菌-藻共生生物膜污水处理研究进展
2021-06-15廖怀玉孙丽李济斌何頔
廖怀玉,孙丽,李济斌,何頔
(广东工业大学 环境生态工程研究院 广州 510006;南海海洋科学与工程广东省实验室(广州),广州 511458)
随着工业化的不断推进,人类生产生活产生的废水日益增多。水体中富含氮磷等营养元素,其中,工业废水中更含有难降解有机物、致病菌等有毒有害物质,此类废水若未经有效治理而直接排放至天然水体,严重威胁生态环境及人类健康[1]。目前,污水处理技术以活性污泥法及其衍生工艺为主,这些传统工艺虽然技术成熟、工艺稳定,但存在运行能耗高、泥水分离效果差、难降解有机物处理处置效果不佳、微生物对环境毒性以及盐度等耐受力差等一系列问题。菌-藻共生污水处理技术的兴起有助于缓解污水处理压力,利用细菌与微藻协同作用机制、藻体强耐受能力及生物质高效资源化,可同步实现水中污染物的有效去除与微藻生物量收获,具有运行成本低、能耗小、效率高等优点[2],能有效克服传统污水处理工艺存在的弊端。因此,菌-藻共生污水处理技术引起了学者们的广泛关注。
自20世纪50年代Oswald等第一次利用菌-藻协同净化污水以来,以菌-藻共生为基础的污水处理技术在水质净化机理、藻种筛选、反应器设计、工艺条件控制及藻细胞加工利用等方面取得了积极的进展。目前,菌-藻共生体系主要以悬浮、固定化以及菌-藻共生生物膜的形式存在。在悬浮态菌-藻系统中,因微藻自身尺寸小且密度接近于水,在水中多以悬浮态存在,易随出水流出,造成藻细胞流失,影响污水处理效果[3];固定化能够有效克服悬浮态菌-藻存在的弊端,但因存在包埋基质价格昂贵、无毒无害基质难寻、操作繁琐复杂等问题[4],没有得到广泛应用。为突破上述两种菌-藻共生体的技术瓶颈,菌-藻共生生物膜系统应运而生,并得到广泛发展。将惰性载体引入污水处理体系,利用菌-藻定向吸附特性,在载体表面形成结构稳定的菌-藻共生生物膜,最终强化污水处理效果[5]。笔者以分析菌-藻共生生物膜污水处理技术的关键原理为出发点,综述目前菌-藻共生生物膜污水处理的研究进展,主要内容包括:菌-藻共生生物膜的形成过程和污水净化原理;菌-藻共生生物膜形成的主要影响因素;菌-藻共生生物膜应用于污水处理的优势;菌-藻共生生物膜污水处理的应用。并对菌-藻共生生物膜污水处理应用过程中存在的问题进行分析总结,从而对菌-藻共生生物膜污水处理未来的发展方向进行展望。
1 菌-藻生物膜净化污水优势
相较于传统活性污泥法和悬浮菌-藻共生法污水处理技术,菌-藻生物膜体系具有独特优势,主要包括:系统抗冲击负荷能力强,污染物去除效果好,节能降耗。
1.1 系统抗冲击负荷能力强
细菌分泌的胞外聚合物(Extracellular polymeric substance,EPS)能促进微藻与细菌在载体表面附着,并为菌-藻共生体提供一层保护屏障,抵抗极端环境(如干燥、极端的pH值和温度)及缓冲有毒物质侵害[6-7],同时,增强生物膜结构稳定性。此外,菌-藻生物膜系统微生物群落结构丰富[8-9],可同时存在自养和异养微生物,有利于维持菌-藻生物膜的生态系统稳定。自养微生物主要包括光能自养菌(如蓝细菌、绿弯菌等)、化能自养菌(如硝化细菌等)、藻类(硅藻、绿藻等);异养微生物主要包括异养细菌、真菌及原生动物[10-11]。自养和异养微生物间能基于代谢产物交换形成互利共生关系,更好地适应寡营养环境。Wen等将微藻分泌的胞外有机物(Extracellular organic matter,EOM)作为WGX-9的碳源,结果发现,在C/N为5的环境下,仍具有较好的脱氮效果。
