石羊河下游人工梭梭水分利用策略季节变化
2021-06-18张文龙朱淑娟柴成武姜生秀耿冬梅
张文龙,赵 鹏,朱淑娟,柴成武,姜生秀,耿冬梅,张 富
(1.甘肃农业大学林学院,甘肃兰州730010;2.甘肃省水利水电勘测设计研究院有限责任公司,甘肃兰州730010;3.甘肃省治沙研究所/甘肃民勤荒漠草地生态系统国家野外科学观测研究站,甘肃民勤733000)
水分是干旱地区植物生长和繁殖的主要限制性因素。植物可以利用的水分主要来自地表径流、土壤水、大气降水及地下水,其中土壤水主要来源于降水与地下水。植物从外界吸收水分通过根茎导管运输,一部分用于光合和呼吸作用,同时通过叶片的蒸腾作用向大气中散失水汽,水分代谢处于动态平衡之中。一般认为,除少数泌盐植物外,在陆地生态系统中植物根系吸收的水分在到达叶片或幼嫩未栓化的枝条等器官之前未发生同位素分馏[1-4]。降水氢氧稳定同位素的时空差异会导致土壤水、地表水、地下水及植物水的时空差异。土壤水与降水、地下水稳定氢氧同位素之间显著的差异也是量化植物水分来源的重要前提[5-6]。因为土壤水输入存在季节性变化,另外由于地表的蒸发和地下水的补给,所以造成了土壤水氢氧稳定同位素垂直分布梯度的形成。植物木质部水的稳定同位素代表了潜在水源的混合值。盐生、旱生植物的根部内外皮层细胞壁发达,利用共质体途径运输水分造成根部δD富集,导致木质部水δD不能完全反映土壤水的δD。因此,一般选择δ18O来研究植物水分来源[7]。氢氧稳定同位素由于破坏性小、精度高的特点,已用于植物水分来源识别技术[8-10],一方面可确定植物根系吸收水分最活跃的区域[11],另一方面可有效量化植物对不同水源的选择性利用。荒漠生态系统中长期稳定水源是影响植物生存的重要因素,可选择性利用不同水源的物种一般具有较强的竞争力。因此,研究荒漠生态系统植物的水分来源,有助于更好理解土壤-植物间的相互作用及物种共存机制[12]。
梭梭(Haloxylon ammodendron)属藜科梭梭属小乔木,抗旱、耐寒、耐盐碱、耐土壤贫瘠、抗风蚀沙埋,具有极强的生态适应能力,已成为干旱区防风固沙林的首选造林树种。从20世纪50年代末开始,河西走廊沿线绿洲外围开始了大规模的以梭梭耐旱灌木为主的固沙林营造活动。