抗菌肽及其在食物储藏与保鲜中的应用
2021-06-21王小方焦路阳
杨 悦,李 燕,王小方,汪 磊,焦路阳*
(1.河北地质大学 数理学院,河北 石家庄050031;2.新乡医学院第一附属医院 检验科,河南 新乡453100)
对抗生素的过分依赖和滥用导致细菌耐药菌株,如耐甲氧西林金黄色葡萄球菌(MRSA)[1]、耐万古霉素粪肠球菌(VER)[2]、NDM-1超级细菌[3]等的出现,引起了全世界范围的普遍担忧,“禁抗”呼声越来越高,研发能替代抗生素的新型抗感染产品成为当务之急。抗菌肽(antimicrobial peptides,AMPs)广泛存在于多种生物体内,是宿主固有免疫系统的一类抵抗外源性病原微生物的天然免疫小分子多肽。网站http://aps.unmc.edu/AP/main.php公布了3 800多种抗菌肽。天然抗菌肽通常由10~40个氨基酸残基组成[4],具有广谱抗菌、活性稳定、免疫原性低,绿色无公害等优点[5-6]。不同生物体产生的抗菌肽具有特定的抗菌谱,具有一定的免疫学活力[7],可特异性抑杀细菌、真菌、病毒、支原体、寄生虫、肿瘤细胞,对正常的真核细胞几乎无毒性或只有低毒性,因此抗菌肽一直被视为理想的抗生素替代物。
在食品工业中,加入防腐剂可抑制致病菌的增殖,从而降低致病菌引发人类患病的风险。防腐剂(图1)分传统和天然防腐剂。传统防腐剂用于食品中常常具有副作用[8],如亚硝酸盐和硝酸盐与白血病、结肠癌、膀胱癌和胃癌有关,山梨酸盐和山梨酸与荨麻疹和接触性皮炎有关,苯甲酸盐可能与过敏、哮喘和皮疹有关。抗菌肽作为天然防腐剂,可以抑制致病菌的繁殖,且热稳定性好,安全无毒害,在食品的防腐和保鲜方面具有广阔的应用前景。乳铁蛋白,蛙皮素,乳酸链球菌素(nisin)等已被广泛应用于食品的防腐和保鲜领域中。
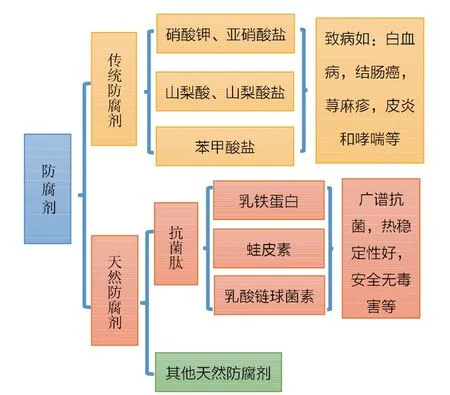
图1 传统防腐剂和抗菌肽的特点Fig.1 Characteristics of traditional preservatives and antibacterial peptides
