不同种源野鸦椿一年生苗生长规律研究
2021-06-22陈菊艳陈志萍邓伦秀
陈菊艳, 陈 锐, 陈志萍, 邓伦秀
(贵州省林业科学研究院, 贵阳 550005)
野鸦椿(Euscaphisjaponica)为省沽油科(Staphyleaceae)野鸦椿属(Euscaphis)的落叶小乔木或灌木[1-3],又名鸡肾果、鸡眼椒、芽子木、蝴蝶果、山海椒、情人果、酒药花、红三季等[4],是具有地域特色的多年生、低维护、低成本、三至四季均可观赏,同时药用价值极为珍贵的一种植物[5]。野鸦椿冠形舒展、树形优美,春夏之交盛开黄白色花朵,满树银花,秋天红果满枝,集赏花、观果和观叶为一体,是一种理想的“一季观花,三季观果”树种,可作为观赏树种用于园林绿化。野鸦椿的药用价值也很高,果实、种子、根和根皮、花均可入药,具有解毒、清热、利湿、祛风散寒、行气止痛等功效。此外,野鸦椿木材还可用于制作器具,种子提取油可用于制皂,树皮可提取栲胶[1-3]。野鸦椿资源分布区域较为零星广泛,主要分布于长江以南各省区,野生于山坡、山谷、河边的丛林及灌丛中,在贵州大部地区均有分布[6]。多搭配乔、灌木混生,极少为单纯片植的林子[7]。我国从20世纪50年代开始对野鸦椿进行研究,主要集中在形态分类学、解剖学与生态生物学[8-9]、药理开发[10-15]、繁殖栽培技术[16-18]、选择优良品种[19]、植物生理生化[20]等方面。
目前,有关野鸦椿苗期试验的研究,未见以苗期生长规律为主的相关报道,本研究在贵州野鸦椿自然分布区选取了5个代表种源进行苗期对比试验,揭示不同种源野鸦椿一年生苗木在贵州的生长规律,以期为科学培育野鸦椿优质壮苗和选育适合贵州种植的优良种源提供依据。
1 材料与方法
1.1 试验地概况
试验地选择在贵州省林业科学院树木园,位于贵阳市西南郊,106°43′E,26°30′N,海拔1 100 m,属于亚热带高原季风湿润气候,气候温和,冬无严寒,夏无酷暑,年平均气温15.2 ℃,极端最低气温-7.3 ℃,极端最高气温37.5 ℃,降雨主要集中在5—9月,年平均降雨量1 198.9 mm,平均相对湿度77%,无霜期278 d[21]。
1.2 试验材料
试验材料选择贵州不同地区的5个种源:遵义市道真县(107.58°E,28.82°N,海拔1 420~1 620 m)、毕节市大方县箐梁子(105.10°E,25.71°N,海拔1 521~1 760 m)、榕江县月亮山(108.24°E,25.63°N,海拔1 148 m)、黔西县(105.87°E,27.29°N,海拔870~1 220 m)、纳雍县纳雍林场场部工区(105.26°E,26.67°N,海拔1 950~1 665 m)。
1.3 方 法
种子于2011年10月分别在5个种源地进行采集并沙藏保存两年,待种子10%露白即可进行田间播种,播种方式为宽幅条播,行距15 cm,将种子置于播种沟内,深1~1.5 cm,覆盖0.5~1 cm厚腐熟树皮,播种后搭农膜小拱棚,外盖遮阳网。播种后1个月左右开始出苗,出苗整齐后及时间苗、除草,合理进行水肥、病虫害防治等管理,并揭开农膜。对5个种源随机选定的30株标准株进行挂牌编号,自2013年5月8日起,至2013年11月30日为止,每10 d进行一次苗高和地径的测定。苗高生长量用钢尺测量(精确到0.1 cm),地径用游标卡尺测量(精确到0.01 mm)。
1.4 数据处理
Logistic方程能够很好的模拟植物生长过程,野鸦椿不同种源苗高和地径生长动态采用Logistic生长方程进行拟合[22-24],其模型表述和拟合过程以及生长时期的划分见文献[25]。