截至2016年,甘肃河西走廊人工梭梭林总面积达8.9×104hm2,其中民勤县人工梭梭林发展时间最早、造林规模最大,保存面积达到4.35×104hm2,在绿洲边缘构筑了2~5 km的梭梭防风固沙带,对改善小气候、改良土壤、保护生物多样性和增加碳汇起到关键性作用。从20世纪70年代开始,由于气候干旱、地下水位下降、造林密度过大等原因,部分区域人工梭梭林出现了大面积的衰退甚至死亡,严重威胁到绿洲的生态安全,亟需开展恢复重建工作。已有研究主要集中在梭梭衰亡原因[13]、恢复措施[14]、群落与水土因子的关系[15]、自然稀疏过程[16]、光合生理特性与形态[17]、群落特征[18]、净碳交换及其影响因子[19]、人工梭梭林自然更新的土壤条件[20]、土壤种子库[21]、立地条件[22]、更新苗的空间分布特征[23]、土壤种子库动态及其与地上植被的关系[24]等方面。目前,关于民勤绿洲边缘人工梭梭水分利用策略季节变化的研究尚未见报道。为此,本研究基于氢氧稳定同位素技术,通过对降水、地下水、土壤水及木质部水的季节观测,运用IsoSource模型研究了不同季节人工梭梭的水分利用策略,研究成果对退化人工梭梭林的恢复与重建具有重要的理论指导意义。
1 材料与方法
1.1 研究区概况
据甘肃民勤荒漠草地生态系统国家野外科学观测研究站长期观测资料,研究区属于典型的大陆性荒漠气候,多年平均降水量110 mm,多集中在7月、8月、9月,占全年总降水量的57%。年均气温7.4℃,年均地表温度为11.6℃,全年无霜期175 d,相对湿度为57%,≥10℃积温3 248.8 ℃·d,日照时数2 410.4 h,平均气压866 hPa,最大冻土深度103 cm。年蒸发量1 583.3 mm,7月蒸发量最大,为263.4 mm,1月蒸发量最小,为29 mm。主导风向为西北风,年平均风速0.7 m·s−1,5月最大风速13 m·s−1,年均沙尘暴日数6 d,年均扬沙日数20 d,浮尘日数23 d。民勤属于干旱温带荒漠灰棕漠土地带,地带性土壤为灰棕漠土,研究区土壤类型为风沙土。
1.2 试验设计
选择甘肃民勤治沙综合试验站40年林龄人工梭梭为研究对象,于2016年4月−10月下旬在样地选取3株长势一致的梭梭(表1),剪取非绿色栓化小枝(长3~5 cm,直径为0.1~0.3 cm),将皮去掉后装入8 mL采样瓶,封口膜密封,恒温箱冷藏,带回实验室−18℃冷冻保存。在距离梭梭植株主干100 cm处的根际土壤区域,用土钻从表层开始,每20 cm为一层进行取样,直到200 cm。