水分胁迫对千金子种子萌发及生理生化特性的影响
2021-06-22薛静怡杨浩娜彭亚军王立峰
薛静怡, 杨浩娜, 唐 易, 彭亚军, 王立峰
(1.湖南大学研究生院隆平分院, 长沙 410082;2. 湖南省农业科学院生物技术研究所, 长沙 410125)
千金子[Leptochloachinensis(L.) Nees]是世界性农田恶性杂草,严重影响水稻生长,具有繁殖力强、旱生喜湿等特性。与水稻相比,千金子的分蘖多,且种子量大,平均每株结籽4.5万粒,具有很强的竞争力[1-3]。目前防治千金子以氰氟草酯等化学药剂防除为主,但化学药剂防除对千金子防治效果不佳,且环境污染问题严重[3]。千金子在萌发时期不耐水淹,水分条件对其萌发的影响较大[2]。而稻田可用水资源充足,建立适宜水层,在一定程度上可以抑制千金子出草量。
应对胁迫植物自身会产生渗透调节性物质,这类物质包括脯氨酸和可溶性蛋白等[4]。植物正常生长时,在呼吸作用和光合作用中会产生活性氧(ROS)[5]。水分胁迫等环境胁迫条件下会增加ROS的产生,从而导致氧化应激,过量的ROS通过氧化蛋白质、脂质和DNA来干扰正常的细胞新陈代谢[6]。氧化应激可能使植物细胞膜脂发生脂质过氧化现象,导致较多的丙二醛(MDA)产生;植物有一系列的酶协同工作来清除不同细胞器中的ROS[5]。通过探究水分胁迫对脯氨酸、可溶性蛋白、MDA和抗氧化酶等生理生化指标的影响,可以了解千金子种子应对水分胁迫、在维持渗透调节平衡和活性氧代谢平衡中的响应。本试验研究了水分胁迫对千金子种子萌发及生理生化特性的影响,旨在全面了解千金子种子应对水分胁迫的响应,探索抑制千金子种子萌发的适宜水分条件,为环保安全的防控千金子提供新思路。
1 材料与方法
1.1 材 料
供试千金子种子于2018年6月30日在湖南省常德市汉寿县(29°4′N,111°59′E)采集。
1.2 试验设计
挑选颗粒饱满的千金子种子1 000粒,用4 ℃无菌水浸泡24 h催芽,吸去多余水分备用。试验在温度为32 ℃,12 h(光)/12 h(暗),光照强度为12 000 lx的光照培养箱中进行。采用购于广东华兴石英砂厂的石英砂作为发芽介质,粒径为0.15~0.4 mm,在使用前经过如下处理:a.洗涤;b.消毒:将洗净的湿砂在130~170 ℃高温下烘干约2 h;c.加水拌匀。
实验前测定所用石英砂的饱和含水量,设置6个水分处理, 包括干旱处理和淹水处理。分别为砂床饱和含水量的50%(T 1)、75%(T 2)和100%(T 3),以及0.5 cm(T 4)、1 cm(T 5)和1.5 cm(T 6)的淹没处理。其中T 3为水分正常处理,也可以视为0 cm的淹水处理;T 1和T 2为干旱处理,T 4、T 5和T 6为淹水处理。计算6个水分处理所需的水量,在石英砂中分别加入相应含水量的蒸馏水,混合均匀后分装至200 mL塑料杯中,每杯均匀放入30粒种子,播种在砂床0.5 cm处,每个处理3次重复。使用保鲜膜将塑料杯封口,最后置于光照培养箱中培养。间隔12 h称重法定量补充蒸馏水,将蒸馏水加入到塑料杯内至原重量,以保持水分含量不变。
种子发芽以种子的胚根长达到种子长度,且胚芽达到种子长度的一半为标准[7]。
发芽势(%)=[(发芽初期(规定日期内)发芽种子数/供试种子总数)]×100%;
