Shifts of sediment bacterial community and respiration along a successional gradient in a typical karst plateau lake wetland (China)*
2021-06-15PinhuaXIAJianZHANGJinboLIULifeiYU
Pinhua XIA , Jian ZHANG , Jinbo LIU , Lifei YU,**
1 Guizhou Key Laboratory for Mountainous Environmental Information and Ecological Protection, Guizhou Normal University,Guiyang 550001, China
2 College of Life Science, Guizhou University, Guiyang 550025, China
3 Collaborative Innovation Center for Mountain Ecology& Agro-Bioengineering, Guizhou University, Guiyang 550025, China
4 Department of Hepatobiliary Surgery, the Affi liated Hospital of Southwest Medical University, Luzhou 646000, China
Abstract Bacteria are important regulators of carbon cycling in lakes and are central to sediment ecosystem processes. However, the sediment microbial communities and their respiratory responses to the lake wetland succession are poorly understood. In this study, we collected sediment samples from four diff erent succession points (the Potamogeton lucens zone, the Scirpus tabernaemontani zone, the Scirpus triqueter zone, and the Juncus eff usus zone) in the Caohai Wetland of the Guizhou Plateau (China).The bacterial communities at these succession points were studied using a high-throughput sequencing approach. The sediment microbial respiration (SR) was measured using static chambers in the field and basal respiration (BR) was determined in the laboratory. The results show that the dominant bacterial taxa in the sediment was Proteobacteria (34.7%), Chloroflexi (17.8%), Bacteroidetes (7.3%), Acidobacteria (6.6%),and Cyanobacteria (6.1%). Principal coordinate analysis showed that the microbial community structure diff ers significantly at diff erent sampling points along the successional gradient, indicating that the bacterial community structure is sensitive to the lake wetland succession. Diff erent hydrological regimes and soil characteristics such as NH 4 +-N, Fe 2+, Mn 2+, and sediment organic carbon (SOC) content may be important factors responsible for the diff erences in the sediment microbial characteristics of the diff erent successional stages in the Caohai wetland. Additionally, it was found that the SR increased significantly from the P. lucens zone to the J. eff usus zone, but BR had the opposite response. The shifts in the bacterial community structure along the successional gradient may be the main reason for the observed diff erences in sediment respiration.
Keyword: sediment bacteria; microbial respiration; lake wetland; Caohai Wetland
1 INTRODUCTION
Wetlands, the transitional zone between aquatic and terrestrial environments, have important ecological functions, and play an important role in carbon cycling. In wetlands, the long-term flooding and hypoxia lead to an imbalance between net primary production and decomposition. Therefore, wetlands represent a major sink for atmospheric carbon (C)(Schulze and Freibauer, 2005). Although wetlands occupy less than 4%–6% of the terrestrial land surface worldwide, they contain approximately one-third of the world’s carbon (15× 1014kg) stored in terrestrial soil reservoirs (Kraigher et al., 2006; Kayranli et al.,2010).
Climate change is expected to increase the frequency and intensity of droughts in the future(Cook et al., 2014; Chen et al., 2018). Droughts in wetlands caused by climate change and human activity have been widely observed in recent years(Liu et al., 2006; Wu et al., 2017). It is estimated that more than half of the world’s wetlands will disappear in the next century. Both drought and drainage can result in significant reductions in water levels, leading to wetland succession. Wetland succession occurs because oflake filling. The succession includes various stages, specifically, the submerged plant stage, the erect plant stage, and the wet herbaceous stage. These diff erent succession stages have diff erent hydrological conditions, soil characteristics, and vegetation communities (Xia et al., 2015).
Wetland succession often threatens carbon stores,as increasing decomposition leads these wetlands to serve as a carbon source rather than a carbon sink.Drought-induced carbon loss in wetlands at high latitudes has been observed (Fenner and Freeman,2011; Planas-Clarke et al., 2020). Falling water levels result in increased oxygen availability in the soil,which potentially causes more rapid decomposition of organic matter. It is well known that soil respiration in wetlands under anoxic conditions is much lower than that in oxic environments (Chen et al., 2018).However, some studies have found that drought in wetlands may lead to unchanged or reduced CO2emission in some wetlands, thus increasing the carbon levels in these environments (Laiho, 2006; Noormets et al., 2010). Thus, the long-term outcome of sediment carbon remains unclear. Hence, it remains a significant pressing global problem to understand wetland succession eff ects and their inherent feedback mechanisms related to carbon decomposition (Wang et al., 2015). Because of changes in the climate or surrounding land use, wetland succession can result in altered local environmental conditions that can in turn directly aff ect the carbon cycle (Laiho, 2006).