AMPs种类繁多,根据生物学来源分为植物来源、动物来源和微生物来源3大类[9]。植物抗菌肽于1942年首次发现[10],后相继从小麦、苦参、拟南芥、马铃薯等分离得到,主要包括硫堇(thionins),植物防御素(defensins),脂质转移蛋白(LTPs),黑芥子酶(myrosinase)结合蛋白,橡胶蛋白 (heveins)和knottins样多肽,蛇毒、环肽以及水解物中的肽。动物抗菌肽最早是由瑞典科学家Boman等人发现的天蚕素(cecropins),后继从动物体内获得的magainin,MSI-78,PR-39,pleurocidin以及dermaseptin S4等均具有抗菌活性。微生物抗菌肽包含nisin,polymycin和片球菌素(pediocin)等。抗菌肽的氨基酸序列和结构具有显著差异,其共性为:均为阳离子肽,且具有两亲性的活性结构。抗菌肽可分别归属为α-螺旋、β-折叠、β-发夹、扩展和环状结构。Indolicidin呈现扩展结构,无固定的构象[11],LL-37在胶束(micelles)模拟膜中呈现典型的α-螺旋结构[12]。
1 抗菌肽的制备方法
抗菌肽的生产成本是制约其作为临床特效药和新型防腐剂商业化推广应用的一个重要因素。天然提取和人工合成抗菌肽成本高,产量低,且污染环境。相比于传统的天然提取和人工合成,基因工程法有着可以大规模生产,成本低等潜在优势,是生产抗菌肽的有效方法。近些年分子生物学和DNA重组技术的发展也为抗菌肽的基因工程化提供了良好的技术支持[13]。基因工程制备抗菌肽主要利用大肠杆菌表达系统,酵母表达系统,杆状病毒-昆虫细胞表达系统,植物表达系统和动物表达系统[14]。
大肠杆菌表达系统中,大肠杆菌成本低、易培养、生长迅速,成为最常用的宿主细胞[15-16]。但抗菌肽具有抗菌活性,会对宿主菌产生杀伤作用,而且易被宿主菌的蛋白酶降解[17]。抗菌肽添加融合标签共表达可防止抗菌肽对宿主菌的杀伤而提高产量,把抗菌肽表达为包涵体可防止宿主菌的蛋白酶对抗菌肽的降解[16,18-19]。融合表达蛋白包括谷胱甘肽转移酶(glutathione transferase,GST)、麦芽糖结合蛋白(maltose-binding protein,MBP)、硫氧还蛋白(thioredoxin,Trx)、酰胺磷酸核糖基转移酶(amidophosphoribosyl transferase,PurF)等[17]。臭鲑蛙皮肤分泌抗菌肽ORBK在大肠杆菌中已与MBP融合蛋白共表达过,并进行了分离纯化[20]。重组抗菌肽OsTHION15可通过大肠杆菌Rosetta-gami(DE3)来制备[21]。