发芽率(%)=[(发芽终期(规定日期内)发芽种子数/供试种子总数)]×100%。
此次试验发芽初期为3 d,发芽终期为7 d。在第7天每个处理取30粒种子,用体式显微镜拍照,并用全自动超景深数码显微成像系统(机型:德国蔡司Smart Zoom 5)对胚根、胚芽的长度进行测量。
1.3 测定方法
1.3.1砂床饱和含水量的测定
取一底部为铁丝网的圆柱体,铁丝网上放一层湿润滤纸,称重为W1;在圆柱体中加满砂,砂上放一层干滤纸,称重为W2;将圆柱体置一水盆中,水面刚好淹至底部铁丝网,在砂上干滤纸中部刚好湿润时移开圆柱体,称重为W3,则砂床饱和含水量(%)=[(W3-W2)/(W2-W1)]×100%[7]。
1.3.2脯氨酸与可溶性蛋白含量的测定
将约0.2 g千金子种子样品液氮研磨,加3%的磺基水杨酸溶液5 mL,沸水浴15 min,自然冷却后取上清液,采用酸性水合茚三酮显色法进行脯氨酸的测定[8]。可溶性蛋白含量的测定采用考马斯亮蓝(G-250)法[9],试剂盒购自南京建成生物工程研究所。
1.3.3MDA含量的测定
将约0.1 g千金子种子样品液氮研磨,加0.05 mmol·L-1磷酸钠缓冲液(pH=7.8)5 mL,10 000 r·min-1离心5 min,取上清液,采用硫代巴比妥酸显色法进行MDA含量的测定[10]。
1.3.4抗氧化酶活性的测定
超氧化物歧化酶(SOD)活性的测定采用黄嘌呤氧化酶法[11]。过氧化物酶(POD)活性的测定采用愈创木酚比色法[12]。过氧化氢酶(CAT)活性的测定采用可见光钼酸铵法[11]。试剂盒购自南京建成生物工程研究所。
1.3.5数据处理与分析
采用IBM SPSS 20.0统计软件分析所有数据。ANOVA单因素方差分析,显著性水平设置为0.05。
2 结果与分析
2.1 水分胁迫对千金子种子萌发指标的影响
T 1和T 2处理下千金子种子极少发芽,大部分种子种皮未开裂,部分种子只长出胚根,胚芽未长出(图1 a,b)。T 3和T 4处理下千金子种子普遍发芽,胚芽与胚根均正常生长(图1 c,d)。T 5和T 6处理下千金子种子部分发芽,胚根生长缓慢,胚芽显著长于胚根(图1 e,f)。
从表1可以看出,随水分增加,胚根长先增加后下降,T 4处理的胚根最长;随水分增加,胚芽长逐渐增长, T 6处理的胚芽最长;胚根胚芽比先降低后增加再下降,T 1处理的胚根胚芽比最长。对胚根长、胚芽长、肧根胚芽比3个种子萌发指标分别进行单因素方差分析,发现T 3、T 4、T 5和T 6处理的胚根长差异不显著(p>0.05),T 1、T 2、T 5和T 6处理的胚根长差异不显著(p>0.05);T 1和T 2处理的胚芽长与T 3、T 4、T 5和T 6处理之间差异显著(p<0.05);T 1处理的胚根胚芽比与其他处理之间均差异显著(p<0.05),T 5和T 6处理的胚根胚芽比显著降低,与其他处理差异显著(p<0.05)。这可能是由于胚根对水多缺氧条件的反应比胚芽更为敏感[7]。种子萌动时,胚的生长随水分供应情况不同而存在差异。当水分相对少时,胚根先出;当水分过多时,胚芽先出,较胚根生长迅速。
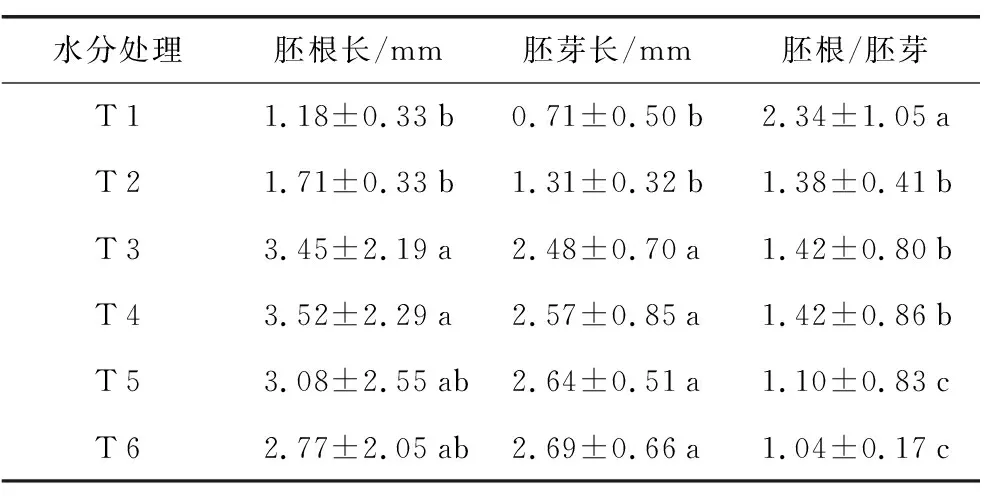
表1 不同水分处理对千金子种子萌发的影响Table 1 Effects of different water treatments on seed germination of L. chinensis