Bacteria are important regulators of carbon cycling in lakes and are central to the processes of sediment ecosystem processes. Several studies have shed light on the link between wetland soil respiration and microbial community composition (Jaatinen et al.,2008; Mäkiranta et al., 2009), but the eff ects of wetland success to the sediment bacterial communities and respiration are still unknown. Caohai is a complete and a typical karst plateau lake wetland system,located in the Guizhou Plateau within the Changjiang(Yangtze) River watershed area. As the water level declined, the Caohai Lake showed succession from a shallow lake (submerged plant stage) to swamp (erect plant stage) to swampy meadow (wet herbaceous stage). In this study, we investigated the bacterial communities in sediment samples along the succession gradient using the high-throughput sequencing of 16S rRNA genes. Sediment microbial respiration(SR) was measured using the static chambers in the field, and sediment basal respiration (BR) was determined in the laboratory. While both SR and BR are measures of soil microbial respiration. SR reflects the in-situ soil respiration, while BR reflects the optimal microbial respiration. In other words, BR can be recognized as a measure of the aerobic decomposition potential of a substrate, that is, how much substrate can be utilized under the optimal oxygen and temperature conditions (Jaatinen et al.,2008). We aimed to: 1) characterize the changes in the bacterial community structure along the succession gradient; 2) understand how sediment respiration responds to the wetland succession and the relation with the microbial community structures; and 3)identify the main factors influencing microbial communities and sediment respiration.
2 MATERIAL AND METHOD
2.1 Site description
The Caohai Wetland (26°47′32″N–26°52′52″N,104°10′16″E–104°20′40″E) is located on the Guizhou Plateau, in southwestern China, at an altitude of 2 170 m above sea level and covers an area of 120 km2,with water covering 25 km2of the area (Xia et al.,2020). Caohai is subject to a subtropical climate, with an average annual temperature of 10.5 °C and a relative humidity of 79%. The average annual precipitation is 950.9 mm, with the majority (70%) of rainfall occurring from May to September (Sun et al.,2020). The unique geographical and climatic conditions contribute to a rich fauna and flora and the Caohai Wetland was designated as the National Wetland Nature Reserve in 1992.
In the 1970s, due to artificial drainage for grain farming, the water area of the Caohai Lake has decreased sharply from 45 km2to 5 km2but recovered to more than 20 km2after the 1980s. Because of the impact of regional climate change and human activities, the eco-hydrological situation of the Caohai Lake changed significantly over the years, with serious degradation of the wetland resulting in a clear hydrological gradient where the vegetation shifts from the submerged to the mesophyte succession.Four sample points (CH1, CH2, CH3, and CH4) were chosen based on the vegetation distribution. CH1 was located in the open water area of the lake, wherePotamogetonlucensis the dominant vegetation species at the deepest water level. In CH2,Scirpustabernaemontaniis the dominant vegetation species at the sub-deepest water level. In CH3, the vegetation is dominated byScirpustriqueterand shows seasonal submersion. In CH4, the driest area that is only occasionally submerged,J.eff ususdominates (Xia et al., 2015).

Fig.1 The sketch map of study areas at the Caohai Lake
2.2 Sediment sampling and experimental design
Sediment samples were collected in May 2014. In theP.lucens(CH1) andS.tabernaemontani(CH2)zones, undisturbed sediment core samples were collected using a plexiglas corer; while in theS.triqueter(CH3) andJ.eff usus(CH4) zones,samples comprising the top 10 cm of sediment were collected using a soil auger; Triplicate sampling plots were set in each site (Fig.1), and five sediment cores with a depth of 0 to 10 cm (excluding surface litter)were taken randomly from each plot, and mixed into one composite sample in a sterile plastic bag,transported to the laboratory on ice and kept at -80 °C.In addition, the surface material in five plots (0.1 m2),equally spaced along each sites, was clipped, and collected for aboveground plant biomass measurement.