酵母表达系统中,单细胞真核生物的酵母可以直接表达出有活性的抗菌肽,该抗菌肽对酵母无毒性,其基因表达调控机制较大肠杆菌更完备,具备对表达产物分泌和加工修饰能力。酵母菌主要有酿酒酵母(Saccharomyces cerevisiae),毕赤酵母(Pichia pastoris),多形汉逊酵母(Hansenula polymorpha)和耶罗维亚酵母(Yarrowia lipolytica)。其中酿酒酵母和毕赤酵母应用最为广泛。毕赤酵母可有效表达重组蛋白HD5,hBD2,羊的BD1或猪的BD2[22-24]。LL-37可通过毕赤酵母的表达系统制备[25]。酵母系统的缺点是在28℃的不稳定性和在发酵培养基中的蛋白水解活性太高[26]。
杆状病毒-昆虫细胞表达系统中,昆虫/杆状病毒表达系统安全,易于操作,能够高效表达大的外源基因,可对表达的外源蛋白进行修饰[27],但表达成本高,而且还需要病毒转导以及病毒库等[28]。抗菌肽牛乳铁蛋白N端和C端可通过昆虫/杆状病毒系统表达。
植物表达系统中,植物因其易生长、成本低、受内毒素和人类致病菌污染风险小的特点,具有大规模纯化的潜力[32],成为有前景的抗菌肽生物工厂系统。植物表达系统制备的cecropin A能抵抗细菌和真菌病原体[32-33]。植物表达系统表达外源蛋白,通常是整合到植物叶绿体基因组[34-37]。其制备的重组蛋白产量高,可高达2 kg[37]。线状阳离子抗菌肽cecropin A可通过水稻种子制备[32],CanThio1-8可由辣椒在其花中制备[38],紫花苜宿中的OsTHION15在其种子、叶子、幼苗、根和花絮等组织器官中均有表达。
动物表达系统中,动物能够促使蛋白质正确折叠,提供翻译后加工功能,产生类似于人类的天然蛋白质,成为生产重组肽的首选。CHO细胞系是制备蛋白质的主要哺乳动物宿主[29]。CHO培养体系是大型搅拌槽生物反应器[30]。随着培养基组分的改进和生物工艺技术的发展,其产量可达到1~5 g/L[31]。
2 抗菌肽的作用机制
抗菌肽的作用模式分为结合,插入及孔道形成3个步骤[39],主要有桶板模型、环孔模型和毯式模型,其中抗菌肽与细胞膜结合是抗菌肽发挥活性的必要条件。与传统抗生素对细菌靶点或代谢通路发挥作用的方式不同,抗菌肽特异性靶向细胞膜,作用于非特异性的分子靶点并迅速破坏细胞膜。