如图2所示,千金子种子的发芽率和发芽势呈先上升再下降的变化趋势。T 1处理千金子种子的发芽率和发芽势极低,仅为19.44%和11.11%。T 3处理下种子的发芽率和发芽势都达到最高值,之后随着水分的增加,千金子种子的发芽率和发芽势都逐渐下降。对发芽势和发芽率分别进行单因素方差分析,发现水分处理对发芽率影响均差异显著(p<0.05);T 3和T 4处理的发芽势之间的差异显著(p<0.05),T 2、T 5和T 6处理的发芽势之间差异不显著(p>0.05),T 1和T 6处理的发芽势之间的差异不显著(p>0.05)。由于T 5与T 6处理的发芽势差异不显著(p>0.05),在实际生产中为避免抑制同环境下作物发芽率,可以选择T 5处理抑制千金子的发芽来达到早期防控的目的。
2.2 水分胁迫对脯氨酸和可溶性蛋白含量的影响
应对胁迫种子自身会产生渗透调节性物质,渗透调节性物质可以维持种子细胞的水分平衡,保证种子的正常生长,这类物质包括脯氨酸和可溶性蛋白等[4]。如图3所示,随水分增加,千金子种子的脯氨酸和可溶性蛋白含量都呈先下降后上升的变化趋势,都在T 3处理时达到最小值。可溶性蛋白任意两水分处理下的指标含量差异显著(p<0.05);T 1和T 5处理的脯氨酸含量之间的差异不显著(p>0.05)。脯氨酸含量在T 6处理时达到最大值,可溶性蛋白含量在T 1处理时达到最大值,说明T 3水分正常处理下产生的渗透调节性物质含量显著低于其他处理,在干旱和淹水胁迫下渗透调节性物质含量升高,可以提高细胞的渗透调节能力。不同处理间脯氨酸含量表现为:T 6>T 1>T 5>T 2>T 4>T 3,说明淹水胁迫产生了更多的脯氨酸。不同处理间可溶性蛋白含量表现为:T 1>T 2>T 6>T 5>T 4>T 3,说明T 1和T 2干旱处理产生了更多的可溶性蛋白。随干旱和淹水胁迫程度的加深,脯氨酸和可溶性蛋白的含量呈增加趋势。
2.3 水分胁迫对MDA含量的影响
水分胁迫可能会引起种子活性氧代谢失控,种子细胞膜脂可能会发生脂质过氧化现象,导致较多的MDA产生,可以将MDA作为描述种子细胞膜脂过氧化程度的指标[4]。从图4可以看出,随水分增加,千金子种子的MDA含量总体呈先降后升的变化趋势,且任意两水分处理下的指标含量差异显著(p<0.05)。
T 3处理MDA的含量最低, T 6处理MDA的含量最高,说明T 3处理条件下膜脂质过氧化程度最低,MDA含量随胁迫程度的加深呈增加的趋势。T 1、T 2、T 4、T 5和T 6处理之间差异显著,且T 1>T 6>T 2>T 5>T 4 ,MDA的含量随胁迫程度的加深呈增加趋势。
2.4 水分胁迫对抗氧化酶活性的影响
种子细胞内抗氧化系统能调动多种保护酶应对胁迫,清除内部的过氧化物和自由基,以维持正常生长。细胞内抗氧化系统常见的保护酶有SOD,POD等[13]。