1.2 污染物去除效果好
菌-藻间通过代谢产物交换维持良好的互利共生关系,使菌-藻生物膜保持较高的生长代谢活性,同时,微藻对氮磷无机盐具有高效的同化作用,有利于实现同步脱氮除磷。Tang等[12]在研究菌-藻生物膜SBR系统与对照组不含藻类的SBR系统时发现,实验组菌-藻生物活性较对照组有所提升;此外,对照组TN、TP的去除效果分别为47.11%±4.79%、31.86%±2.25%,而试验组TN、TP的去除效果分别为65.8%±6.86%、89.3%±4.49%,证实菌-藻生物膜体系具有更优异的营养盐去除效果。
1.3 节能降耗
一方面,微藻光合作用产氧可降低系统曝气强度,节约能耗。Babu等[13]研究发现,菌-藻生物膜在光照强度为85~95 μE/(m2·s)时,其溶解氧(Dissolved oxygen,DO)浓度可达3.2~4.1 mg/L,硝化速率1.1~1.6 g N/(m2·d);Yang等[14]研究发现,在没有额外曝气且持续光照的条件下,菌-藻生物膜反应器对模拟生活废水中的COD和氨氮去除效果均可达90%以上。另一方面,附着生长体系回收生物质时需输入的能量更低。由于微藻细胞自身的特性,藻水分离难,不仅影响出水水质,还会导致系统难以维持高浓度生物量[3]。在菌-藻生物膜系统中,微生物通过自身的粘附性相互聚集并附着生长在载体表面,且从载体表面刮取的生物质含水率较低[15],降低了后续生物质回收时的能量输入。
2 菌-藻共生生物膜形成过程及净水原理
2.1 菌-藻共生生物膜形成过程
菌-藻共生生物膜的形成过程可分为3个步骤,即迁移运输、附着及生长过程[3-4,16](具体如图1所示)。1)迁移运输过程:菌-藻生物膜形成初期,悬浮细菌和微藻在定向吸附、重力作用、水力动力以及藻类趋光性等共同作用下,向固体介质表面运输、移动。2)附着过程:迁移至载体表面的微藻、细菌细胞与载体发生相互作用,依次进行可逆附着和不可逆附着。其中,可逆附着主要包括微生物利用鞭毛、纤毛等外部细胞器以及外层膜蛋白在载体表面发生物理性附着,由于该过程的粘附性较弱,易受外力的影响,发生脱落;不可逆附着是指初步附着的藻体和细菌通过分泌EPS,使其紧密粘附在载体表面,形成初级生物膜,不易受外界环境影响而脱落。3)生长过程:根据微生物的群落结构特征将生物膜生长过程分为细菌增殖阶段、微藻增殖阶段以及成熟阶段。细菌生长繁殖速度高于微藻,故生物膜生长初期优势种群以细菌为主,在细菌增殖阶段,细菌快速生长的同时分泌大量EPS;在微藻增殖阶段,绿藻、丝状藻以及其他藻类附着在EPS表面,同时,利用周围环境的营养物质增殖;在成熟阶段,生物膜中藻类和细菌的比例达到相对稳定状态,并形成良好的互利共生关系。最终,附着在载体表面的细菌和藻类形成一个成熟稳定、由EPS胶连,且具有三维结构的网状菌-藻共生体。

图1 菌-藻共生生物膜形成过程示意图Fig.1 Schematic diagram of the bacterial-algal symbiotic
2.2 菌-藻群落结构及相互作用关系