原核生物和真核生物细胞膜成分不同,抗菌肽特异性破坏原核细胞膜结构而杀死细菌[40]。原核生物细胞膜中的磷脂主要是带有阴离子的磷脂酰甘油,磷脂酰丝氨酸和心磷脂,且阴离子脂质暴露于表面,而抗菌肽为阳离子多肽,它们之间发生相互作用,第一作用力是带正电氨基酸和带负电细胞膜之间的静电作用,第二作用力是肽的双亲性结构域与膜磷脂之间的疏水相互作用。如果达到抗菌肽的阈值浓度,上述的这些作用力是剧烈的,进而达到对细菌细胞膜的破坏。而真核生物细胞膜中主要是两亲性磷脂,阴离子脂质位于细胞内侧[41],故抗菌肽对正常的真核生物细胞一般不起作用。
抗菌肽的具体抗菌方式取决于细菌的结构组成。革兰氏阳性菌和革兰氏阴性菌结构组分不同,故抗菌肽的作用机制也存在差异(图2)。革兰氏阳性菌和革兰氏阴性菌均含有肽聚糖。革兰氏阳性菌的肽聚糖裸露于细胞外,数层层叠在一起和磷壁酸共同构成细胞壁。革兰氏阴性菌的细胞壁则由外膜和周质间隙组成,细胞壁除含有一层肽聚糖外还有较厚的脂类层[42]。脂多糖(lipopolysaccharide,LPS)是革兰氏阴性菌膜外层的主要脂质成分。抗菌肽在与革兰氏阳性菌作用时,主要作用于革兰氏阳性菌的磷壁酸;与革兰氏阴性菌作用时,通过解离带负电荷的LPS胶束发挥作用。
真菌具有外壳——细胞壁。壳聚糖是细胞壁的主要组分[43]。抗菌肽的抗真菌机制包括:与真菌细胞壁壳聚糖有效结合,干扰真菌细胞壁合成;与细胞膜中的麦角甾醇/胆固醇结合,破坏真菌细胞膜结构;解聚肌动蛋白细胞骨架以及靶向细胞内的线粒体细胞器[9,44]。
3 抗菌肽在食品防腐和保鲜中的应用
抗菌肽作为天然防腐剂,安全无毒、热稳定性好、抗菌性强,只对原核生物和突变细胞起作用,而对真核生物无害,因此抗菌肽在食品的防腐和保鲜中显示出了广阔的应用前景。
羊毛硫抗生素(lantibiotics)是革兰氏阳性菌通过核糖体合成的一类抗菌肽,具有19~38个氨基酸,常含有一些非天然氨基酸,如羊毛硫氨酸、甲基羊毛硫氨酸、脱氢丙氨酸和2-氨基异丁酸,其相对分子质量小于5 000。它可抑制微生物特别是革兰氏阳性菌的生长,对革兰氏阴性菌、酵母和真菌基本无抑制作用,主要作用于细胞膜。阳离子肽nisin是由乳酸乳球菌产生的一种相对分子质量为3 500的羊毛硫胺素抗生素,属于A型羊毛硫抗生素的环形抗菌肽,能够通过破坏细菌细胞膜而杀死靶细菌,可抑制革兰氏阳性菌的生长而对食品的感官特性没有影响,保持食品的营养价值和风味特性,已成功作为天然食品防腐剂应用于鱼类等食品防腐保鲜领域中[45-46],成为传统防腐抗菌药物的良好替代品(表1)。
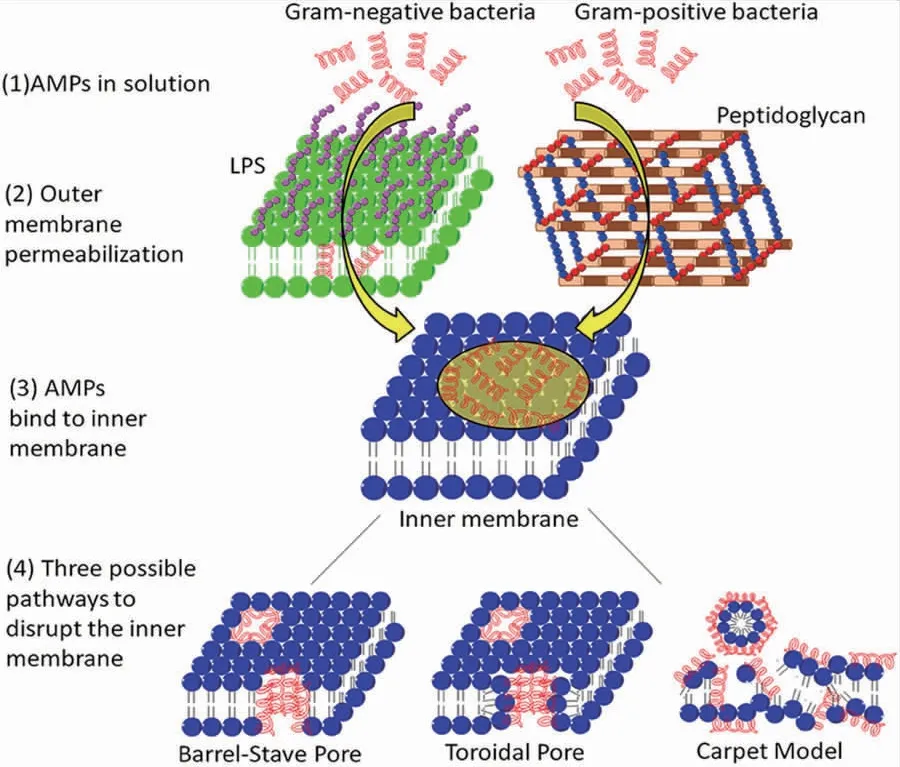