如图5所示,随水分增加,千金子种子的SOD和POD活性总体呈先下降后上升的变化趋势,均在T 1处理达到最大值,在T 3处理达到最小值。任意两水分处理下的SOD和POD活性差异显著(p<0.05)。SOD与POD二者最低值均出现在T 3,说明随干旱或淹水胁迫加重,SOD和POD活性增加,可加速分解种子内部产生的超氧自由基,减缓膜脂过氧化速度,与上述MDA含量的变化一致。不同处理间SOD活性表现为:T 1>T 6>T 2>T 5>T 4>T 3,说明重度的干旱和淹水胁迫(T 1和T 6)下SOD活性较轻度胁迫(T 2、T 4和T 5)表现为不同程度的升高,与上述MDA的现象相符。且重度的干旱和淹水胁迫不利于种子的自我调节修复,表现为发芽率的降低。不同处理间POD活性表现为:T 1>T 2>T 6>T 5>T 4>T 3,说明淹水胁迫下的POD活性较干旱胁迫低。
3 讨论与结论
温度、光照和水分对千金子的萌发至关重要,千金子一般在夏季萌发,此时的温度和光照条件均适宜千金子种子萌发和生长,因此水分就成为影响千金子萌发出苗的关键因子[2]。
本研究中,在T 1处理下千金子种子的发芽率和发芽势最低,在T 3处理下种子的发芽率和发芽势最高,之后随着水分的增加,千金子种子的发芽率和发芽势都下降。说明种子受到干旱胁迫时,显著抑制种子的发芽;种子受到淹水胁迫时,也对种子的发芽有一定程度的抑制。T 1处理时胚根/胚芽最大,T 6处理时胚根/胚芽最小,随着水分的增加,呈先降低后增加再降低的趋势。说明干旱胁迫下,胚根较胚芽优先生长,可以通过胚根吸收能力的提高缓解干旱胁迫[14];淹水胁迫下,由于胚根对氧的利用减少,胚根的细胞进行无氧呼吸,乙醇等有害物质大量积累,导致根系细胞死亡[15]。
研究表明,植物受到水分胁迫后,会造成渗透胁迫,细胞可通过合成更多的可溶性蛋白和脯氨酸使植物细胞渗透势增加,提高细胞的渗透调节能力以维持正常生长[15]。本实验中,脯氨酸含量在T 6处理时达到最大值,可溶性蛋白含量在T 1处理时达到最大值。淹水胁迫产生了更多的脯氨酸,T 1处理和T 2处理干旱处理产生了更多的可溶性蛋白。
当植物处于适宜的环境条件下,ROS的产生和清除存在动态平衡,植物受到环境胁迫后ROS的代谢平衡被打破,ROS含量升高导致MDA积累,使膜脂质过氧化[15-16]。植物会通过启动细胞内与ROS平衡相关的抗氧化系统,如抗氧化酶系清除过量的ROS从而降低膜脂的过氧化损伤[17]。本研究中,MDA含量总体呈先下降后上升的变化趋势,T 3处理MDA的含量最低,T 6处理MDA的含量最高;T 1>T 6>T 2>T 5>T 4>T 3,说明干旱与淹水胁迫均导致种子内部活性氧代谢平衡被打破,导致MDA含量的增加,细胞膜系统的损伤。本试验中,SOD与POD的活性在T 1处理下最高,活性随水分增加降低,在T 3处理下达到最小值,之后随水分增加逐渐升高,且SOD和POD活性随干旱或淹水胁迫加重都表现为不同程度的升高。
综上所述,干旱能显著抑制千金子种子发芽,淹水胁迫也对种子发芽有一定程度的抑制,且干旱处理下胚根较胚芽优先生长,淹水处理下缺少氧气胚根缓慢生长。在干旱和淹水胁迫下脯氨酸和可溶性蛋白含量较T 3处理显著升高,且与胁迫程度呈正比。随干旱或淹水胁迫程度的加深,MDA的含量增加,相应的SOD和POD的活性也增加,以进行自我调节修复。由于T 5处理与T 6处理的发芽势差异不显著(p>0.05),T 1处理与T 5处理的脯氨酸含量差异不显著(p>0.05),T 4处理与T 5处理的可溶性蛋白、脯氨酸和MDA含量以及SOD和POD活性之间的差异显著(p<0.05),且在实际生产中为避免抑制同环境下作物发芽率,推荐选择T 5处理(淹水1 cm)抑制千金子种子的发芽来达到早期防控千金子的目的。
