巨龙竹林分结构及物种多样性特征1)
2021-06-25郭强官凤英
郭强 官凤英
(国际竹藤中心,北京,100102)
刘蔚漪 辉朝茂
(西南林业大学)
邹学明
(沧源县林业和草原局)
林分结构特征及其物种多样性是林分在发展过程如生长更新、自然稀疏、经历干扰活动等的综合反映,很大程度决定了林分的生长稳定性、发展可能性[1-3]。目前,以毛竹(Phyllostachysedulis)为代表的散生竹林在结构、物种多样性等方面已有较多报道,涉及年龄结构、直径结构、密度结构、生物量、生产力预测等多个领域、层次[1,4-6],形成了较为系统且深入的研究体系。但有关丛生竹类的研究较少[7-9]。巨龙竹(Dendrocalamussinicus)为禾本科竹亚科牡竹属特大型合轴丛生竹种,有“竹中之王”的美誉[10-12]。该竹种也是我国西南亚热带、热带地区极为重要的竹子资源,其生态功能价值、社会文化价值、经济价值潜力巨大[13-14]。但由于巨龙竹分布偏远且零散、分布山区地形与气候复杂、竹丛形态巨大等综合原因,巨龙竹林分结构、物种多样性等方面的研究还较为空白,未见有关报道。
本研究在对滇西南巨龙竹林分结构因子连续调查的基础上,探讨了近自然生长的巨龙竹林分结构、变化特征,并利用群落学样方调查法,分析了巨龙竹林分植被组成、物种多样性、林下植被种-面积关系。旨在认识、了解巨龙竹林分结构特征、植被群落发展水平,为滇西南巨龙竹植被群落的保护、发展提供参考。
1 研究区概况
研究区位于云南省临沧市沧源佤族自治县(23°4′~23°40′N,98°52′~99°43′E),属亚热带季风气候,干湿季分明,一般春、冬季为旱季,夏、秋季为雨季;年平均气温约21 ℃,年积温约7 500 ℃,年降水量1 900~2 200 mm,月均空气相对湿度约为80%;土壤类型为红壤、赤红壤、砖红壤等。
2 研究方法
2.1 样地设置
在研究区内,通过预调查筛选出具有一定数量规模且连续分布的巨龙竹林分为研究对象,并将其所在的整个林地设为固定试验样地。试验样地的巨龙竹在近7 a内无任何人为经营管理措施,属于近自然生长状态。巨龙竹试验样地总面积为1.83 hm2,海拔1140~1160 m,坡度为7°~33°,坡向为西向,建群种面积为1.80 hm2,干扰树面积为0.03 hm2,林分郁闭度为0.35~0.65。