图2 抗菌肽对革兰氏阳性菌和革兰氏阴性菌的作用机制Fig.2 Interaction mechanism of AMPs to the Gram-positive and Gram-negative bacteria[39]
Nisin可抑制乳酸菌(LAB)、单核细胞增生李斯特氏菌(L.monocytogenes)、热杀索丝菌(B.thermosphacta)、蜡状芽孢杆菌(B.cereus)、肠炎沙门氏菌(Salmonella enteritidis)、大肠杆菌(E.coli)和无害李斯特菌(L.innocua)的生长,在肉和鸡蛋类食品中有良好的防腐保鲜效果。Nisin能有效防止LAB导致的肉酱香肠的腐败[47];对土耳其发酵香肠中L.monocytogenes的抑制与其浓度呈正相关[48];联合溶菌酶加入真空包装猪肉中可有效抑制B.thermosphacta和LAB的生长[49-50];与茶多酚联用,能够有效延长低温肉制品的保藏期。因为β-环糊精(β-CD)提高了nisin在胰蛋白酶中的稳定性,推迟了nisin的降解,nisin与β-CD联合应用可以有效降低熟猪肉中的细菌生长[51]。Nisin与肠肽CRL35联用,可有效控制牛肉泥中和肉类加工系统中的L.monocytogenes[52]。Nisin可延长鸡蛋的保存期。Nisin加入巴氏杀菌液体中,可降低鸡蛋中L.monocytogenes和B.cereus的数量,从而延长产品的冷藏货架期[8,53-54]。Nisin和pediocin PA-1/AcH对L.monocytogenes具有拮抗作用,在热处理下两者具有协同作用[53,55-56],在全鸡蛋和蛋清杀菌过程中增加了Salmonella enteritidis的热敏感性[56]。Nisin联合高压处理全蛋液,使得E.coli和L.innocua的数量显著降低,且这些细菌在冷藏一个月后完全变性[57]。
Nisin在米饭和面包的储藏中也显示了良好的效果,可抑制菌落的增加,从而延长米饭和面包的储藏时间[58]。Nisin由于可以抑制嗜酸耐热菌、水杨酸杆菌、嗜热脂肪芽孢杆菌和嗜热梭菌的生长,对饮料的保鲜效果也十分明显。在苹果汁和葡萄汁的保鲜中,它可抑制嗜酸耐热菌和水杨酸杆菌的生长,减缓果汁和饮料的变质[59-61]。在豆浆中,它可抑制嗜热脂肪芽孢杆菌和椰奶、椰汁中的嗜热梭菌的增殖[62]。Nisin还可使酸土脂环酸芽孢杆菌(A.acidoterrestris)内孢子失活[59-60,63]。Nisin的防腐保鲜不影响食品的风味、质地和产品结构。但是它在高温条件下性质不稳定,在60~70℃,其活性大约降低80%。它对蛋白酶K、胰蛋白酶、糜蛋白酶敏感,在pH非常小的范围内发挥生物学活性。
