西农萨能奶山羊SLC7A5 基因克隆及其功能初步研究
2021-06-19岳子婷
陈 冲,邬 娇,岳子婷,王 瑾,潘 檀,罗 军,李 聪
(西北农林科技大学动物科技学院,陕西省遗传育种与繁殖重点实验室,陕西杨凌 712100)
山羊奶因具有较高的营养价值和独特的营养功能而越来越受到人们的关注与喜爱。首先,羊奶可以增强新生儿的免疫功能并在其生长发育过程中起重要作用,主要是羊奶中含有丰富的乳铁蛋白、免疫球蛋白等小分子活性蛋白质[1]。其次,山羊奶中的酪蛋白含有人体及畜体需要的绝大多数必需氨基酸,且氨基酸含量充足、种类齐全,并且蛋白质消化率可达98%,更有利于人体的消化吸收[2]。山羊奶在人类健康方面的生理意义越来越被重视,研究羊乳中乳蛋白的合成调控机制对其营养价值的充分发挥具有重要意义。
溶质转运蛋白家族(Solute Carrier,SLC)成员来源广泛,主要由65 个家族组成,其功能多样,可编码大约400 种功能蛋白质基因[3-4],同一家族成员间具有约25% 的序列同源性[5];溶质转运蛋白家族在细胞吸收营养物质、吸收药物和其他外源物质的生理过程中具有重要作用。实验室前期对泌乳高峰期具有高低乳蛋白含量的6 只奶山羊乳腺组织进行转录组测序,发现SLC7A5基因表达差异显著,可作为调控奶山羊乳蛋白合成的关键候选基因(未发表)。SLC7A5(LAT-1)属于APC(氨基酸多胺有机阳离子)超家族的SLC7 家族,由15 个成员组成,其中2 个是假基因,另外13 个编码蛋白分为阳离子氨基酸转运蛋白和异二聚氨基酸转运蛋白(LATs)2 个亚组[6-7]。SLC7A5(LAT-1)是一种非依赖于Na+的转运蛋白,主要和LAT2(SLC7A8)将亮氨酸、异亮氨酸、缬氨酸、苯丙氨酸、蛋氨酸、酪氨酸、组氨酸和色氨酸运输到细胞中[8],还可通过与糖蛋白SLC3A2二硫键相结合形成复合物转运亮氨酸等必需氨基酸。这些必需氨基酸不仅在蛋白质合成中发挥重要作用,而且还参与多种信号传导途径[9]。人类SLC7A5基因位于16 号染色体上,由507 个氨基酸构成,具有12 次跨膜蛋白,分子质量为55 ku,长达39 477 nt,含11 个外显子[10]。而山羊SLC7A5基因位于18 号染色体上,编码505 个氨基酸,含10 个外显子。
本研究以西农萨能奶山羊为研究对象,利用PCR技术克隆萨能奶山羊SLC7A5基因CDS 序列,并使用多款生物软件和在线工具进行生物信息学分析,同时分析其组织表达情况,为进一步研究SLC7A5基因在奶山羊乳蛋白质合成方面的调控作用提供理论依据。
1 材料与方法
1.1 材料与试剂 材料准备:在西北农林科技大学萨能羊原种场选择健康并处于泌乳高峰期的西农萨能奶山羊母羊3 只,屠宰后手术采集肝、脾、乳腺、肾、瘤胃、小肠等组织1~2 g,放置于焦碳酸二乙酯(DEPC)处理的H2O 中漂洗,除去外层薄膜及残余血迹后,迅速投入液氮保存,以用于组织总RNA 提取。试剂准备:Primer Star MAX 聚合酶、pMD19-T 载体、DNA marker、RNA 提取试剂盒、荧光定量PCR 试剂盒均购自大连宝生物工程有限公司(TaKaRa);2K plus II、大肠杆菌(E.coli)。