2.2 样地调查
2017—2019年,每年的12月对巨龙竹竹丛数、丛立竹干数、胸径、年龄等林分结构因子进行调查,并记录期间的枯死竹。2019年12月,分别从东西、南北2个方向测定其竹丛冠幅、竹蔸直径。通过分布参数值分析巨龙竹林分年龄、直径、密度等结构特征[15-17];利用数学模型表征其结构因子分布规律[18-20];通过相关性分析探讨各林分结构因子间的相关性。
2019年11月,在林分内沿“S”形均匀测设39个5 m×5 m的灌木样方,并在各灌木样方内以五点取样法测设5个1 m×1 m的草本样方,共计39个灌木样方、195个草本样方。同时对各样方的植物种类、植株数量、高度、盖度等因子进行调查。利用重要值(Pi)反映各林层植被物种的生存优势(Pi>30%,为优势种);利用物种丰富度(M)、Shannon-Wiener多样性指数(H)、Simpson多样性指数(D)、Pielou均匀度指数(JH)4个指标反映各林层物种多样性[21-22]。具体公式如下:
物种重要值=相对频度+相对高度+相对盖度。
(1)
M=S。
(2)
(3)
(4)
(5)
式中:S为植被物种种数;Pi为第i个物种的重要值。
2019年11月,在林分内均匀测设3个20 m×20 m的林下植被大样方;在各样方内沿边界划分为0.5 m×0.5 m、1.0 m×1.0 m、1.5 m×1.5 m、…、19.5 m×19.5 m、20.0 m×20.0 m的变化样方,并分别记录对应变化样方中的灌木或草本植物物种数。通过种-面积曲线研究巨龙竹林下植被物种数与调查样方面积间的变化关系,探讨可以涵盖巨龙竹林下大部分植物种类的最小调查面积。
2.3 数据处理
利用Office Excel 2007对外业调查所取得数据进行整理,制成相关图表;利用SPSS 19.0进行数据方差分析、显著性检验;利用ForStat 2.0进行分布模型拟合、检验。
3 结果与分析
3.1 林分结构与变化特征