利用Excel 2008软件进行数据的整理和图、表处理;用SPSS 18.0软件进行方差分析、Logistic方程拟合以及其他统计分析处理。
2 结果与分析
2.1 野鸦椿一年生苗木生长节律变异分析
不同种源野鸦椿一年生苗的苗高和地径的生长量见表1。一年生苗木苗高生长以纳雍种源表现最优(平均苗高33.174 cm);其次是黔西种源(平均苗高24.672 cm)、大方种源(平均苗高23.768 cm)、榕江种源(平均苗高20.876 cm),表现最差的是道真种源(平均苗高15.490 cm)。地径生长以黔西种源表现最优(平均地径8.217 mm),其次是纳雍种源(平均地径7.835 mm)、大方种源(平均地径7.012 mm)、榕江种源(平均地径6.713 mm),表现最差的是道真种源(平均地径6.181 mm)。方差分析结果(表2)显示,苗高、地径在种源间存在极显著差异。

表2 不同种源野鸦椿一年生苗苗高、地径的方差分析Table 2 Variance analysis of annual seedlings ofE. japonica from different provenances
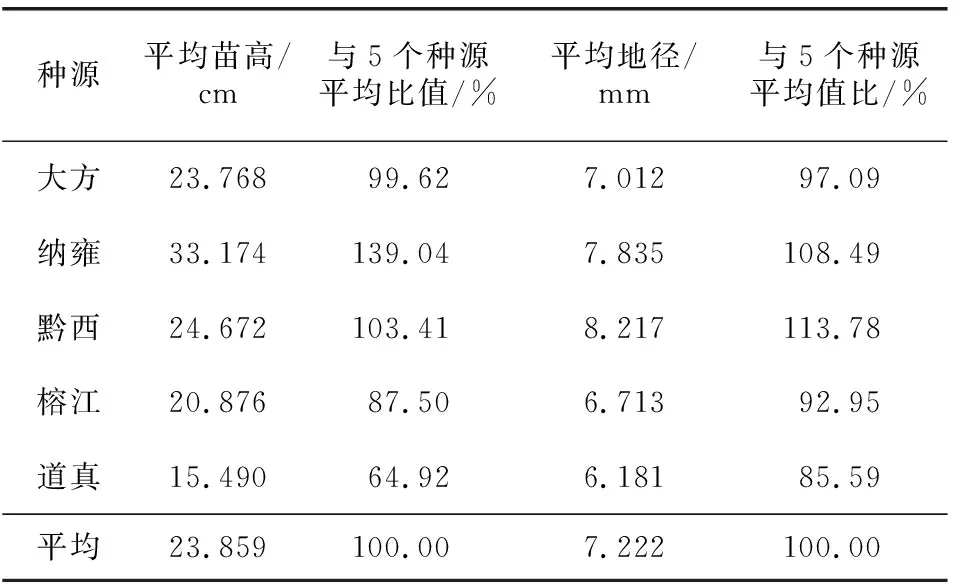
表1 不同种源野鸦椿一年生苗的苗高、地径生长量比较Table 1 Comparison of seedling height and ground diameter growth of annual seedlings of E. japonica from different provenances
2.2 野鸦椿一年生苗木生长节律模型的建立与拟合
为了解野鸦椿一年生苗的年生长节律,利用Logistic方程分别对其生长动态进行拟合,其拟合曲线见图1、图2。回归曲线的方程和参数估计值见表3。
从表3可知,野鸦椿一年生苗的年生长动态与“S”型生长曲线拟合均达到极显著水平。一般认为相关系数接近于1,其回归方程的结果就比较可靠,野鸦椿苗高拟合方程相关系数最小为0.957,最大为0.988,平均为0.969;地径拟合方程的相关系数最小为0.984,最大为0.992,平均为0.988。5个种源苗木生长总体拟合程度与实测值相吻合,说明用Logistic方程的理论值可以较准确地估测不同种源苗木生长的实际值。

表3 不同种源野鸦椿一年生苗苗高、地径的Logistic回归及相关参数Table 3 Logistic regression of seedling height and ground diameter and related parameters of annual seedlings ofE. japonica from different provenances
结合野鸦椿不同种源苗高拟合曲线(图1)可以看出,5个种源野鸦椿一年生苗木的苗高均呈现明显的“慢—快—慢”的“S”型生长节律,在215 d观测期内,播种32 d后苗木出齐,5个种源的苗高生长趋势基本相似,32 d到80 d苗高生长缓慢,80 d以后生长量开始迅速增加,180 d左右苗高生长减缓。但各种源苗木在各阶段的生长特点存在明显差异。纳雍种源苗木的苗高在生长初期没有大方种源生长得快,但是到生长中后期就进入生长旺盛阶段,生长较好,苗高累积生长量表现最好,大方种源苗高生长前期较快,到后期逐渐变慢,榕江种源和黔西种源苗高生长节律相似,道真种源生长平稳增长,苗高累积生长量较其他种源缓慢。各种源的苗高在215 d的生长期内生长量分别为,大方23.768 cm,纳雍33.174 cm,黔西24.672 cm,榕江20.876 cm,道真15.49 cm,总体来看,纳雍种源苗高生长要强于其他种源。