1.2SLC7A5基因CDS 区克隆根据NCBI 中山羊SLC7A5基因序列(登录号:XM_005691915.3),采用Primer Premier 6.0 软件设计引物。上游引物(5'→3'):GCCACCATGGCGGGCTCAGGCCCCA;下游引物(5'→3'):TTACGTCTCCTGGGGAACCACGTG。提取山羊乳腺上皮细胞总RNA,通过反转录得到的cDNA 作为模板进行PCR 扩增。PCR 反应体系:上、下游引物(10 μmol/L)各1 μL,模板(50 μmol/L)0.2 μL,Primer Star MAX酶10 μL,加灭菌水补充至20 μL。PCR 反应条件:94℃预变性5 min,98℃变性10 s,60℃退火5 s,延伸10 s。PCR 产物保存于4℃。采用1%琼脂糖凝胶电泳法对PCR 产物进行检测,切胶回收正确条带。16℃连接PMD-19T 载体12 h。连接体系:胶回收产物(DNA片段)7 μL,Solution I 5 μL,PMD-19T 载体0.5 μL。经过转化(整个操作过程在冰上进行),抽提质粒,电泳后选择阳性克隆质粒送生工生物工程(上海)股份有限公司测序。
1.3SLC7A5基因的生物信息学分析 经NCBI 中的ORF Finder(https://www.ncbi.nlm.nih.gov/orffinder/)鉴定开放阅读框;采用ProtParam(http://www.expasy.org/tools/protparam.html)在线分析SLC7A5基因编码蛋白质的基本理化性质和特征;NetPhos(http://www.dabi.temple.edu/disphos/)预测蛋白质的磷酸化位点;采用TMHMM 软件预测蛋白质跨膜结构;SignalP 4.1 Server 软件进行蛋白质信号肽分析;TargetP 1.1 Server软件预测蛋白质亚细胞定位;以NPS(https://npsaprabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_gor4.html)预测蛋白质二级结构,经Phyre2(http://bioinf.cs.ucl.ac.uk./psipred/)程序分析预测蛋白质三级结构;将SLC7A5基因的DNA 序列和氨基酸序列在NCBI 上进行Blast 同源比对,采用MEGA7.0 构建系统进化树;利用STRING(http://www.string-db.org/)交互式数据库进行蛋白质互作分析。
1.4SLC7A5基因组织表达谱分析 分别提取3 只萨能奶山羊肝、脾、乳腺、肾、瘤胃、小肠6 个组织的总RNA,以及处于不同泌乳时期乳腺组织的总RNA。在TaKaRa 反转录试剂盒的操作说明下,将RNA 反转录合成为cDNA。反转录采用两步法进行,操作过程要在冰上进行。以反转录后的cDNA 作为模板,通过实时荧光定量PCR 检测其相对表达量,每只个体重复2 次,每次3 个重复。内参基因为UXT和MRPS9,定量引物SLC7A5(片段全长286 bp)为F:CTCACTGCCGTG AACTGCTA;R:CCAGGGGCAGGTTTCTGTAG;进行qRT-PCR,反应体系15 μL:SYBR®Premix EX TaqTM(2×)7.5 μL,模板cDNA(1 000 μg/μL)1 μL,上下游引物(10 μmol/L)各0.7 μL,ddH2O 5.1 μL。SLC7A5基因的组织相对表达量采用2-ΔΔCt法计算。采用GraphPad Prism v7.0.4 进行绘图。