Nisin可用于水果的保鲜,但对酸性环境中的水果保鲜效果不明显。Nisin可抑制甜瓜片中L.monocytogenes的生长,和噬菌体联用,其活性可增强。但它对酸奶中水果的保鲜效果不明显[64]。Nisin还可用于白玉琵琶的保鲜,降低琵琶的腐烂率[65]。Nisin和bovicin HC5可抑制罐装芒果果肉中C.tyrobutyricum的生长和随后的产气[66]。Nisin还可用于蔬菜的保鲜。它减少了冷藏豆芽中L.monocytogenes的数量[67]。在罐装蔬菜中,nisin可以抑制非嗜酸杆菌 (Bacillus stearothermophilus,Clostridium the rmosaccharolyticum)和嗜酸杆菌(Clostridiumpasteurianum,Bacillusmacerans,Bacillus coagulans)导致的腐败。在加工过程中,把nisin加入到巴氏杀菌和真空包装的土豆泥中,可充分抑制细菌的生长[62]。
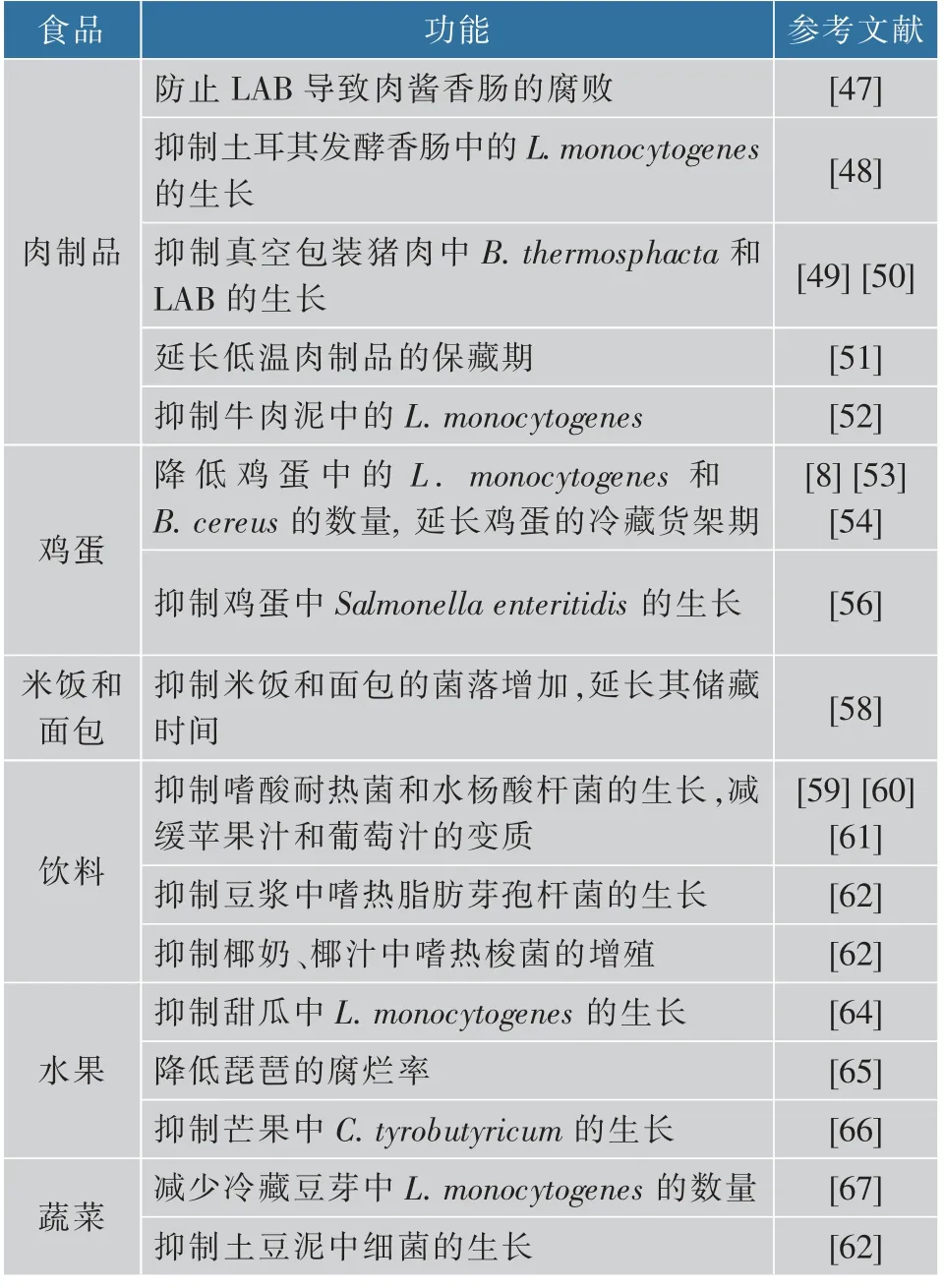
表1 Nisin在食品防腐与保鲜中的应用Table 1 Applications of nisin in food antisepsis and preservation
4 展 望
抗菌肽种类繁多,来源广泛,制备方法除了天然提取和人工合成,还可利用基因工程法中的大肠杆菌表达系统、酵母表达系统、杆状病毒-昆虫细胞表达系统、植物表达系统和动物表达系统。其主要作用机制是抗菌肽和细胞膜之间发生相互作用。关于该相互作用目前多是关于两亲性的抗菌肽在脂质体表面形成孔洞的理论研究,但是对孔洞形成机理的动力学方面,尤其在实验条件下关于这一动态过程的检测与验证研究并未报道。抗菌肽在生物体内广泛存在,具有广谱抗菌性,绿色无污染,甚至还有营养保健作用。它作为天然防腐和保鲜剂起到了良好的效果,因此被广泛应用于食品防腐和保鲜领域中。但其制备工艺复杂,成本较高,抑菌浓度相对较高,不利于大规模生产,这大大限制了其商品化进程。因此,研发抗菌肽的制备工艺,提高产量,降低成本,并研究其动态作用机制的细节,可有效推动抗菌肽进入食品防腐和保鲜领域。