2017—2019年的巨龙竹林分结构及变化特征如图1。分析可知,2019年巨龙竹干龄分布呈右偏缓峰状(峰度为-0.88,偏度为0.16),以1~4年生的竹干居多,其累积数量比例达80.88%,平均干龄为3.10 a;竹丛年龄分布呈左偏聚集状(峰度为1.54,偏度为-0.76),以2.5~3.5年生的竹丛居多,其累积数量比例达73.15%,平均竹丛年龄为3.13 a;直径分布呈左偏聚集状(峰度为0.57,偏度为-0.59),以18、20 cm径阶的竹干居多,其累积数量比例达53.38%,平均直径为17.95 cm;丛立竹密度分布呈右偏缓峰状(峰度为-0.39,偏度为0.69),以10~30干/丛的竹丛数居多,其累积数量比例达51.85%,平均丛立竹密度为25.57干/丛。
除2018年与2019年的直径分布差异不显著(P>0.05),其余结构因子在各生长季间分布的差异均极显著(P<0.01,图1)。除2019年的1~2年生巨龙竹数量较2018年有所减少,其余干龄的巨龙竹数量均逐年增多,且数量差距逐渐减小,分布趋于缓和。因此,竹丛年龄分布向右偏移并集中在更高的年龄范围;直径分布逐年向上且微右偏移,集中在更大的直径范围,2019年出现特大径级竹(26 cm);丛立竹密度分布逐年向右偏移,其分布范围扩大,且多数竹丛集中在较高的密度范围。
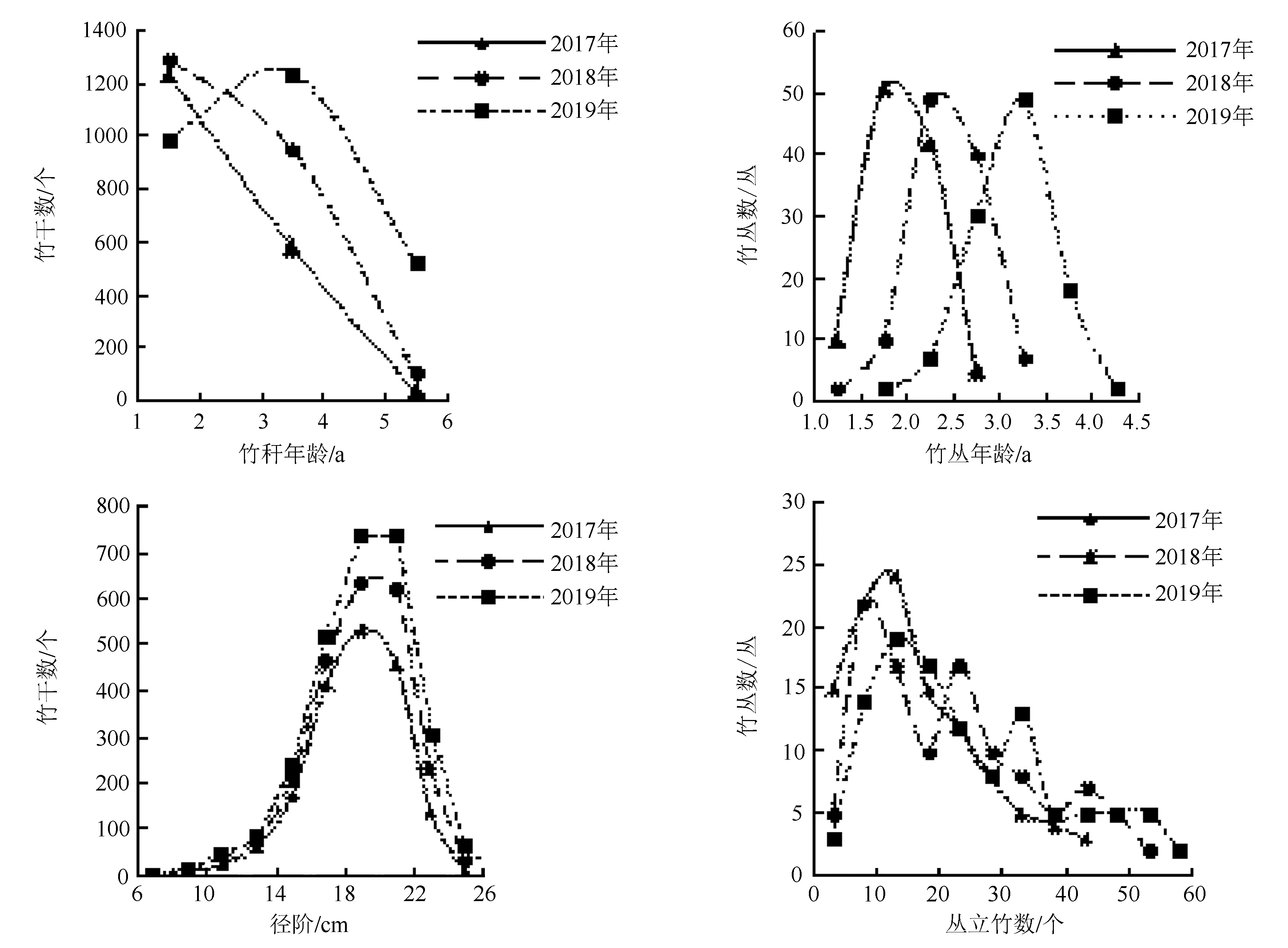
图1 2017—2019年巨龙竹林分结构及变化特征
3.2 直径与密度分布拟合
将2017—2019年的巨龙竹林分直径、丛立竹密度分布标准化(开平方根)后,利用相关模型对其进行分布拟合、卡方检验(表1,表2)。分析可知,在各生长季均以威布尔拟合其分布的卡方值(χ2)最小、概率值(p,p>χ2)最大,表明其分布的实际值与期望值差别最小、拟合效果最优。
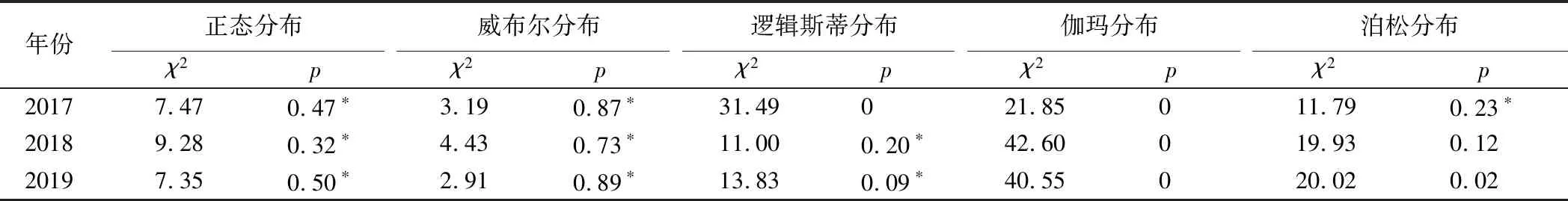
表1 2017—2019年生长季的巨龙竹林分直径分布拟合与卡方检验

表2 2017—2019年生长季的巨龙竹丛立竹密度分布拟合与卡方检验
3.3 林分结构因子相关性
巨龙竹林分结构因子相关性分析(表3)结果表明,竹冠投影面积与竹蔸面积、丛立竹干数均呈极显著高度正相关;与平均胸径呈显著正相关;与平均年龄的相关性较弱且未达显著水平。竹蔸面积与平均胸径、年龄均呈显著低度正相关。丛立竹干数与平均胸径、年龄均呈显著低度正相关。竹丛立竹干数越多,竹蔸面积越大,其竹冠投影面积也越大。竹干平均胸径、年龄仅表征个体的平均生长状态,属于间接影响因子,与竹冠投影面积的相关性较弱[23]。