1.5 统计分析 利用软件SPSS 17.0 进行One-Way ANOVA显著性检验,实验结果均以平均值± 标准差表示。当P<0.05 时,统计学差异显著;当P<0.01 时,统计学差异极显著。
2 结果
2.1SLC7A5基因CDS 区克隆及测序 克隆后经过1%琼脂糖凝胶检测,扩增条带如图1 所示,全长约为1 518 bp,经与NCBI 上传序列比对,可以确定为山羊CDS 区序列。碱基含量分别为:A=15.8%,T=21.2%,C=34.1%,G=28.9%。

图1 SLC7A5 基因扩增产物琼脂糖凝胶电泳图
2.2SLC7A5基因生物信息学分析
2.2.1 SLC7A5 蛋白质理化性质分析 西农萨能奶山羊SLC7A5 蛋白质含有氨基酸505 个;氨基酸分子量55.01 ku;理论等电点为8.18;总分子式为C2573H4022N616O670S22。分析得到不稳定系数为37.74,为稳定蛋白。其中亮氨酸(Leu)、缬氨酸(Val)和丙氨酸(Ala)含量占比相对较高,分别占14.1%、9.7%、9.5%。组氨酸(His)和天冬氨酸(Asp)含量占比相对较低,分别占1.0%和1.6%。带负电残基总数天冬氨酸(Asp)+谷氨酸(Glu)为31 个,带正电残基总数精氨酸(Arg)+赖氨酸(Lys)为34 个。推测SLC7A5 为碱性蛋白质。
2.2.2 氨基酸序列及磷酸位点分析 通过DISPHOS1.3分析西农萨能奶山羊SLC7A5 蛋白的氨基酸组成,结果见图2。该蛋白共含有2 个丝氨酸(Ser)磷酸化位点,1 个苏氨酸(Thr)磷酸化位点,并在115 位有1 个酪氨酸(Tyr)磷酸化位点。
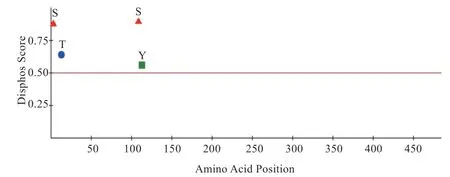
图2 SLC7A5 基因磷酸化位点图
2.2.3 SLC7A5 蛋白的亚细胞定位、跨膜结构及结构域分析 SignalP 4.1 Server 分析显示,SLC7A5 蛋白为无信号肽蛋白,其所分泌信号肽的概率低至0.003 2。TargetP 1.1 Server 预测SLC7A5 蛋白为膜蛋白,主要分布于质膜(69.6%)、内质网(17.4%)、液泡(8.7%)。通过TMHMM 预测跨膜结构域,结果显示SLC7A5 是具有11 段跨膜螺旋的跨膜蛋白。
2.2.4 SLC7A5 蛋白质二级结构、三级结构预测 在线蛋白分析系统分析显示,西农萨能奶山羊SLC7A5 蛋白质α螺旋可能由141 个氨基酸形成,延伸链可能由124 个氨基酸形成,无规则卷曲可能由240 个氨基酸形成,三者氨基酸含量占比分别为27.92%、24.55%、47.52%。SLC7A5 蛋白质的二级结构、三级结构分别如图3 和图4 所示。

图3 SLC7A5 蛋白质二级结构预测图

图4 SLC7A5 蛋白质三级结构预测图