所取土样一部分装入铝盒内,用于测定土壤含水量;另一部分装入8 mL采样瓶,用封口膜密封,放入恒温箱冷藏,带回实验室冷冻保存,用于测定土壤水分的稳定氢氧同位素值。根据各土层土壤水δ18O值分别将土壤水划分为表层0−20 cm、浅层20− 60 cm、中层60−120 cm、深层120−200 cm不同潜在水源。研究区地下水水位埋深为27.5 m,地下水样品采自附近农田灌溉井水。降水样品采用雨量器收集,记录降水日期及降水量,封口膜密封,4℃冰箱保存。
1.3 指标测定及方法
含水量测定采用烘干称重法[25],土壤含水量计算公式:土壤含水量=(土壤湿重− 土壤干重)/土壤干重×100%。
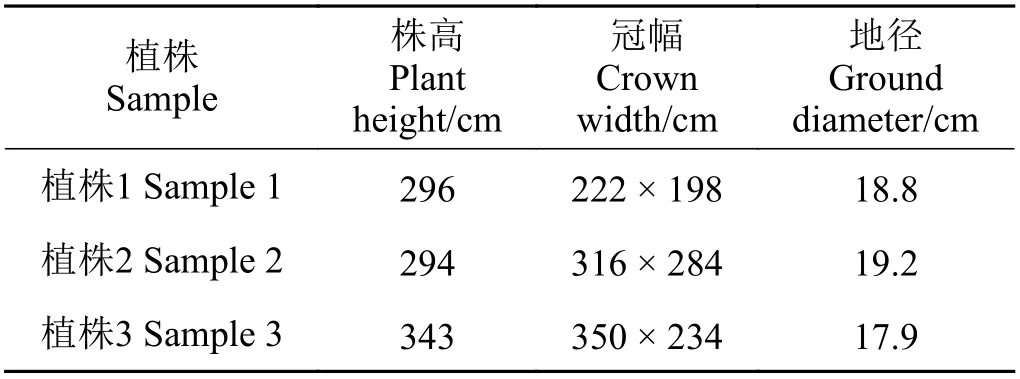
表1 梭梭植株生物性状Table 1 Biocharacteristicsof artificial Haloxylon ammodendron
样品分析测试在甘肃省荒漠化与风沙灾害防治国家重点实验室培育基地碳水同位素实验室完成。采用低温真空抽提系统提取土壤水和植物水。将抽提收集的水分装入2 mL样品小瓶,封口膜封口,置于2℃冰箱中保存。氢氧稳定同位素利用Picarro L2120-i水同位素分析仪测定,用相对于维也纳标准平均海水(VSMOW)的千分值偏差表示:

式中:R样品和R标准分别为样品和国际通用标准物中元素的轻重同位素之比(18O/16O)。测试精度为0.5‰(δD)、0.1‰(δ18O)。
采用Orgin 8.0软件的Plot功能对数据进行分析作图。运用同位素混合模型(Isotope mixing model)计算植物对各潜在水源的可能利用比例。该模型以同位素质量平衡的多元线性混合方程为基础,用于计算多个水源时植物对各水源的利用比例[26]。在初步判定两种植物的水分来源后,将植物木质部水和各潜在水源的δ18O值代入模型,构建质量平衡公式:

式中:δA、δB、δC、δD为各水源的δ18O值,δM为植物木质部水的δ18O值,FA、FB、FC、FD为各潜在水源所占比例。模型为植物的每个水源的利用比例赋值,比例增加的步长视为来源增量,如1%或2%,各水源被利用比例的组合总计为100%(式2),利用迭代方法计算可能的比例组合。当各水源的δ18O值加权值之和等于植物木质部水的δ18O值(式1),或小于某一可接受范围(质量平衡公差)时,视为符合条件的比例组合。模型运行结果中,植物对各水源的利用比例是一个范围(最小值–最大值)。本研究中,模型的来源增量设为1%,质量平衡公差设为0.1‰。