表3 巨龙竹林分结构因子相关性
巨龙竹竹冠投影面积为25~85 m2/丛,竹蔸面积为8~30 m2/丛,其累积数量比例分别为63.46%、54.81%(图2)。根据巨龙竹竹冠投影面积与竹蔸面积、丛立竹干数间的相关性,可建立通用线性方程:
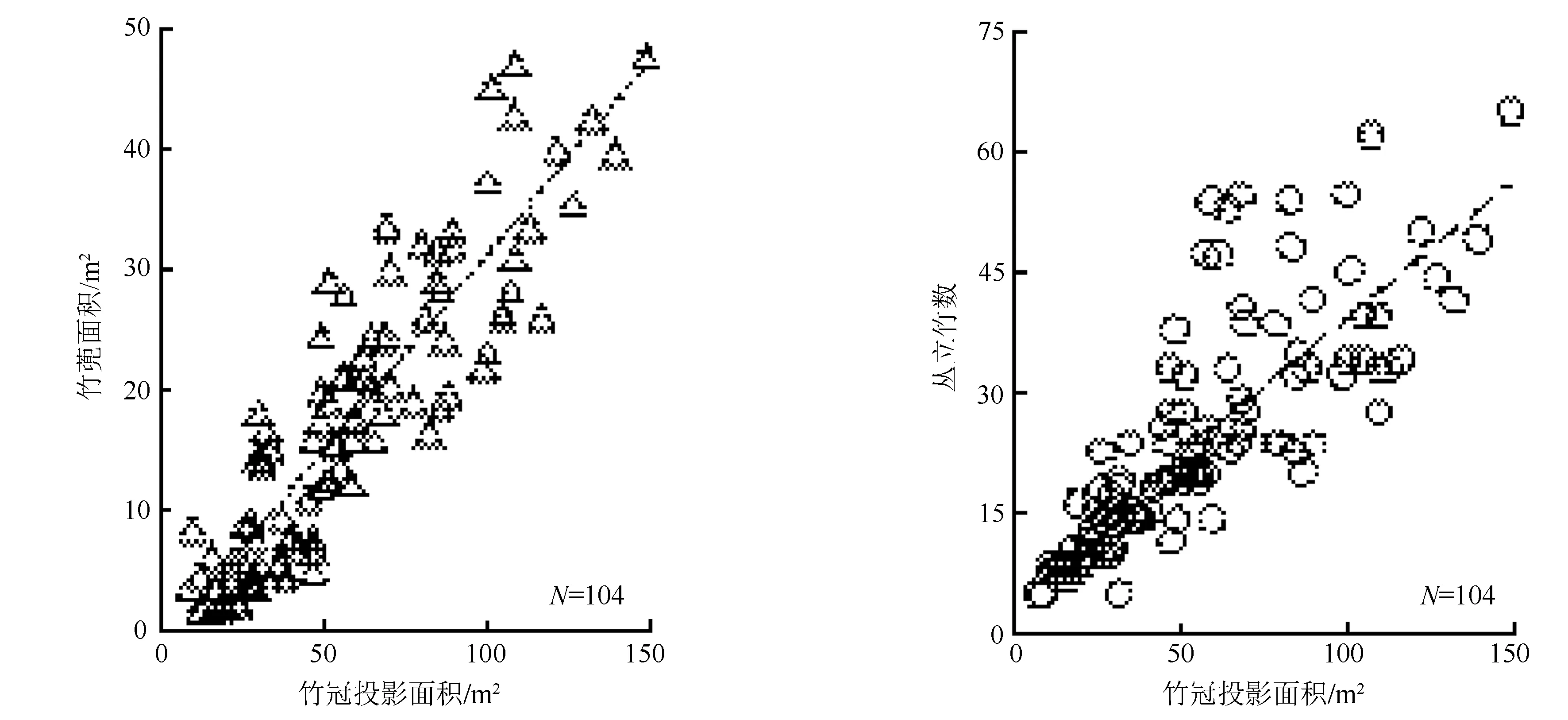
图2 竹冠投影面积与竹篼面积、丛立竹数分布关系
SBS=0.325 4×SPC-0.973 8,R=0.888 4;
NS=0.344 5×SPC+5.369 7,R=0.791 6。
式中:SBS为竹蔸面积;SPC为竹冠投影面积;NS为丛立竹干数;R为相关系数。
3.4 植被组成与重要值
林分共计植物种类73种,隶属于39科67属。其中豆科植物7种、禾本科植物6种、菊科植物5种、葡萄科植物4种,各占总种数的9.59%、8.22%、6.85%、5.48%;其余科属植物的占比则均在5.00%以下。
林分乔木层植被共计2科2属3种,分别为禾本科竹亚科牡竹属的巨龙竹、龙竹及桑科榕属的榕树,其中巨龙竹为绝对优势植物。
灌木层植被共计22科31属33种,其中乔木类植物10科11属12种、灌木类植物18科20属21种(表4)。各植物种重要值均小于30%,表明此林层无优势植物。乔木类植物的总种数少于灌木类,但总重要值(184.03%)远大于灌木类(115.97%),说明乔木类植物比灌木类发达,具有一定的生存优势。

表4 巨龙竹林分林下植被层物种重要值
草本层植被共计22科25属25种,其中1年生类草本5科6属6种、多年生类草本11科15属15种、蕨类植物4科4属4种。其中多羽凤尾蕨(Pterisdecrescens,重要值为39.89%)、破坏草(Ageratinaadenophora,重要值为30.80%)为竞争优势植物。多年生类草本植物(重要值为167.64%)相对发达。
层间植被共计7科11属13种,其中草质类藤本6科6属7种、木质类藤本4科6属6种。其中以荜拔(Piperlongum,重要值为40.01%)、异形南五味子(Kadsuraheteroclita,重要值为36.88%)为优势植物。林分内未发现大型木质类藤本植物。