2.2.5 SLC7A5 蛋白相互作用预测 经分析得到SLC7A5蛋白相互作用预测结果如图5 所示,SLC7A5 与SLC3A2、SLC38A2、SLC1A5、SLC1A6 等10 种蛋白存在互作关系。

图5 SLC7A5 蛋白质相互作用预测图
2.2.6 SLC7A5 蛋白系统发育树 将克隆的SLC7A5基因转换成氨基酸序列,并在NCBI 上根据同源性查找山羊、绵羊、牛、水牛、小鼠、人、大猩猩、美洲野牛8种氨基酸序列,通过MEGA7.0 得到蛋白系统发育树。如图6 所示,奶山羊与山羊、绵羊和牛的亲缘关系最近,分别为100%、99.60%、99.01%。

图6 SLC7A5 蛋白系统发育树
2.3SLC7A5基因组织表达谱分析 如图7 所示,SLC7A5基因以肝脏为对照组,其在乳腺组织中表达量最高(P<0.05),其次是脾脏(P<0.05),在小肠和肾脏中表达含量较低(P>0.05)。对SLC7A5基因在不同泌乳时期乳腺组织表达谱分析(图8)发现,以干奶期为对照组,SLC7A5 在泌乳后期表达量最高(P<0.05),其次是前期(P<0.05),泌乳盛期差异不显著。

图7 SLC7A5 在不同组织中相对表达量
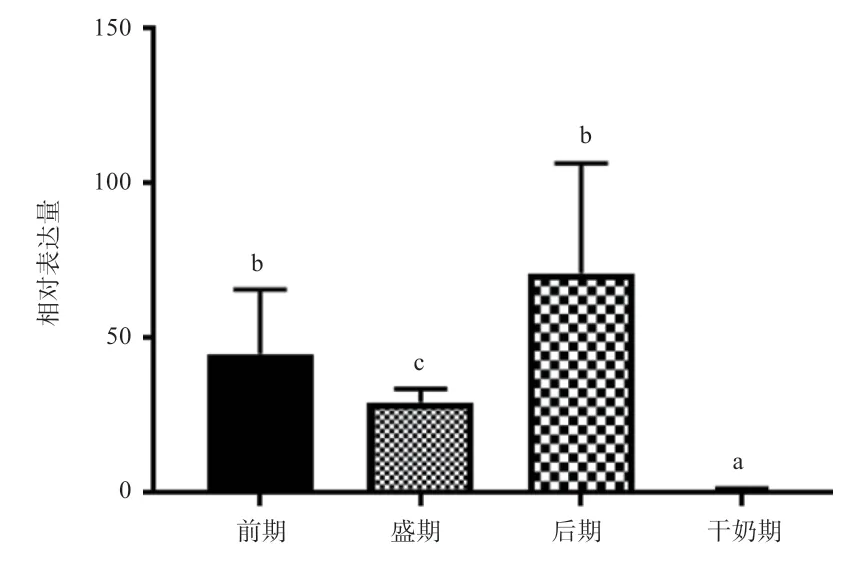
图8 SLC7A5 在不同泌乳时期乳腺组织的相对表达量
3 讨 论
SLC7A5 是一种钠非依赖性反转运蛋白,转运必需氨基酸参与机体活动[8]。本研究通过克隆所获得的CDS 序列经序列比对、蛋白结构分析显示,其编码蛋白为无信号肽、具有跨膜结构域的稳定碱性蛋白质,主要分布于质膜。Nakamura 等[11]通过克隆小鼠4F2 L-链 SLC7A5 cDNA,推导的氨基酸序列表明4F2 L-链至少有11 个或12 个螺旋跨膜结构域,计算分子量为56 ku,并发现4F2 L-链与最近报道的大鼠LAT1 高度同源(98% 同源性),为哺乳动物细胞中特定的氨基酸转运蛋白到质膜的“引导分子”。有研究表明,通过对高、低乳蛋白奶牛乳腺组织切片进行免疫荧光染色,发现LAT1 位于乳腺上皮细胞的整个质膜上,进一步揭示LAT1 为乳腺提供了必需氨基酸[12]。在乳蛋白含量较高的奶牛乳腺上皮细胞中,LAT1 表达下调可降低细胞活力和β-酪蛋白表达,而过表达则相反[13]。本研究发现SLC7A5在乳腺中的表达量最高,其次是脾脏组织,各个组织中的表达存在显著差异,在乳腺组织的不同泌乳时期,泌乳后期表达量最高,其次是前期、盛期及干奶期。在乳腺组织的高表达表明SLC7A5的生物学功能很大程度在奶山羊乳腺组织中发挥作用。研究表明,SLC7A5在啮齿类动物乳蛋白合成中的表达增加[14]。LAT1 参与泌乳奶牛乳腺中乳蛋白的合成,mTORC1 信号通路可能是调控LAT1 表达的一个控制点,最终可以用来改变乳蛋白的合成[13]。通过SLC3A2/SLC7A5异二聚体将必需氨基酸转运到细胞中被认为是限制EAA激活mTORC1 的速率限制因素,即利用细胞内L-谷氨酰胺作为外排底物来调节细胞外L-亮氨酸的摄取,从而导致mTORC1 的激活[15]。SLC7A5基因在奶山羊小肠、肾脏中的相对表达较低甚至不表达,Nakamura 等[11]在小鼠的肠和肾中几乎没有检测到4F2 L-链。而本实验中SLC7A5在泌乳后期相对表达量高,具体调节机制需要进一步研究。