成熟的菌-藻生物膜具有丰富的微生物群落结构,其中,细菌多以α变形菌、β变形菌、γ变形菌以及鞘脂杆菌为主[17-18],藻类多以丝状绿藻、绿球藻为主[17]。其来源主要包括自然生长、原始接种液投加及后期物种进化演替。与自然生长相比,针对不同废水及工程所需,选择特定的微藻及细菌投加到系统中能够强化污染物去除效果。如无色杆菌(Achromobacteria)、产碱菌(Alcaligenes)、芽孢杆菌(Bacillus)、噬氢菌(Hydrogenophaga)和假单胞菌(Pseudomonas)可以进行异养硝化和好氧反硝化,脱氮效率高[19]。一些聚磷菌如假单胞杆菌(Pseudomonas)和Accumulibacter能超量吸磷并将其转化为胞内聚合磷化合物[20]。人工接种可强化所选物种的丰度,提高微生物生长速率,从而实现快速高效的污染物去除效果。当原始微生物形成初级生物膜后,生物膜中物种发生演替,这种演替主要受环境因素影响,进而影响菌-藻间相互作用关系,主要包括协作共生及竞争关系。微藻与细菌的生物协同作用在微生物代谢和生长、污染物去除能力和生态功能中起着至关重要的作用。细菌及藻类间不仅发生CO2和O2的交换,同时,彼此释放促生长因子及微量元素,促进对方生长发育,增强菌-藻共生体生物活性,Roeselers等[21]研究发现,Betaproteobacteria释放的一种微量物质能够促进Microcoleusvaginatus的生长及粘附作用;同样,Park等[22]研究发现,将Brevundimonassp.添加至Chlorellaellipsoidea中能够延长其对数增长时间,生物量增加50倍。菌-藻间竞争关系多出现于营养物质匮乏以及特定菌-藻间的敌对作用,相关研究发现,有些微藻的代谢物具有杀菌作用,如小球藻素对革兰氏阳性菌和革兰氏阴性菌具有杀菌作用[23],而一些藻类产生的代谢物能够抑制细菌的群体感应,不易于生物膜形成[24]。同样,一些细菌也会分泌链霉素等有毒活性物质,影响藻类光合作用相关基因转录,阻碍微藻电子传递,进而抑制藻类生长繁殖[25]。总的来说,微藻-细菌之间的协作共生或竞争作用是调节菌群结构和对外界条件的响应机制,进而影响菌-藻共生生物膜的形成及水质处理效果。
细菌间、菌-藻间存在着复杂的相互作用关系,而物种的选择是建立一个菌-藻稳定共生、污染物高效去除的菌-藻共生生物膜体系至关重要的环节。然而,目前所研究的附着菌群大多是通过自然培养形成,这可能会在系统效能方面达不到预期效果。随着遗传学和代谢组学的迅猛发展,菌种的基因改良为实现特定难降解有机污染物的高效去除提供了可能,通过针对性的基因改良,新物种的添加强化了某种生物功能,改善了菌种间相互作用关系,但新物种的添加在提高污染物去除效果的同时,对生态安全具有潜在风险[26]。因此,从生态工程角度出发,应优先选择和改进生物膜中土著微生物物种,以形成稳定的群落结构。所以,合理利用现代基因工程和生态工程手段对菌-藻生物膜进行生物强化具有一定的实际意义。
2.3 菌-藻共生生物膜净化污水原理
2.3.1 碳的去除 菌-藻生物膜中含碳污染物的去除主要通过异养细菌、兼养藻类的氧化分解作用以及自养藻类的光合作用。异养细菌以有机碳源污染物为电子供体,O2为电子受体,通过矿化作用将含碳有机物氧化分解为CO2,其中一部分CO2逸散至环境中,另一部分被藻细胞吸收利用,吸收进藻细胞的CO2在核酮糖二磷酸羧化酶(Rubisco)的作用下,经过暗反应的卡尔文循环转化为葡萄糖等有机物固定在微藻细胞内;同时,微藻光合作用产生的O2为异养细菌提供了充足的电子受体,强化了细菌对含碳有机物的氧化分解过程。当水环境偏碱性时,水体中的无机碳主要以HCO3-的形式存在,微藻可利用胞外碳酸酐酶将HCO3-通过主动运输吸收至细胞内,转化为CO2,再进行利用,从而实现污水中无机碳的去除[27]。此外,生物膜中兼性营养型的微藻可以CO2及有机碳为碳源,同时进行光合作用和呼吸作用,完成污水中含碳物质的去除[28]。
