MCG5。5个MCG的频率和地理分布存在差异。MCG1是最大的亲和群,包含15株菌株,占总菌株数的53.57%,分布于除驻马店外的所有采样地;MCG2的菌株数量仅次于MCG1,包含9株菌株,占总菌株数的32.14%,分布于南阳、新乡和驻马店3个采样地;其余3个MCG占总菌株数的14.29%,其中MCG3、MCG4、MCG5分别包含2、1、1株菌株,其分布均局限于1个采样地,分别为南阳、郑州和驻马店。在这5个MCG中,仅来自商丘的菌株被划分到1个亲和群中,而其他采样地的菌株分别被划分到不同的亲和群中,说明除商丘外,其他采样地出现的MCG与菌株的地理来源无相关性。

表5 28株花生白绢病菌菌株的MCG及其多样性指数Tab.5 Mycelial compatibility groups and MCGs-related diversity index of 28 Sclerotium rolfsii isolates from peanut
菌株的MCG多样性分析结果表明(表5),28株花生白绢病菌菌株的香农-威纳多样性指数(H)为1.13。其中,来自商丘的菌株H为0,来自南阳的菌株H为0.98,来自新乡的菌株H为0.41,来自郑州和驻马店的H均为0.69,说明这5个采样地花生白绢病菌MCG的多样性水平不一致。
2.5 28株菌株对萎锈灵的敏感性
采用菌丝生长速率抑制法测定了28株花生白绢病菌株对萎锈灵的敏感性(表 6),其中最大值(0.299 0 mg/L)是最小值(0.114 4 mg/L)的2.61倍,平均EC50值为0.182 8 mg/L。上述结果表明,28株花生白绢病菌株对萎锈灵均表现为敏感,且不同菌株对萎锈灵的敏感性存在一定的差异。

表6 28株花生白绢病菌菌株对萎锈灵的敏感性Tab.6 Sensitivity of 28 Sclerotium rolfsii isolates from peanut to carboxin

续表6 28株花生白绢病菌菌株对萎锈灵的敏感性Tab.6(Continued) Sensitivity of 28 Sclerotium rolfsii isolates from peanut to carboxin
分析不同采样地的花生白绢病菌对萎锈灵的敏感性可知,不同地区菌株的EC50平均值间存在差异,但未达到显著水平(表7)。其中,郑州的菌株最敏感,EC50平均值为0.144 7 mg/L,而驻马店的菌株最不敏感,EC50平均值为0.206 7 mg/L,后者EC50平均值是前者的1.43倍。

表7 河南省不同采样地花生白绢病菌菌株对萎锈灵的敏感性比较Tab.7 Comparison of sensitivity of Sclerotium rolfsii to carboxin from different locations in Henan Province
从敏感性频率分布图(图3)可以看出,供试花生白绢病菌对萎锈灵的敏感性分布呈单侧峰曲线分布,说明供试菌株中没有出现敏感性下降的群体。
3 结论与讨论
齐整小核菌作为花生白绢病的致病因子,了解花生田中齐整小核菌群体多样性和流行病学的基本知识,有助于制定和采取有效的防治措施对花生白绢病进行可持续性治理。本课题组(河南农业大学植物保护学院花生病虫害课题组)于2019年对河南省5个花生主产区23个花生田块的实地调查显示,花生白绢病的发病率为3.6%~50.4%,与我国其他多个省份的报道基本一致[5-8]。调查还发现,夏播花生田块白绢病的发病率均低于春播花生田块,且前茬作物为小麦和大蒜的田块发病率较低。ZEIDEN等[27]和MINTON等[28]也报道了花生与大蒜、小麦轮作可降低花生白绢病的发病率。但也有报道齐整小核菌可侵染小麦,引起麦苗猝倒、腐烂,不能作为花生轮作作物[29]。
本研究发现,河南省不同采样地的28株花生白绢病菌株在菌落特征、菌丝生长速率及菌核的数量、干质量和直径等形态学特征上存在着一定差异,说明河南省5个花生主产区的花生白绢病菌菌株具有多样性,这与宋万朵等[30]对我国12个省市的39株花生白绢病菌株形态学研究结果一致。LE等[11]研究发现,越南偏远地区的3个花生白绢病菌菌株产生的菌核显著大于其他地区菌株产生的菌核,本研究也发现,新乡菌株的菌核显著大于其他地区菌株的菌核,推测花生白绢病菌的菌核大小可能与地理位置有关。此外,本研究对28株菌株的菌丝生长速率、菌核数量、菌核干质量和菌核直径两两进行相关性分析,发现菌株的菌核数量、干质量和直径之间存在显著相关性,证实和扩展了前人对齐整小核菌形态学特征中各测量值间关系的研究结果[7,31]。
早期关于白绢病菌MCG分析的研究表明,在一个特定的寄主或在一个有限的地理区域内白绢病菌群体能够被划分为不同的MCG[17,31-32];来自同一MCG的菌株在遗传上比来自不同亲和群的菌株更为相似[19,33-34]。本研究中,采自河南省5个花生主产区的28株花生白绢病菌株被划分为5个MCG,进一步表明这28株菌株间存在多样性。其中,最大的MCG1中菌株分布于除驻马店外的其他4个采样地,表明MCG1中的菌株可能通过种子调运、雨水、昆虫和农事操作等方式在河南省多数花生产区已广泛传播,REMESAL等[32]对甜菜白绢病菌MCG的研究也得出了类似的结论。宋万朵等[30]研究发现,驻马店的7株花生白绢病菌株被划分为6个亲和群,而本研究中采自驻马店的2株菌株被划分为2个亲和群,推测该区花生白绢病菌群体多样性水平较高。值得注意的是,本研究中还发现MCG与菌株的地理位置似乎没有相关性,与前人的研究结果一致[19,30]。基于MCG的测定结果,香农-威纳多样性指数(H)被广泛用于测定病原菌群体多样性水平的高低[25,31,35]。本研究中28株花生白绢病菌株总的香农-威纳多样性指数(H)为1.13,与前人的研究结果相比[31-32,35],其多样性水平相对较低。
本研究结果表明,采自河南省5个花生主产区的28株白绢病菌株对SDHIs杀菌剂萎锈灵均很敏感,且不同菌株间敏感性无显著差异,28株供试菌株对萎锈灵的敏感性呈单侧峰曲线分布,未检测到有敏感性明显分化的群体存在,该结果能反映出田间自然情况下病原菌对SDHIs类杀菌剂的敏感性特征。但由于SDHIs杀菌剂靶标的作用位点单一,已被杀菌剂抗性行动委员会(Fungicides Resistance Action Committee)归为中等至高抗性风险[36],且FRANKE等[12]报道了美国乔治亚州花生白绢病菌对SDHIs类杀菌剂氟酰胺已产生抗药性,因此,有必要进一步加强河南省花生白绢病菌对萎锈灵的抗性监测、风险评估及治理措施研究。