In the laboratory, samples from each plot were homogenized for each plot and sieved through 2-mm mesh to remove visible stones, grass, and roots. A portion of the fresh sediment was used immediately for determination of sediment respiration and total water moisture content, and another portion was airdried and stored at room temperature until measurement of the other sediment characteristics.The remainder of the fresh sediment was stored at-80 °C for subsequent DNA extraction.
2.3 Sediment physicochemical properties analysis
Sediment physicochemical properties were measured using standard methods. Briefly, the sediment pH was measured by a pH meter (Sartorius PB-10, Germany), using a sediment to water ratio of 1:2.5. The Oxidation-Reduction Potential (ORP) was determined by an ORP analyzer (HAD-FJA-6, China).Ammonium (NH4+-N) and sediment nitrate (NO3ˉ-N)were extracted with 1-mol/L KCl and their concentrations measured using an Automated Discrete Analyzer (AQ2+, SEAL Analytical Inc., United Kingdom) (Guo et al., 2015). The sediment water content (SWC) was gravimetrically determined after drying at 105 °C for 12 h. The sediment organic carbon (SOC) was measured with the dichromate digestion method, and the dissolved organic carbon(DOC) using a TOC analyzer (Vario TOC, Elementar,Germany). The sediment bulk density (BD) was determined by the measurements of the cutting ring.The concentrations of Fe2+and Mn2+were analyzed using Atomic Absorption Spectroscopy (Zeenit700p,Analytik Jena, Germany). The water level (WL) was measured with a ruler. The plant biomass (PB) was the dry weight of the aboveground part of the plant.
2.4 Sediment respiration measurements
The SR in the CH1 and CH2 zones was measured using static floating opaque chambers (50 cm×50 cm×50 cm). The Field SR in the CH3 and CH4 zones was measured using the same chambers as used in the CH1 zone; in addition, aluminum collars were installed in the study sites prior to study initiation. At four time points, at 30-min intervals, a 40-mL gas sample was collected from each of the chambers using a polypropylene syringe.
The sediment BR was measured in the laboratory.Twenty grams of original fresh sediment samples were incubated in 250-mL serum bottles at 25 °C for 24 h. The concentration of CO2was measured via Gas Chromatography (GC-14C, Shimadzu) using a 10-m×2-mm stainless steel column and an Flame Ionization Detector (FID). The inlet, column, and detector temperatures were 50 °C, 50 °C, and 90 °C,respectively. H2served as a carrier gas with 40 mL/min flow rate (Tang et al., 2011a).
2.5 DNA extraction, library preparation, and sequencing
A Fast DNA spin kit for soil (MP Biomedical, CA,USA) was used to isolate gDNA from the sediment samples using the manufacturer’s protocols. The 515F and 907R primers (Xiong et al., 2012) were then used for amplifying the bacterial 16S rRNA V4-V5 region, and each sample was given an 8-nucleotide barcode during amplification. The thermocycler settings were: 95 °C for 2 min, 25 cycles of 95 °C for 30 s, 55 °C for 30 s and 72 °C for 5 min. Three PCR reactions were conducted per sample, with the total volume of 20-μL volume containing 10-ng template DNA, 2-μL dNTPs (2.5 mmol/L), 4 μL of 5×FastPfu Buff er (Transgen, Biotech, China), 0.4 μL of FastPfu Polymerase (Transgen), and 0.8-μL each of forward and reverse primers (5 μmol/L).
After amplification, 2% agarose gel electrophoresis was performed and an AxyPrep DNA Gel Extraction Kit (Axygen Biosciences, CA, USA) was used to isolate the resultant amplicons. Next, QuantiFluor™-ST (Promega, WI, USA) was used to quantify sample nucleic acid content. Equimolar amounts of all samples were then pooled and subjected to standard paired-end sequencing (2×250 bp) with an Illumina MiSeq platform (Illumina, San Diego, CA, USA).
2.6 Bioinformatic analysis
QIIME (v1.17) was used for the de-multiplexing and quality filtering of raw FASTQ files based on the following criteria: (i) truncation of reads was performed at any site with an average quality score of<20 using a sliding 50-bp window, with resultant reads <50-bp long being discarded; (ii) any reads that contained only barcodes, ambiguous reads, or two or more nucleotide mismatches were discarded; and (iii)only sequences with >10 bp of overlap were assembled based upon overlapping sequences. In addition, any reads which were not successfully assembled were discarded.