不同种源野鸦椿的苗高增长高峰期有所差异,根据计算结果(表3)可以看出,生长高峰期主要集中在7月30日—8月23日,黔西、榕江和纳雍种源的苗高增长高峰期分别在出苗后124 d、108 d、109 d,对应生长速度最快的时间分别为8月23日、8月7日和8月8日,道真和大方种源苗高生长高峰期均出现在出苗后的100 d(7月30日),各种源中高峰期出现较晚的是黔西种源,较早的是道真和大方种源。生长初期苗高生长趋势基本一致,从7月初开始不同种源苗高生长出现明显差异。
从图2可知,不同种源野鸦椿地径年生长过程也呈现“S”型生长节律。生长初期(播种至80 d)地径增加十分缓慢,生长盛期(80 d至180 d)地径迅速生长,生长后期(180 d以后)地径生长逐渐减缓直到停止生长。各种源地径生长速度接近、生长曲线相似。大方种源苗木地径生长前期较快、生长量高于其他种源,但是到中后期地径生长速度变慢,没有纳雍种源地径增长快,表明大方种源地径生长前期竞争力较强[26];纳雍种源苗木地径前期生长仅次于大方种源,到了中后期地径生长较快、生长量较高,表明纳雍种源后期竞争力较强;榕江种源苗木地径各时期平稳增长,在各种源中地径累积生长量排在第三,道真种源苗木地径在生长初期没有黔西种源生长快,但是到了后期黔西种源超过了道真种源,在各种源中地径累积生长量表现出道真种源生长较慢。
5个种源地径增长高峰期也不相同,黔西种源地径增长高峰期在出苗后第140天,道真、纳雍和榕江种源地径增长高峰期分别在出苗后115 d、111 d和106 d,这3个种源高峰期时间相差不大,大方种源地径增长高峰期出现最早,在出苗后95 d。
从苗高和地径生长情况可以看出(图1、图2),地径生长停止的时间与苗高生长停止的时间不一致,苗高在10月中旬左右停止生长,而地径在11月上旬左右停止生长,苗高、地径生长期相差半个月左右。
2.3 野鸦椿一年生苗木生长阶段的划分
2.3.1苗高生长阶段的划分及特点
据前所述,野鸦椿一年生苗木生长节律呈现明显的“慢—快—慢”的“S”型生长曲线,表明其具有明显的阶段性。根据拟合模型、计算结果及其生长特点,将野鸦椿苗木整个生长时期划分为出苗期、生长初期、速生期和生长后期4个阶段,各阶段起始时间、各阶段持续天数、净生长量和生长量占总生长量的百分比见表4。5个种源各阶段的生长特点存在差异,在出苗期,生长时间都相同(从3月20日到4月21日),持续时间均为32 d,但出苗期净生长量有差异,表现为纳雍种源的净生长量较高,为1.909 cm,道真种源的较低,为0.731 cm;各种源生长初期长短和结束时间不同,纳雍种源生长初期持续时间最长,为64 d,净生长量为6.268 cm。前期生长较快的种群,能在相同时间和相同的生长环境下快速占据生长空间,有利于苗木后期的生长发育[27],纳雍种源就是这样的种类,因此苗高总生长量最高;黔西、榕江及大方种源生长初期持续时间分别为57 d、47 d、39 d,道真持续时间最短,为30 d,道真种源最早进入速生期(5月22日),且速生期持续时间最长,为138 d,纳雍种源最晚进入速生期(6月25日),速生期持续时间最短,为87 d,但苗木净生长量最多,为19.677 cm,榕江、黔西、大方3个种源分别于6月8日、6月18日、5月31日进入速生期,且速生期持续时间分别为120 d、131 d、120 