2.3.2 氮、磷的去除 菌-藻共生生物膜去除含氮磷元素污染物主要包括生物及化学两种途径:其中,生物途径主要为细菌及微藻的同化、异化作用,化学途径则是共生环境中pH值的变化引起相应的氮磷去除。当含氮有机化合物进入水体后,生物膜好氧区域中的细菌利用藻类光合作用释放的O2,经过氨化反应、硝化反应等将有机氮化合物分解转化为硝酸盐氮等无机氮化合物,生成的硝酸盐氮在生物膜的厌氧区域内经反硝化作用,还原成气态氮,完成细菌生物脱氮的过程;同时,经细菌氧化分解产生的无机氮化合物通过同化作用吸收转化为细菌(主要吸收氨氮)及微藻胞内物质,实现自身生长发育[8]。值得注意的是,微藻对无机氮磷的吸收具有一定顺序,当NH4—N、NO3—N和NO2—N在水体中共存时,微藻优先吸收NH4—N,并入碳骨架合成胞内物质,当NH4—N被消耗尽时,NO3—N和NO2—N作为N源被微藻吸收利用,并在硝酸盐还原酶以及亚硝酸盐还原酶的作用下转化为NH4—N,再被微藻同化为胞内物质[4, 28]。针对菌-藻生物膜生物除磷途径,其主要为聚磷菌及微藻的过量吸磷作用,Schmidt等[29]曾报道,小球藻(Chlorellavulgaris)和莱茵衣藻(Chlamydomonasreinhardtii)具有超级吸磷作用,其吸磷能力强于同质量下的聚磷菌。化学脱氮除磷主要是由于藻类光合作用消耗了水中大量酸性CO2,使得水体pH值升高,呈碱性状态,氮元素则在碱性条件下部分以NH3形态溢出水体得以去除[30],磷在碱性条件下易与共存的钙镁离子形成难溶性磷酸盐沉淀,实现去除;此外,有研究者在研究菌-藻生物膜EPS时发现,菌-藻生物膜中的EPS含有蛋白质和多聚糖等成分,具有丰富的含氧官能团,能够与有机磷发生配位作用,使有机磷与菌-藻生物膜表面的EPS结合,通过吸附作用去除[31]。菌-藻共生生物膜去除污水中的C、N、P污染物原理如图2所示。

图2 菌-藻共生生物膜去除C、N、P示意图Fig.2 Schematic of the pollutant removal mechanism
2.3.3 重金属的去除 菌-藻生物膜对重金属的去除原理主要包括吸附作用、生物富集以及由水体pH值变化引起的沉淀作用[5]。菌-藻生物膜对重金属的吸附作用主要包括静电吸附和络合作用。其分泌的EPS中含有含氧官能团,如羟基、羧基、氨基和膦酰基,具有两方面的作用,即:使EPS表面带负电,通过静电作用力吸附重金属离子;提供有效结合位点,使重金属与氧、氮、磷等原子通过配位作用生成配位键,形成络合物[32]。除EPS的吸附作用,菌-藻生物膜还可通过细胞生物膜和细胞壁的吸附作用去除重金属,这是由于微藻和细菌的细胞壁或细胞膜成分中包含蛋白质、多聚糖等成分,其表面的配位基团包含的阴离子基团和酸性官能团,如氨基、羧基、羟基以及硫化物等,具有与EPS类似的吸附作用,通过静电吸附和络合作用将重金属离子吸附至微生物细胞表面。发生在活细胞体内的生物富集作用也是重金属去除的途径之一,微生物通过主动运输和胞吞作用使重金属进入细胞质并与细胞器相结合,使重金属在细菌和微藻体内发生生物富集[5,33]。除此之外,由于微藻光合作用引起水体环境pH值升高,可使重金属在碱性环境下通过沉淀作用去除。
3 菌-藻共生生物膜形成的影响因素
菌-藻共生生物膜的生长状况、附着状态均能影响污水处理效果。菌-藻生物膜同细菌类生物膜类似,当其生长到一定阶段时,将发生脱落,引起菌-藻生物膜脱落的原因主要包括:水体中的底物被逐渐消耗,使得微生物生长进入稳定期和衰老期;由于环境扰动,水体剪切力大于菌-藻生物膜与固体介质之间的吸附力,使生物膜发生机械性脱落;随着生物膜厚度增加,内层微生物的传质阻力增加,以及光能利用降低,使得内层细菌活性和微藻的光合作用效率下降,导致生物膜粘附性减弱而发生脱落。因此,通过认识菌-藻生物膜形成的影响因素,设置合理的工艺参数,对提升生物膜稳定性、防止或降低脱落频次,对保证工艺高效稳定运行具有重要意义。影响菌-藻生物膜形成的因素主要包括环境因素、生物因素、载体类型等。