A 97% similarity score served as a cut-off criterion for operational taxonomic unit (OTU) clustering,which was performed using UPARSE v7.1, with UCHIME used to remove chimeric sequences (Edgar et al., 2011; Rui et al., 2015). Ribosomal Database Project (RDP) Classifiers were used to assign 16S rRNA sequence taxonomy against the SILVA(SSU115) database with a 70% confidence threshold(Amato et al., 2013).
2.7 Statistical analysis
To correct for sampling eff ects, we normalized all samples to an even number of sequences per sample(5604) for further analysis (including phylogenetic distance, Chao1 richness and Shannon’s diversity index, and observed species). Principal coordinates analysis (PCoA) was used tor assessing structural changes in bacterial communities based upon Bray-Curtis distance, using R (http://cran.r-project.org/web/packages/vegan/index.html) for all analyses.Variance partitioning analysis (VPA) and canonical correspondence analysis (CCA) were conducted with CANOCO5 (Microcomputer Power, Ithaca, NY) in order to assess how diff erent environmental variables(WL, SWC, SOC, NH4+-N, NO3ˉ-N, Fe2+, Mn2+, ORP,and PB) contributed to observed phenotypes. Oneway analyses of variance (ANOVAs) were used for comparing relative OTU abundance between samples,with Pearson’s correlations were used for comparisons of diversity, species abundance, soil properties, and environmental factors with SPSS 18.0 used for analysis.
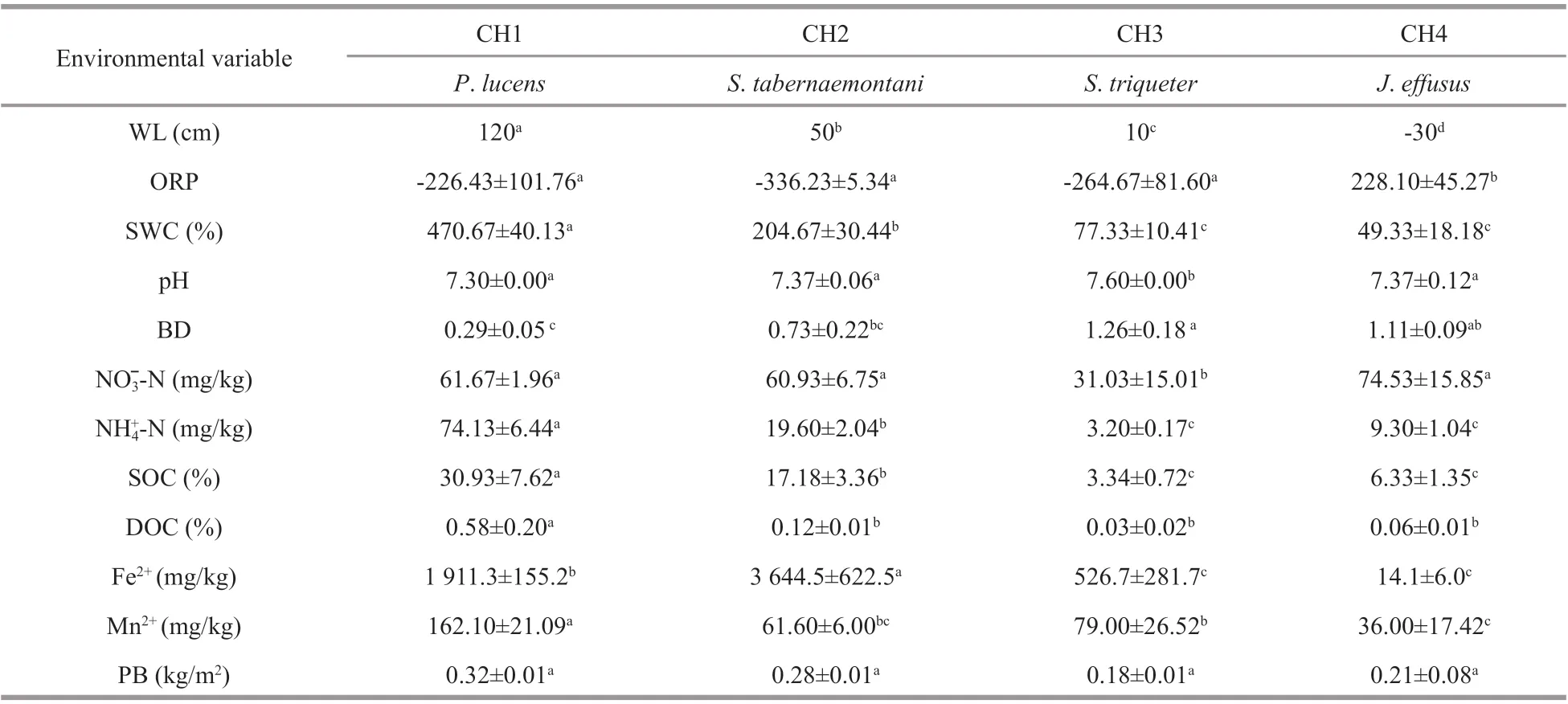
Table 1 The sampling stations and sediment physicochemical parameters with standard deviations for each plot
3 RESULT
3.1 Changes in sediment properties among the diff erent successional gradient
The basic physicochemical characteristics for the diff erent successional gradients are shown in Table 1.The hydrological regime (SWC and WL) showed a significant declining trend along the wetland succession (from CH1 to CH4), while the ORP was significantly increased (P<0.05), and the sediment BD increases firstly and then decreases. Most of the physical and chemical properties (SWC, NH4+-N,SOC, DOC, and Fe2+) of the sediment showed decreased in value from the CH1 zone to the CH4 zone. In addition, the Sediment Organic Carbon(SOC), which was highest (30.93%±7.62%) in CH1 decreased to its lowest level (3.34%±0.72%) in CH3.However, the PB and pH values were similar among the diff erent points in the successional gradient, with the exception of the CH3 zone.