2 结果与分析
2.1 土壤水及其δ18O的季节变化
人工梭梭根际土壤水分与降水及其蒸腾耗水的季节变化密切相关。由于地下水埋深已达20 m以下,大气降水是石羊河下游荒漠区土壤重要的补给源。2015年11月至2016年3月共降水32.6 mm,前期土壤水分补给充足。石羊河下游人工梭梭一般在4月中旬开始萌发,萌发阶段水分消耗较少。4月和5月气温较低,蒸发损失小,造成人工梭梭根际0−200 cm深度土层平均土壤水分含量分别为1.05%和1.22%,显著高于6月(0.83%)、7月(0.80%)、10月(0.67%)、9月(0.65%)。虽然7月降水量达26.1 mm,由于长期高温引起的较大的蒸散量,降水对梭梭根际土壤水分补给效应不明显(图1)。

图1 2016年人工梭梭根际土壤水分与降水的关系Figure 1 Relationships between soil moisture and rainfall in the rhizosphere of Haloxylon ammodendron in 2016
人工梭梭根际土壤水分的垂直分布(图2)显示,由于春季地表蒸散相对较小,0− 60 cm表层土壤水分随深度增大逐渐减小,60−120 cm土壤水分随深度增加略为增大,春季土壤水分显著高于夏季和秋季。夏季降水多,温度高,是梭梭蒸腾耗水最活跃的季节,0−100 cm土壤水分从0.35%增加至0.83%。在100 − 140 cm出现一个峰值,即在120 cm深度达到最大值(1.13%)。秋季温度下降,0−40 cm土壤水分逐渐增大至0.56%,同样在100−140 cm出现一个峰值,即在120 cm深度达到最大值(0.91%)。春、夏、秋季梭梭根际120 cm深度处土壤水分峰值的出现可能与其根系的水分再分配功能有关。即在夜间弱蒸腾条件下,梭梭根系吸收下层湿润土壤中的水分并释放至上层干燥土壤中,以缓解干旱环境下植物的水分亏缺,水分再分配作用可促进植物蒸腾及光合作用,增加植物的根冠比和根系碳分配[27]。