3.5 林下植被多样性
林下植被物种多样性分析(表5),结果表明:物种丰富度、Shannon-Wiener多样性指数、Simpson多样性指数、Pielou均匀度指数在各林下植被层中的变化规律一致,其各指数由高到低依次为灌木层、草本层、层间植物,表明巨龙竹林下植被物种多样性以灌木层最高,草本层次之,层间植被最低。

表5 巨龙竹林分林下植被层的物种多样性
3.6 林下植被种-面积关系
方差分析表明,灌木层、草本层的植物物种数变化在3组重复样方间无显著差异(灌木层的P为0.77;草本层的P为0.33,P>0.05),其均值反映了其种-面积关系。灌木层、草本层植物物种数均随调查样方面积增大而增多,且当灌木样方面积达210.25 m2(14.5 m×14.5 m)、草本样方面积达90.25 m2(9.5 m×9.5 m)时,样方可分别涵盖林分85%的灌木、草本植物种类,且后续变化趋势不明显(图3)。灌木层、草本层的植物种-面积曲线拟合公式(自然对数线性转化)分别为:
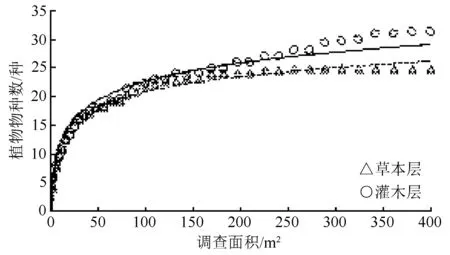
图3 林分林下植被“种-面积”关系
N1=4.693 5·lnA+1.058 8,R=0.974 8;
N2=3.839 7·lnA+3.176 6,R=0.982 9。
式中:N1为灌木层植物物种数;N2为草本层植物物种数;A为调查样方面积;R为相关系数。
4 讨论
4.1 林分直径生长与分布差异
2019年的林分直径生长较2018年有增大趋势,但其直径分布差异未达显著水平,这与2019年云南雨季降水量较往年明显减少有关(图4)。巨龙竹发笋和幼竹生长时的雨水较少,难以满足其生长、发育需求,所以其直径生长缓慢且较去年的变化不大[24]。此外,调查发现,林分内有竹鼠啃食竹蔸的现象,这在一定程度也会影响巨龙竹的生长、发育。

图4 2014—2019年云南雨季降水特征
4.2 丛立竹干数与竹干胸径的关系
林分密度与林木个体的直径生长一般呈负相关关系,即林分密度越大,个体间竞争越强,其直径越小。但本研究中,巨龙竹丛立竹干数与平均胸径呈显著低度正相关,即竹干胸径随丛立竹密度增大而增大。2017—2019年的林分直径、密度分布均呈逐年增大趋势,这与巨龙竹独特的生长特性有关,即新竹直径较老竹持续增大,从而避免因种源繁衍困难、分布稀少,而被自然淘汰。