3.2 Changes in the overall bacterial communities along the successional gradient
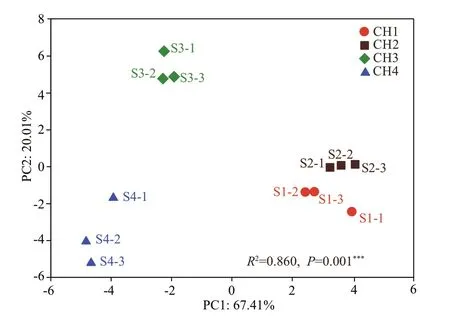
Fig.2 Principal coordinate analysis of soil bacterial communities using weighted UniFrac
In total, 98 292 reads were obtained on Illumina sequencing. The average read length was 397 bp, and the samples contained 5 604 to 10 165 reads. A total of 845 OTUs (each OTU containing more than five reads) were identified in these samples based on a 16S rRNA sequence similarity threshold of 97%. The number of OTUs, Chao1 richness index and Shannon index suggested that both richness and diversity were the highest for the CH2 zone samples. Simpson index was the highest in CH3 zone and lowest in the CH2 zone (Table 2). Overall, there was no significant change in the diversity index between the CH1 zone and the CH4 zone, although higher values were seen in the CH2 zone (Table 2).

Fig.3 Changes in the dominant bacterial groups (i.e., the percentage compared to the total number of reads) at the phylum level along the degradation gradient in Caohai Lake
The similarity of the bacterial communities in the 12 samples were evaluated via PCoA and the PERMANOVA test. The bacterial communities in these 12 samples clustered well into four groups(Fig.2): (i) group I contained the three samples from the CH1 zone with the highest water level; (ii) group II contained the three samples from the CH2 zone, in which water level was lower than that in the CH1 zone; (iii) group III contained the three samples from the CH3 zone, which is often subject to dry to wet cycling; (iv) group IV contained the three samples from the CH4 which is dry and inundated occasionally.The PERMANOVA test showed that the bacterial community structures diff er significantly in the four diff erent successional stages (R2=0.860,P=0.001).These results indicate that the bacterial community structure is sensitive to wetland succession.