梭梭根际土壤水δ18O的垂直变化(图2)表明,土壤水δ18O春季在−5.61‰~4.71‰,平均值为−1.624‰。夏季在−14.84‰~−1.34‰,平均值为−5.003‰。秋季在−8.34‰~1.24‰,平均值为−2.103‰。春、夏、秋生长季梭梭根际垂直0−200 cm土层间土壤水δ18O最大相差10.32‰。研究区土壤水靠降水的入渗补给,降水进入土壤向下入渗的过程中,与土壤自由水发生不同程度的混合。同时,表层土壤在蒸发的过程中受到动力分馏后形成δ18O最大富集层。降水少时多以活塞流下渗,降水大时较易形成优先流补给深层土壤水,入渗方式决定着土壤水稳定氢氧同位素垂直分布格局。不同土层间土壤水δ18O差异明显,0−60 cm表层富集效应明显,变化剧烈,中下层逐渐减小且相对稳定。
2.2 水分来源判定
基于氢氧同位素特征的分析,可以判断出不同水体间相互转化关系。降水氢氧同位素变化:δD为−152.7‰~54.237‰,变幅为201.69‰;δ18O为−15.60‰~8.75‰,变幅为24.35‰(图3)。当地大气降水线斜率和截距分别为7.29、−1.81,均小于全球降水线的斜率和截距。土壤水氢氧同位素值主要分布在当地大气水线的右下方,较降水相对富集,δD为−134.92‰~1.93‰,变幅为136.84‰;δ18O为−18.39‰~13.17‰,变幅为31.57‰。土壤水δD的变幅小于降水,δ18O的变幅大于降水。土壤水蒸发线斜率为3.66,截距为−48.7,远低于当地大气水线,意味着降水入渗过程中发生了蒸发分馏。地下水氢氧同位素变化范围:δD为−64.06‰~−58.04‰,变幅为6.02‰;δ18O为−9.90‰~−8.83‰,变幅为1.07‰。梭梭木质部水氢氧同位素变化:δD为−91.52‰~−55.94‰,变幅为35.58‰;δ18O为−10.04‰~−3.19‰,变幅为6.85‰。梭梭木质部水δ18O与土壤水交叉分布在大气水线以下,表明土壤水是梭梭的潜在水源。
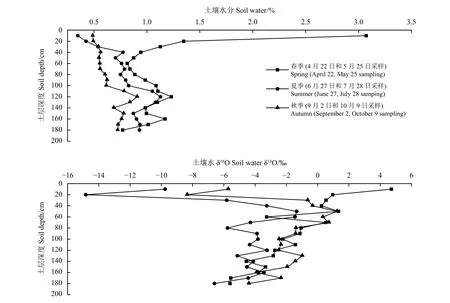
图2 人工梭梭根际土壤水分(δ18O)时空变化Figure2 Temporal and spatial variation of soil moisture (δ18O)in the rhizosphere of Haloxylon ammodendron