本研究发现SLC7A5 蛋白质与SLC3A2、SLC38A2、SLC1A1、SLC1A2、SLC1A4、SLC1A5、SLC1A6、SLC1A7、C1H3orf58、BSG(肿瘤细胞胶原酶激活因子)10 种蛋白存在互作关系。SLC7A5 蛋白与SLC3A2 蛋白的互作关系进一步印证了SLC7A5基因通过与SLC3A2形成二聚体参与mTORC1 过程,通过转运机体所需的必需氨基酸进而调节乳腺乳蛋白的合成[15]。SLC1 家族(高亲和力的谷氨酸和中性氨基酸转运体)主要参与神经兴奋的终止和谷氨酸代谢,其中高亲和力谷氨酸转运体EAAC1(SLC1A1)、GLT1(SLC1A2)、EAAT4(SLC1A6)和EAAT5(SLC1A7)通过3 个Na+和1个H+的共转运和1 个K+的反转运介导细胞摄取谷氨酸盐。因此,它们保护中枢神经系统免受谷氨酸引起的神经毒性。SLC1 家族还包括2 个中性氨基酸转运蛋白ASCT1(SLC1A4)和ASCT2(SLC1A5),这2 种 转运蛋白促进神经元和或外周组织细胞中氨基酸的电中性交换[16]。研究表明SLC7A5主要参与血脑屏障,从而介导营养物质和药物进入中枢神经系统(CNS)[17];并且SLC7A5对维持大脑中支链氨基酸(BCAAs)的正常水平至关重要,大脑中支链氨基酸(BCAAs)的失衡可能导致神经病态,如自闭症谱系障碍[18]。研究发现,SLC7A5通常与4F2hc/LAT1(SLC3A2/SLC7A5)一起表达,后者是一种能交换大量中性氨基酸的异源反转运体,对于LAT1 的稳定性和定位于血浆膜至关重要。在多项研究中,这2 种转运蛋白都与肿瘤生长和mTOR信号传导有关[19-20],具体地说,LAT-1 输出的谷氨酰胺通过ASCT2 向细胞输入额外的氨基酸营养素(如亮氨酸),后者通过雷帕霉素(mTOR)途径的哺乳动物靶点控制细胞增殖。这2 种交换物在不同类型的癌症如三阴性乳腺癌、前列腺癌、多形性胶质母细胞瘤(GBM)中上调,已成为肿瘤治疗的理想靶点[21]。BGS 作为一种多功能蛋白分子,在炎症反应、胚胎发育、肿瘤的浸润和转移等多种病理生理过程发挥重要作用[22]。而研究发现SLC7A5启动子具有癌基因c-Myc 的典型结合位点,癌基因的过度表达导致SLC7A5的表达增加[21]。目前对SLC7A5基因的研究主要集中在人类疾病方面,值得注意的是,氨基酸转运体的功能紊乱与多种疾病有关,这表明许多溶质转运蛋白在人类疾病中起到潜在靶点作用,随着研究的深入,在输尿管癌、淋巴癌、结肠直肠癌等人类恶性肿瘤中均已发现SLC7A5表达异常升高的现象[23]。
4 结 论
本研究通过克隆得到西农萨能奶山羊SLC7A5基因CDS 区,全长1 518 bp,编码505 个氨基酸,蛋白分子量为55.01 ku。SLC7A5 蛋白质亚细胞定位主要在质膜上,为无信号肽、具有跨膜结构域的稳定碱性蛋白质,有4 个磷酸化位点;SLC7A5 与SLC1A5、SLC3A2 等10 种蛋白存在相互作用;与山羊、绵羊和牛的亲缘关系最近。SLC7A5基因在奶山羊乳腺组织和泌乳后期高表达。本研究结果为进一步研究SLC7A5基因在奶山羊乳蛋白质合成方面的调控作用提供理论依据。