3.3 Changes in the dominant bacterial groups among the diff erent stages of the successional gradient
The relative abundances of the dominant bacterial phyla changed significantly along the successional gradient (Fig.3). The core bacterial phyla were defined as those detected at least two sites and with relative abundances higher than 5% in at least one site. The average abundance of core bacterial phyla was, in descending order, Proteobacteria (34.7%), Chloroflexi(17.8%), Bacteroidetes (7.3%), Acidobacteria (6.6%),and Cyanobacteria (6.1%). The relative abundances of Acidobacteria, Chloroflexi and Actinobacteria increased, whereas the Cyanobacteria decreasedalong the successional gradient. However,Proteobacteria and “Unclassified taxa” first increased and then decreased along the successional gradient.Bacteroidetes decreased from the CH1 zone to the CH3 zone, but increased slightly in the CH4 zone.These results indicate that the relative abundance of the core bacterial phyla diff ered along the diff erent successional gradient.
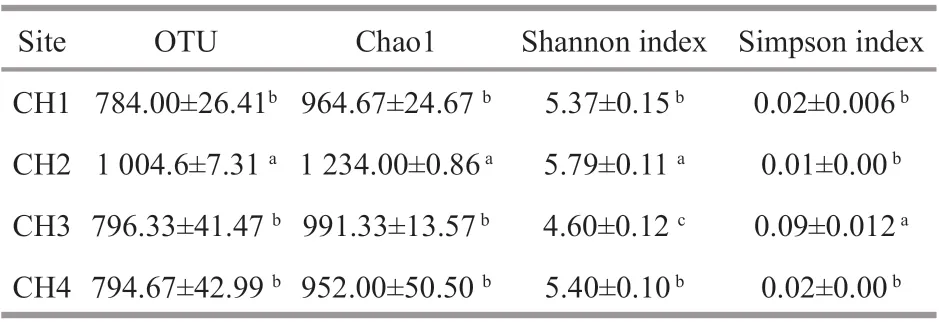
Table 2 Bacteria diversity indices calculated based on 97%similarity of 16S rRNA gene sequences
3.4 The relationship between environmental parameters and bacterial communities

Fig.4 Canonical correspondence analysis (a) and corresponding variation partitioning analyses tested the correlation between bacterial community structure and environmental factors (b)


3.5 Sediment respiration along the successional gradient
SR and BR showed the diff erent patterns along the successional gradient (Fig.5). The SR varied greatly among locations (P<0.05): it was the highest (0.85 g CO2/(m2∙h)) in the CH4 zone and the lowest(-0.02 g CO2/(m2∙h)) in the CH1 zone. BR rates showed the opposite pattern. BR significantly decreased along the successional gradient. The highest rates were observed in the CH1 zone and the lowest rates in the CH4 zone. The CH1 zone represents anaerobic conditions, where the progress of decomposition is aff ected in the sediment. In the CH4 zone, the sediment is exposed to aerobic decomposition, thus the decomposition rate is likely to be higher in this site.


Table 3 Pearson’s correlation analysis of hydrological parameters with sediment respiration and basal respiration along the successional gradient
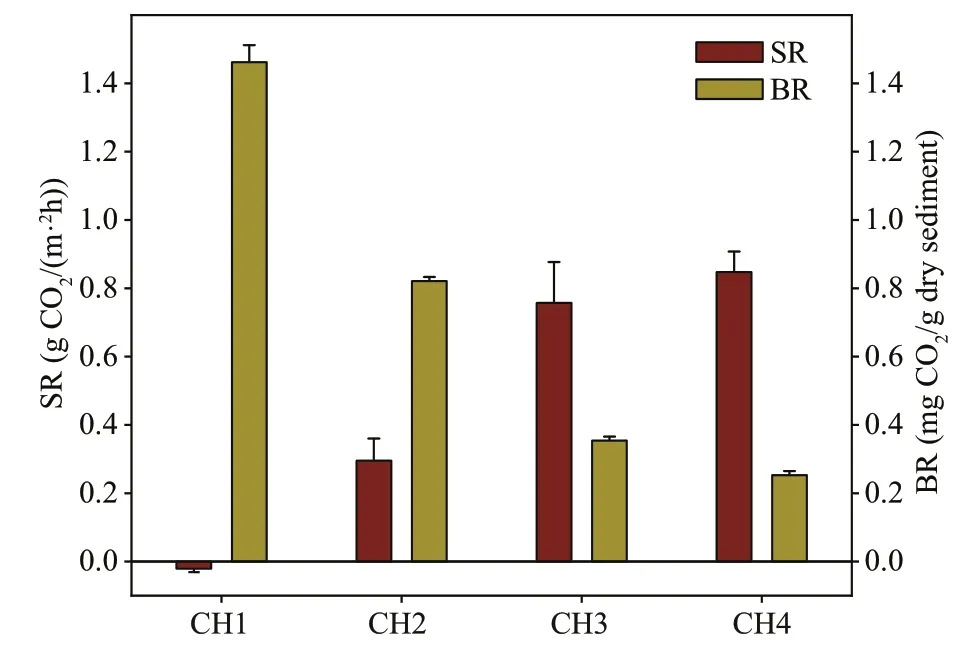
Fig.5 The changes of the sediment respiration (SR) and basal respiration (BR) rates along the succession gradient in Caohai Lake
4 DISCUSSION
Although there have been many reports on the study of microbial communities in wetland sediments,the eff ects of succession on these communities and the wetland carbon stocks are relatively unknown. In this study, the results showed that there are significant diff erences in the hydrological regime, plant community species composition and sediment physics and chemistry at diff erent wetland successional stages. The microbial communities are also vary from the CH1 zone to the CH4 zone. All these factors correlate significantly with the bacterial community composition. These results suggest that changing environmental factors aff ect bacterial community composition in the sediments along the wetland succession and that these compositional diff erences may account for the observed change of respiration,leading to a loss of sediment carbon storage.