3.1 环境因素
光照、CO2浓度、温度、水质特征及水流速度等环境因素均会影响菌-藻生物膜污水处理效果。光照是藻类生长过程中关键性限制因素之一,主要通过光照强度及周期影响藻类生长。在一定的光照强度范围内,微藻的光合作用速率随着光照强度递增而增加,当达到光饱和点时,增加的光照强度会抑制藻细胞内色素吸收光和光能转化的效率,从而影响微藻自身的合成代谢,减缓菌-藻生物膜的形成[34-36]。光照强度对菌-藻共生生物膜的微生物群落结构具有一定的影响。由于生物膜具有一定厚度,光照强度从生物膜表层至内层呈指数衰减,导致微藻光合作用活性随生物膜厚度降低,从而引起氧浓度在生物膜上呈梯度分布[37],因此,生物膜上分布着不同氧含量需求的菌种。此外,光照强度对细菌活性也有一定影响。大量研究表明,强光辐射的环境条件下,氨氧化菌和亚硝酸盐氧化菌的活性都将受到一定程度的抑制。Meng等[38]研究表明,当光照强度高于8 000 lx时,菌-藻共生体系中亚硝酸盐氧化菌活性受抑制,造成水体中亚硝酸盐的累积;Hooper等[39]研究发现,强光主要通过诱导高氧化活性物质的形成,损坏细菌细胞结构,并通过抑制氨单加氧酶和细胞色素-C(一种与电子传递链有关的蛋白)的活性来降低氨氧化细菌和硝化细菌的活性。除光照强度外,光照周期对藻类细胞生长繁殖也具有重要作用,微藻光合作用主要包括光反应和暗反应两个阶段,其中,光反应阶段在光照、光合色素以及酶的作用下将光能转化为化学能储存在细胞内;暗反应阶段则利用光反应产生的化学能以及酶的作用将CO2还原为糖类,转化为胞内物质,所以,适宜的光暗交替有利于光、暗反应相结合,是菌-藻共生生物膜微藻细胞增殖及代谢正常运行的重要保障[40]。
CO2是藻类生长过程中的另一限制性因素。Blanken等[41]研究表明,当水体中通入的CO2含量由0.63%增加到1.25%时,微藻生物量显著提高,随后,当CO2含量由4%增加至10%时,微藻生长并没有明显变化,可见,微藻对CO2的吸收利用存在饱和点,浓度过高或过低均不利于生物膜中微藻的生长繁殖。此外,CO2供气方式对生物膜稳定性具有一定影响,研究表明,半浸没式载体(即部分裸露于空气中)可直接吸收空气中的CO2,气体传递至生物膜表面只需穿过一层较薄的液膜,因此,传质阻力小、传输效率高;而浸没式培养体系通常采取在系统底部曝气的方式提供CO2,该方式传质阻力较大、传质效率较低,同时,曝气形成的上升气流对生物膜表面具有剪切扰动作用,不利于微生物附着,降低了生物膜的稳定性,因此,需设置合理的CO2供气方式及强度[42]。

污水水质,如水体透明度、C/N、DO等均对菌-藻生物膜活性和群落结构会产生重要影响。在菌-藻共生体系中,水体透光度对藻类光合作用影响非常大,水体透光度与水中悬浮颗粒含量及色度有关,其中,悬浮颗粒对光具有很强的散射作用,而水体色度影响光的透射距离,进而影响藻类对光的吸收利用[46]。水体透光性还受污泥/藻含量比值影响,Sun等[47]研究了泥/藻比对藻类生长的影响,发现随着污泥含量增加,水体颜色由绿色转为黄褐色,遮光效应增强,抑制了微藻光合作用活性和增殖速率。除水体透明度外,C/N、DO也通过影响共生菌的生长繁殖进而影响菌-藻共生生物膜的形成,研究者们发现,高C/N比能够促进异养细菌大量繁殖,自养菌种群密度和活性逐渐下降[10, 48]。生物膜中DO梯度影响氨氧化、硝化和反硝化有关的细菌的分布情况[49],进而间接影响自养异养菌的空间分布,而藻类在DO含量较高的条件下光呼吸作用增强,导致内源物质消耗而光合作用受抑制[50],所以,有效控制C/N和DO浓度是维持菌-藻平衡的一个重要因素。