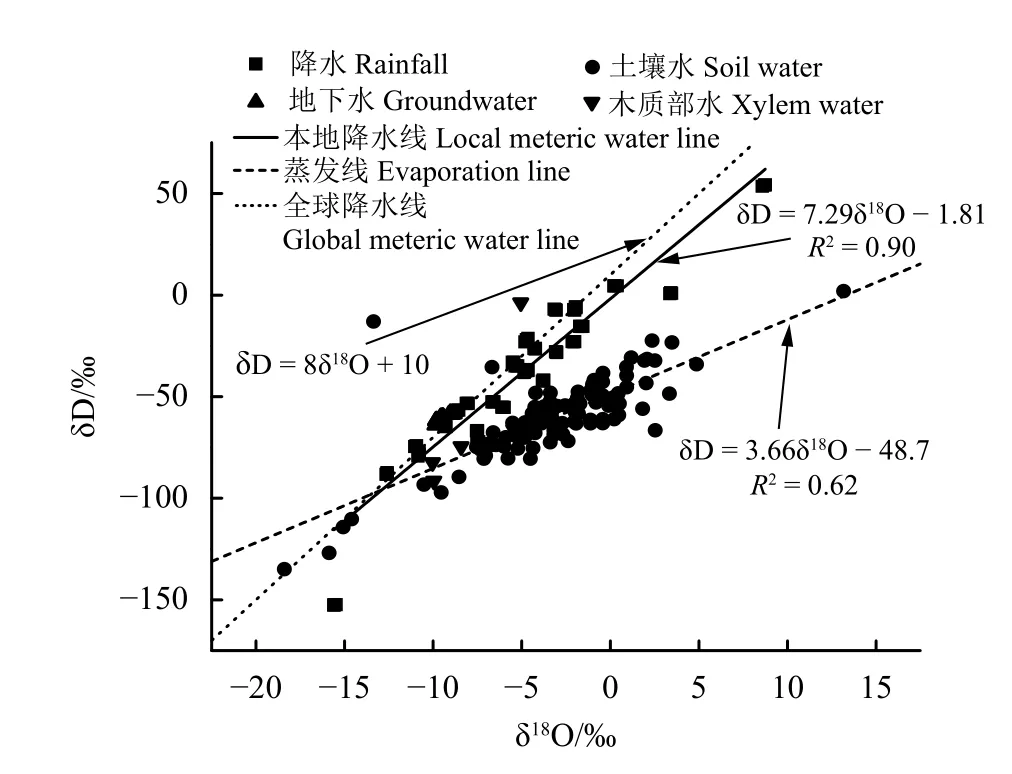
图3 降水、土壤水、地下水、梭梭木质部水δD与δ18O的线性回归关系Figure 3 Linear regression relationship betweenδD and δ18O in rainfall,soil moisture,groundwater,and xylem of Haloxylon ammodendron
2.3 水分来源量化
采用直接比较法可以直观地判断出植物的吸水部位。4月梭梭木质部水δ18O与土壤水δ18O在50、70、160 cm处有3个交点,证明4月梭梭吸收利用土壤水的深度分别为50、70、160 cm(图4)。5月梭梭木质部水δ18O与土壤水δ18O在75、85、100、115、145、160 cm处有6个交点,证明5月梭梭利用土壤水的深度分别为75、85、100、115、145、160 cm。6月梭梭木质部水δ18O与土壤水δ18O在25 cm处有1个交点,证明6月梭梭利用土壤水的深度为25 cm。7月梭梭木质部水δ18O与土壤水δ18O在15、45、90、115、135、155和175 cm处有7个交点,证明7月梭梭利用土壤水的深度为15~175 cm。9月梭梭木质部水δ18O与土壤水δ18O在175 cm处有1个交点,证明9月梭梭利用土壤水的深度为175 cm。10月梭梭土壤水δ18O与木质部水δ18O在15、25 cm处有2个交点,证明10月梭梭利用土壤水的深度为15、25 cm。
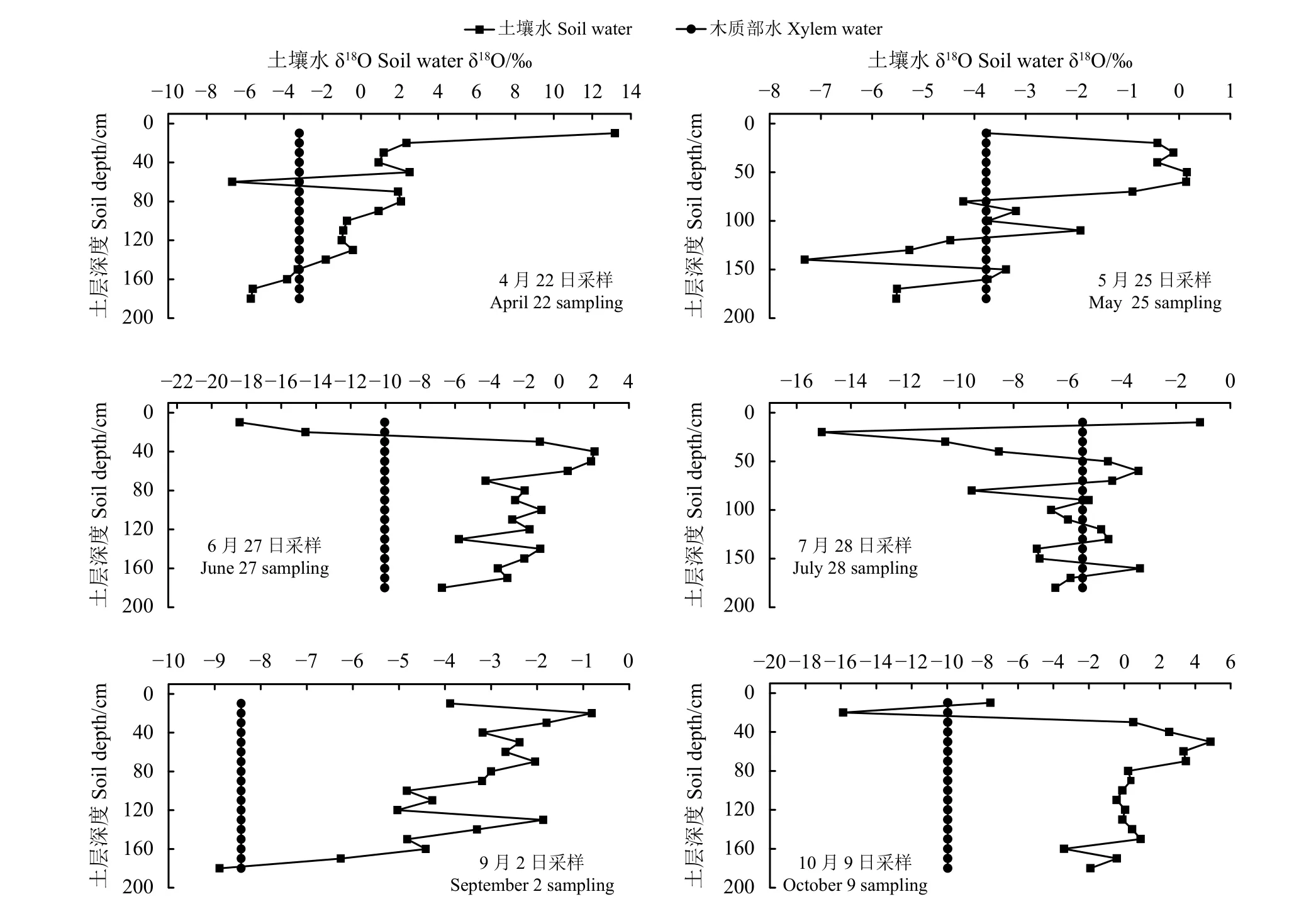
图4 研究区4月−10月梭梭茎干水与土壤水氧同位素关系Figure 4 Relationship between δ18O of xylem water and soil water from April to October
根据各土层土壤水δ18O值分别将土壤水划分为表层0− 20 cm、浅层20− 60 cm、中层60−120 cm、深层120−200 cm不同潜在水源。对梭梭而言,4月主要利用20−60、120− 200 cm土壤水,分别占31.8%和36.2% (图5和表2)。5月主要利用120 − 200、60− 120、0 − 20 cm土壤水,利用比例为54.1%、22.3%和14.9%。