4.1 Eff ect of hydrological regime and soil characteristics on the bacterial community structure
Bacterial communities vary significantly from the CH1 zone to the CH4 zone (Fig.2). These results are consistent with those of a previous study showing variations in the bacterial community composition along the successional stages of the Chongming Dongtan Wetland (Zhang et al., 2013). Tang et al.(2011b) have reported that microbial community structural shifts are linked to changes in the sediment microenvironment. It has also been found that soil microbial communities can be influenced by a wide range of variables, including sediment physicochemical characteristics and climate warming (Balasooriya et al., 2008; Bond-Lamberty and Thomson, 2010; Burns et al., 2015; Jeanbille et al., 2016; Bulseco et al., 2020).The sediment ORP has been considered to be a fundamental indicator of such changes because many sediment biochemical processes are closely associated with the oxygen concentration in sediment, which is,in turn, related to the water level and vegetation. In this study, it was found that diff erent water levels led to diff erent microenvironments and thus to altered microbial community composition along the successional gradient.
We observed significant relationships between the hydrological regime, sediment physicochemical properties, and microbial community composition.Most of the bacterial phyla that were significantly associated with the water level were also significantly correlated with the soil physicochemical properties.The hydrological regime not only directly aff ects the microbial community (Ma et al., 2018) but also indirectly aff ects the microbial community structure by altering the soil physicochemical properties.Therefore, sediment characteristics and hydrology are important factors driving the shifts in soil microbial characteristics of the diff erent parts of the successional gradient in the Caohai Wetland. Sediment properties and hydrology have been observed to significantly aff ect the microbial community composition in floodplains (Mayr et al., 2020). Vegetation also has an important impact on the spatial variation of sediment microbial communities (Jiang et al., 2013). Plant type and sediment physicochemical properties are the two primary factors driving the composition of the soil microbiome due to complex interactions between plants, sediments, and microbes (Garbeva et al., 2004;Mitchell et al., 2010; Yin et al., 2020). However, in this study, no significant relationship between the bacterial relative abundance and plant biomass was found, indicating that the impacts of plant biomass are marginal. This does not, however, mean that the vegetation is not an important factor aff ecting the bacterial communities in sediments in the Caohai Wetland, but indicates that the influence of plants on the bacterial community is not related to the plant biomass. The role of plants in the soil microbial community construction should be further investigated in the Caohai Wetland. Above all, diff erent hydrological regimes and sediment characteristics such as NH4+-N, Fe2+, Mn2+, and SOC may be important factors resulting in the diff erences in sediment microbial characteristics of diff erent successional stages in the Caohai Wetland.