d,5个种源速生期净生长量明显高于其他生长阶段,速生期净生长量占总生长量比重最高的是黔西种源,为69.89%,其次是大方种源,为66.79%,再次是道真种源,为66.38%,第四是榕江种源,为65.64%,最低的是纳雍种源,为59.48%;5个种源进入生长后期的时间和生长后期持续时间均不同,纳雍种源最早进入生长后期(9月20日),且持续时间最长(71 d),黔西种源最晚进入生长后期(10月28日),且持续时间最短(33 d),榕江、道真、大方种源持续天数分别为53 d、54 d、63 d,进入生长后期,苗高生长减缓,净生长量占总生长量比重最高的是纳雍种源,为15.81%,其次是榕江种源,为6.75%,再次是榕江种源,为6.74%,第四是道真种源,为3.18%,最低的是黔西种源,为0.61%。野鸦椿各种源基本于9月下旬到10月上旬停止生长,苗木整个生长期约200 d。

表4 不同种源野鸦椿一年生苗苗高和地径生长时期的划分及生长节律比较Table 4 Division of seedling height and ground diameter growth period andcomparison of growth rhythm of annual seedlings ofE. japonica from different provenances
2.3.2地径生长阶段的划分及特点
根据Logistic曲线及其特点,可从理论上进行野鸦椿地径生长时期的划分及生长情况比较(表4)。各种源在出苗期生长时间都相同,生长初期不相同,表现为黔西种源生长初期时间较长,为79 d,净生长量为2.491 cm,大方种源生长初期持续天数最少,为47 d,净生长量为2.388 cm;5个种源中大方种源较早进入速生期,黔西种源较晚,其他种源居中,但各种源进入速生期的时间大都在6月初到7月初。经计算分析可得野鸦椿5个种源地径生长的速生期分别为:道真种源从6月15日至10月13日,持续天数为120 d,速生期内净生长量为3.689 mm,占整个生长量的61.86%;黔西种源速生期为7月10日至11月06日,持续天数也为120 d,净生长量为5.351 mm,占整个生长量的46.93%;其他3个种源速生期持续天数和净生长量分别为榕江93 d,净生长量3.693 mm,纳雍95 d,净生长量4.368 mm,大方94 d,净生长量3.291 mm。5个种源进入生长后期的时间和生长后期持续时间均不同,大方种源最早进入生长后期(9月10日),且持续时间最长(81 d),黔西种源最晚进入生长后期(11月7日),且持续时间最短(23 d),道真、榕江、纳雍种源持续天数分别为47 d、70 d、64 d,生长后期地径净生长量占总生长量比重最高的是纳雍种源,为15.75%,其次是大方种源,为13.82%,再次是榕江种源,为13.49%,第四是道真种源,为3.17%,最低是黔西种源,为1.18%。由野鸦椿各阶段苗木的生长情况进一步说明,除出苗期以外,苗高生长各阶段的持续时间和地径生长是不同的,大方、榕江、道真和黔西种源苗高比地径提早半个月左右进入速生期,而纳雍种源地径比苗高提前2 d进入速生期,但是不同种源间苗高和地径都表现为速生期生长量所占比重最大,速生期持续时间占总生长期的一半,速生期内苗木累积生长量占总生长量的60%以上,可见,速生期是苗木整个生长过程中的关键时期[28]。
3 结论与讨论
1) 对5个种源野鸦椿一年生苗的累积生长量进行方差分析表明,各种源间存在极显著差异。总体表现出纳雍种源较好,其他种源生长势虽比纳雍种源稍差,但是生长表现良好,一年生苗高均在15.0 cm以上,均能适应栽植地的环境。