6月对0 − 20 cm、120 − 200 cm、60 − 120 cm土壤水利用比例分别为55.5%、16.9%和15.3%。7月对60− 120、20− 60、120−200 cm土壤水利用比例分别为34.2%、31.9%和24.5%。9月对120 −200、60−120、0− 20 cm土壤水利用比例分别为92.1%、3.3%和2.3%。10月对0 − 20、120 − 200 m、60 − 120 cm土壤水利用比例分别为85.9%、5.4%和4.7%。
3 讨论
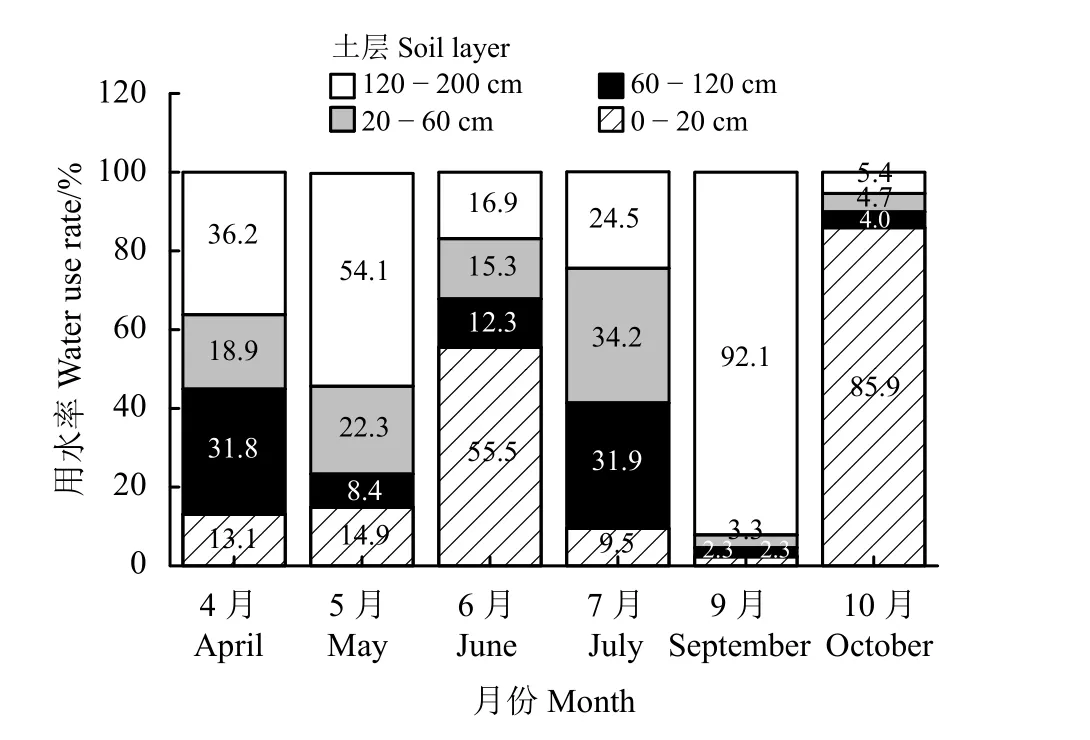
图5 不同季节梭梭潜在水源的贡献比Figure 5 Contributions of potential water sources of Haloxylon ammodendron in different seasons
降水格局在一定程度上决定了植物的水分来源和水分利用策略,进而影响生态系统中的群落结构[28]。柠条(Caragana korshinskii)和沙柳(Salix psammophila)在旱季均主要利用深层土壤水分,雨季更倾向于利用浅层土壤水分[29]。由于降水偏少,小叶锦鸡儿(Caragana microphylla)对浅层水分的利用急剧减少,转而利用更深层次的土壤水分[30]。沙质生境下泡泡刺(Nitraria sphaerocarpa)在降水较少的春季和秋季主要利用地下水,其贡献率可达50%以上;而砾质生境下的泡泡刺无法利用深达11.5 m地下水,其水分来源受降水控制,具有较大的季节变异性。两种生境下的泡泡刺对降水响应显著[31]。准格尔盆地南缘雨前梭梭水分利用来源主要为地下水(贡献率34.14%)和120−180 cm土层土壤水(贡献率29.87%),雨后梭梭水分利用来源主要为60−120 cm土层土壤水(贡献率33.19%)和地下水(贡献率30.67%)[32]。梭梭在古尔班通古特沙漠能够存活主要是因为吸收了降水形成的浅层土壤水分[33]。本研究春季降水偏少,梭梭对深层土壤水的利用比例达到65.6%,夏季和秋季降水多,梭梭对浅层土壤水分的利用比例分别占33.65%、44%,体现了降水格局对梭梭季节水分来源的重要影响。
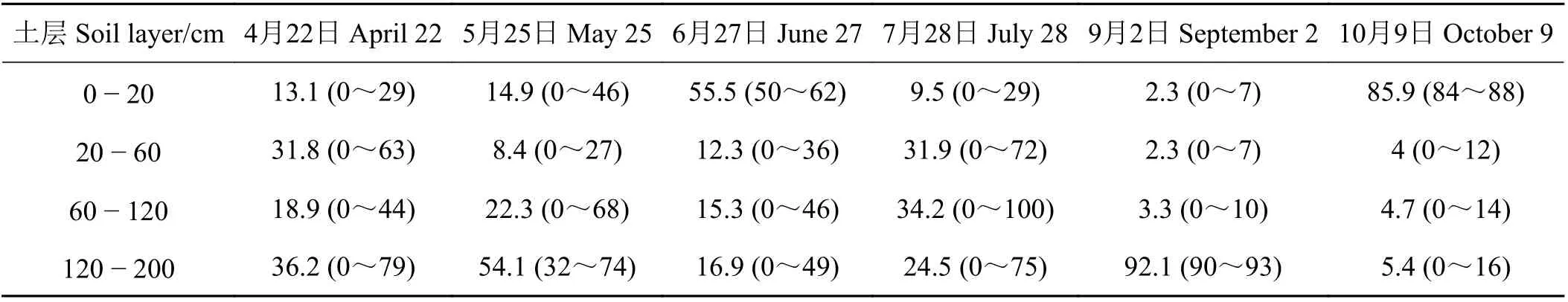