水体流速是菌-藻共生生物膜的形成、挂膜以及稳定性的重要影响因素。水体流速对菌-藻生物膜形成的影响主要表现为:一方面,流速增加促进底物传质,有利于生物膜中底层微生物吸收营养物质,提高生长繁殖速率;另一方面,老化的生物膜可以在水体剪切力的作用下发生脱落,实现菌-藻生物膜更新,从而增强生物膜中微生物活性[8]。然而,水体湍动程度过大将产生较大的剪切力,不利于生物膜在载体表面附着,抑制菌-藻生物膜的形成及稳定,过小则易形成较厚的生物膜,增加了敝光效应和传质阻力,不利于内层生物膜对光能的利用以及生物膜与液相主体之间的底物、代谢产物以及气体交换。因此,水体流速需同时满足生物膜附着生长的稳定性及底物传质的低阻力,并根据生物膜形成的不同时期设定。在生物膜形成初期,较低的流速可以促进微生物附着而有利于生物膜的形成,而在后续阶段,应适当增加流速,以促进底物、气体传质和生物膜自动更新。
3.2 生物因素
菌-藻共生生物膜的形成过程中,其所释放的EPS起关键作用。在生物膜生长繁殖阶段,EPS具有聚集、连接菌-藻细胞以及作为暂存和传递营养物质媒介的作用,构成了菌-藻共生生物膜的骨架[37]。EPS是由蛋白质、多糖、磷脂和核酸等组成的高分子聚合物,根据其与细胞结合的紧密程度,又分为溶解态EPS(Soluble EPS, S-EPS)和结合态EPS(Bound EPS, B-EPS)。EPS官能团同时包含多个亲水/疏水、正电/负电结合位点,故在菌-藻共生生物膜形成过程中,EPS会对细胞表面理化性质(如带电性、亲疏水性、粗糙度等)产生重要影响[51-55]。Li等[56]研究发现,生物膜生长速率与B-EPS的蛋白质和多糖比值、亲疏水性、含量及表面zeta电位密切相关。其中,EPS中带正电的基团,如-NH2,能增加细胞表面zeta电位,从而促进生物膜的稳定性。此外,生物膜的形成与EPS表面能正相关,而EPS表面能又与EPS表面亲水性呈正相关,与疏水性呈负相关。而Chen等[57]还发现,EPS在载体表面呈非均匀分布,增加了载体表面粗糙度,为生物膜形成创造了有利条件。
另一方面,细菌的生理活动以及其与微藻的相互作用对菌-藻共生生物膜的形成也具有关键作用。细菌增殖速度高于微藻,细菌在快速增殖阶段分泌大量的EPS,粘附在载体表面,形成细菌生物膜,随着细菌丰度和多样性的提高,微藻可利用的碳源和氮磷无机盐增多,促使更多的微藻细胞附着在载体表面,形成菌-藻共生生物膜并促进微藻生长[58]。Hodoki[59]研究证实,在培养藻类生物膜的过程中,引入一定量的细菌能显著提高生物膜中叶绿素含量,并增强微藻细胞的附着性;Holmes[60]研究发现,与无菌的藻生物膜培养体系相比,添加细菌的培养体系里,生物膜中的叶绿素含量增加了1~2个数量级;Irving等[61]研究表明,与无菌培养液相比,在未杀菌的经过二级处理的废水中生长的藻类生物膜厚度增加了9倍。
3.3 载体类型