4.3 巨龙竹对其林下生境、植物组成、物种多样性的影响
林下生境会因主林层的密度差异而形成不同的光照、生存空间,进而影响林下植被的分布类型、物种多样性[25]。本研究中,巨龙竹林分虽未有极高的密度特征,其林下生境条件仍较差,主要因为巨龙竹形态过于庞大,其资源竞争优势非常明显。通过竹冠垂直投影面积得出的林分郁闭度,忽略了巨龙竹在垂直方向上的冠层结构特征和生长的巨大优势。当林分处于非正午光照直射时,巨龙竹林下多为零星、稀薄的光照。巨龙竹凋落物对林地表层植被的影响巨大,林分内大量的凋落物干扰了林下植被正常的生长、更新。同时,巨龙竹硕大的干萚脱落时会大面积地覆盖林下幼嫩植被,使林地表层植被的生长、更新更为艰难。
灌木层植物因相对充足的林下空间、较弱的光照、足够的养分条件得以广泛生存,但其长势较差且未有任何优势种。林地表层植物则因上层植被、竹凋落物形成庇荫、狭窄的生境条件而倍受抑制,主要分布一些耐阴性的蕨类和多年生草本植被。在中低郁闭度、竹丛密度但林下生境条件却较差的情况时,巨龙竹林分灌木层的物种多样性大于草本层、层间植被[26-28]。此外,草本层的破坏草虽有极强的竞争优势,但其仅分布林分边缘;同时破坏草是入侵植物,能否将其视为巨龙竹林分草本层的优势植物还有待商讨。
5 结论
近自然生长的巨龙竹林分干龄分布呈右偏缓峰状;竹丛年龄分布呈左偏聚集状;直径分布呈左偏聚集状;丛立竹密度分布呈右偏缓峰状;且均逐年差异性地向更高龄、更大径阶、更高丛立竹密度的范围分布。利用威布尔分布模型能较好地反映巨龙竹林分直径分布、丛立竹密度分布规律。竹冠垂直投影面积与竹蔸面积、丛立竹干数均呈极显著高度正相关关系,可通过建立通用线性方程预估巨龙竹林分结构动态变化。
巨龙竹林分乔木层以巨龙竹为绝对优势植物;灌木层无优势植物;草本层以多羽凤尾蕨为优势植物;层间植被以荜拔、异形南五味子为优势植物,且未发现大型木质类藤本植物。巨龙竹林分植被物种多样性以灌木层最高、草本层次之、层间植被及主林层最低。今后,可将其灌木、草本调查样方分别设置为15 m×15 m、10 m×10 m,保证样方内均涵盖林分85%的灌木、草本植物种类。相关研究结果为今后该区巨龙竹资源及其群落物种多样性的保护、发展提供参考。