2) 利用Logistic方程拟合野鸦椿各种源的生长节律曲线,方程的回归性均达到极显著水平,表明Logistic方程能很好地拟合出不同种源的生长趋势。在215 d观测期内,播种32 d后苗木基本出齐,5个种源的苗高生长趋势基本相似,32 d到80 d苗高生长缓慢,80 d以后生长量开始迅速增加,180 d以后苗高生长逐渐减缓直到停止生长,地径生长规律和苗高相似,但是地径晚半个月左右结束生长初期进入速生期,也晚半个月左右停止生长,各种源的苗木生长过程均呈现“慢—快—慢”的“S”型生长节律,但苗高和地径生长持续时间不一致,地径在苗高结束生长后半个月左右停止生长,苗高、地径生长期相差半个月左右。
3) 根据Logistic模型的特点和苗木生长具有明显的阶段性,将苗木年生长过程划分为出苗期、生长初期、速生期和生长后期4个阶段[29-31]。5个种源出苗期时间都一致,各种源生长初期持续时间和结束时间不同,纳雍种源生长初期持续时间最长,净生长量也最多,道真种源生长初期持续时间最短,净生长量最少。道真种源的苗高速生期开始时间早于其他4个种源,速生期持续时间最长,但是净生长量最少,纳雍种源速生期持续时间最短,但是净生长量最多,纳雍种源速生期占全年总生长时期的1/3,净生长量占全年总生长量的2/3,其他种源速生期占全年总生长时期的1/2,但是净生长量所占比例和纳雍种源一样,可见,纳雍种源苗高生长速生期短而净生长量增长快,而其他种源苗高生长速生期长而净生长量增长慢。5个种源的地径生长速度接近、生长曲线相似,大方种源最早进入速生期(6月8日),黔西种源最晚进入速生期(7月10日),其他3个种源分别于6月中下旬进入速生期,在速生期内黔西种源的净生长量最大,净生长量占总生长量的比重均最大,纳雍种源净生长量次之,道真和黔西种源速生期较长(均为120 d),但在生长盛期黔西种源的苗木地径生长量最大,而道真种源地径不是最大。5个种源在生长后期净生长量均较小,但还有缓慢的增长趋势。到11月初基本停止生长。
从苗木的生长表现来看,纳雍种源明显优于其他种源,优势主要表现在苗高的生长量上,同时其速生期短而生长速度快,进入生长后期较其他种源早,且进入生长后期以后苗高还在不断生长;黔西种源也具有较好的生长特性,主要表现在地径的累积生长量上,其速生期比其他种源晚,进入生长后期也晚,生长期内苗木的地径均高于其他几个种源。苗木生长规律的研究和客观地划分苗木生长阶段,可为科学地培育优质壮苗提供依据,从而人为地加快苗木生长[32]。生长初期主要是根系入土生长,在这个时期苗木嫩弱,抗性差,应对其进行适当的遮阴处理,并间苗移植、适时灌溉,避免土壤干旱;在进入速生期前应拆除遮阴棚,进行1~2次间苗,并适当施低氮高磷的混合肥,为速生期苗木的快速生长打下基础。在速生期内苗高和地径迅速增加,生物量积累最快,应加强水肥管理,可每月施尿素1~2次,同时注意中耕除草,争取使苗木的生长量达到最大;生长后期主要为地径的增粗生长,应在9月中旬(进入生长后期的时候),追施1次氮、磷、钾混合肥,之后应停止施氮肥,多施磷、钾肥,并减少灌溉,一方面促进其高生长,另一方面促进苗木根系的生长和枝干的木质化,提高苗木的抗旱性和抗寒性。
优良种源的选择,是苗木培育成功的关键因素之一,为提高苗木的造林成活率及出圃质量,在育苗时要有目的地选择生长特性较优、抗性较强的苗木种源,并在各个生长阶段采取不同的管理措施,更好地培育优质壮苗[33]。本研究是针对野鸦椿苗期开展的试验,由于苗期性状还不太稳定,各种源间的差异还有待进一步的造林对比试验,本研究的结果只作为野鸦椿苗期优良种源选择的参考。