4.2 Eff ects of wetland succession on SR and BR and their relationships with bacterial community
SR and BR showed diff erent patterns in relation to the diff erent stages of the successional gradient(Fig.5). SR clearly increased from the CH1 zone to the CH4 zone, whereas BR showed the opposite trend. SR was the sediment respiration measured in situ. Along the succession, the water depth gradually decreased and the ORP gradually increased from the CH1 zone to the CH4 zone, indicating that the sediment environment had changed from anaerobic to aerobic. Therefore, the decomposition of organic matter also changed from anaerobic to aerobic decomposition, resulting in a gradual increase in SR.BR can be recognized as a measure of the aerobic decomposition potential of a substrate, that is, how much substrate can be utilized under the optimum oxygen and temperature conditions (Jaatinen et al.,2008). BR was higher in theP.lucens-dominated zone CH1, because these areas represent anaerobic conditions, where the progress of decomposition is impeded in situ. However, in theJ.eff usus-dominated zone CH4, the surface soil has been exposed to aerobic decomposition, and thus the decomposition potential is likely to be higher. BR had a significant positive correlation with SOC (Table 3), suggesting that SOC might be an important factor that influences BR. Sediment substrate (carbon) availability might be a key factor that influences the BR (Jaatinen et al.,2008; Schiller et al., 2019). The soil carbon content decreased from the CH1 zone to the CH4 zone,resulting in a lower rate of BR.
Variation of SR at diff erent stages along the successional gradient may be related to the shifts in the relative abundance of specific microbial populations. The shifts of microbial community structure have been shown to be linked to SR changes in wetland sediments, and the diff erences in sediment microbial community structure in diff erent wetland sediments are important reasons for the diff erent SR values (Li et al., 2011). Diff erent microbial community composition may be a major reason for diff erences in SR along the successional gradient (Jaatinen et al., 2008). Proteobacteria are a group of aerobic or facultative bacteria that often have highly versatile successional capacities (Yang et al., 2005; Sato et al., 2009), which might contribute to the decomposition of organic matter owing to its high abundance in Caohai Wetland. The phylum Actinobacteria is another major taxon that is capable of organic matter decomposition (Tang et al., 2011b),and the relative abundance of Actinobacteria increased dramatically along the successional gradient, which suggests it might be a key taxon driving the changes of SR in the Caohai Wetland.The relative abundance of Anaerolineae and Acidobacteria also increased along the successional gradient, which might also contribute to the change of SR. Substrate is a strong determinant of the microbial community (Chen et al., 2008; Eilers et al., 2010), as the changes that take place in both the carbon quantity and quality of the wetland succession are likely to aff ect the soil communities. Therefore,the composition and community-level physiological profiles of the soil microbial communities change along the wetland succession, which, in turn, aff ect sediment microbial respiration (Tang et al., 2011a).
Overall, diff erences in the hydrology and vegetation of diff erent stages of the successional gradient could lead to diff erences in the soil physical and chemical characteristics, including SWC, SOC, NH4+-N and ORP, which might consequently lead to diff erent soil microbial community compositions. This explains why the bacteria with high successional ability, such as Acidobacteria, Actinobacteria and Chloroflexi,were found to be more abundant in CH4 zone with the lowest water level, which may be an important factor contributing to the higher SR in this site.
5 CONCLUSION
These results support the following conclusions:(1) the wetland succession is accompanied by changes in bacterial community structure; (2) the presence of distinct microbial communities in wetland sediment samples may be the main reason for the observed diff erences in rates of decomposition, which, in turn,aff ect the sediment carbon cycle; (3) hydrological conditions and sediment properties were the major factors driving the shifts in the bacterial communities.Overall, this study indicates that the wetland succession has significant eff ects on the composition of soil microbial communities and that these eff ects can alter the carbon cycle of the lake wetland.
6 DATA AVAILABILITY STATEMENT
The data that support the findings of this study are available from the corresponding author.
杂志排行

Journal of Oceanology and Limnology的其它文章
- Steady increase in water clarity in Jiaozhou Bay in the Yellow Sea from 2000 to 2018: Observations from MODIS*
- Phylogenetic diversity and bioactivity of culturable deepsea-derived fungi from Okinawa Trough*
- Allelopathic eff ects of mixotrophic dinoflagellate Akashiwo sanguinea on co-occurring phytoplankton: the significance of nutritional ecology*
- Investigation of the decline of Ulva prolifera in the Subei Shoal and Qingdao based on physiological changes*
- Effi ciency of phosphorus accumulation by plankton,periphyton developed on submerged artificial substrata and metaphyton: in-situ observation in two shallow ponds*
- Petroleum exploitation enriches the sulfonamide resistance gene sul2 in off shore sediments