表2 不同月份梭梭对潜在水源的利用比例Table2 Contributionsof soil water to Haloxylon ammodendron in different months%
应对环境水分胁迫与改变,荒漠植物具有高效的自我协调与适应能力,不同水平调节适应机制之间存在内在协调性[34]。与白刺(Nitraria tangutorum)、沙拐枣(Calligonum mongolicum)等只利用中浅层土壤水分的植物相比,临泽绿洲荒漠过渡带地下水湿生植物梭梭可以利用多个潜在水源,对干旱胁迫的适应能力更强[35]。梭梭根系具有二态性,在古尔班通古特沙漠5月主要利用浅层(20−40 cm)土壤水、深层(100−350 cm)土壤水及地下水,8月主要利用深层(160−350 cm)土壤水及地下水[36]。临泽荒漠绿洲过渡带5年林龄梭梭主要利用浅层土壤水。10年林龄梭梭的水分来源取决于季节土壤水分条件。尽管垂直根系可以吸收利用深层土壤水,但人工梭梭超过50%的水分来源于中浅层土壤水分。20和40年林龄梭梭利用稳定的地下水。幼龄级梭梭利用降水补给的土壤水分,当根系深度足够深时,旱季水分来源于深层土壤水。大于20年林龄梭梭主要利用深层稳定水源[37]。梭梭垂直根系深达13.5 m,水平根系主要分布在50−100 cm土层[38],根系对水资源的再分配功能是其适应严酷环境的重要因素之一[39]。二态根系对不同深度土壤水分的选择性吸收利用是梭梭应对干旱胁迫的重要对策。此外,最新研究表明高湿度大气条件下,梭梭同化枝能够吸收大气水运输至次级枝或茎干储存,改变植株水势、气孔导度、蒸腾速率、瞬时水分利用效率等气体交换特性[40]。由此可见,不饱和水汽也是梭梭的潜在水分来源,未来应加强荒漠植物利用隐匿降水的研究。
4 结论
本研究发现人工梭梭根际土壤水分均值为0.87%,旱化严重,时空特征明显。春季主要利用120−200 cm深层土壤水分;由于降雨较多,夏季对0−20 cm表层土壤水分的利用比例最大为32.5%;秋季主要对0−20 cm表层、120−200 cm深层土壤水分利用较多,利用比例分别为44.4%、48.75%。由此可见,不同深度土壤水分是人工梭梭重要的季节水分来源,水分来源随土壤水分的季节变化而调整以适应干旱生境。建议在日常抚育管理中可采取人工辅助降水入渗的方法来维持林地土壤水分平衡。