载体是细菌和微藻形成生物膜的关键性场所,其表面性质对菌-藻生物膜的形成、代谢活性具有重要影响。按照材料性质,通常可将载体分为无机载体和有机载体。目前研究的无机载体包括陶瓷颗粒、玻璃、不锈钢板、石灰石等。有机载体分为天然高分子载体和合成高分子载体,其中,天然有机高分子载体包括海藻酸钙、琼脂、棉、木质载体等;合成高分子材料包括聚四氟乙烯(PVDF)、聚氯乙烯(PVC)、聚乙烯(PE)、聚丙烯(PP)、尼龙、纸、聚氨酯(PU)、聚苯乙烯(PS)、醋酸纤维(CA)、硝化纤维(CN)等。在选择载体时,不仅需要综合考虑载体的表面特性、来源、成本以及耐用性,还应根据载体安装方向对微生物附着性的影响来选择不同的载体,如对于水平安装的载体应选择表面相对光滑的材质,诸如PVC、玻璃材料等,便于后续刮取生物质进行回收,而垂直安装的载体应选择表面粗糙度较大的,如棉材质载体,使微生物细胞能够克服重力作用紧密地粘附在载体表面[62]。
微生物和载体表面最初的相互作用力为静电作用力。细菌和微藻细胞表面通常带负电,易于粘附在带正电的载体表面,而在带负电的载体上附着需要克服静电斥力,所以,通常选取带正电的载体作为菌-藻生物膜附着场所[16]。载体的亲/疏水性也影响微生物附着效果。亲水性载体表面容易吸附微生物粒子,但吸附作用较弱,为可逆吸附;疏水性载体表面吸附力较强,但吸附不可逆。适宜的载体应既能满足微生物生理活动需求,又具有较强的吸附力,因此,选择的载体应达到一定亲/疏水平衡值[63]。Gross等[64]、Sekar等[65]分别研究了微藻细胞在不同载体表面的附着性能,结果表明,微藻更容易在疏水性载体表面附着并形成初始生物层。除上述提及的载体表面性质外,载体的生物毒性以及可生物降解性对菌-藻生物膜形成也具有一定影响,例如,铜板对微藻和细菌具有毒害作用,不适合作为菌-藻生物膜的载体,而纸材质的载体虽然对微生物无毒害作用,但因其可被微生物降解,缩短了载体的使用寿命,同样不适合长期作为菌-藻生物膜载体,所以,应选择对菌-藻细胞无毒害作用、使用寿命长且易于微生物附着的材质作为载体,如棉质、PVC、PE等材质。
4 菌-藻生物膜在污水处理中的应用
基于菌-藻生物膜污水处理体系的独特优势,近年来,不同类型的菌-藻生物膜反应器被开发用于废水处理。由于菌-藻生物膜反应器在供气方式、载体类型等方面具有广泛的选择,因此,反应器设计较灵活,文献中报道了多种构型的菌-藻生物膜反应器,根据反应体系是否直接与外界环境进行物质交换,可将反应器分为开放式反应器和封闭式反应器,其中,开放式反应器为开放容器,可直接与外界环境进行CO2/O2等交换;封闭式反应器为密闭容器,不能与外界环境进行气体交换。常见的开放式光生物反应器构型为水力藻类床(Algal Turf Scrubber,ATS)、菌-藻生物膜转盘等,封闭式生物反应器主要包括封闭管状/平板状菌-藻生物膜反应器。笔者主要介绍菌-藻生物膜系统应用于城镇生活污水、工业废水以及养殖废水处理的研究进展。
4.1 城镇生活污水
城镇生活污水是菌-藻生物膜污水处理技术研究最广泛的废水类型,20世纪80年代,Adey等[66]就利用ATS处理城市生活污水,并获得了较好的氮磷去除效果。此后,关于菌-藻生物膜城镇生活污水净水体系的研究广泛展开。Posadas等[67]对比了封闭管式生物膜反应器和实验室规模的ATS体系对模拟生活污水的处理效果,发现开放式菌-藻生物膜反应器对TN、TP的去除效果分别可达92%±5%和96%±2%,而封闭管式生物膜反应器对TN、TP的去除率分别为80%±6%和68%±18%。应用较广泛的载体为二维平面构型,此类载体结构简单且容易被菌-藻附着,但其比表面积较小,挂膜量较少,不适用于污染物浓度高的废水处理。针对这一弊端,研究者选取了比表面积更大的三维填料作为载体,如唐聪聪[68]将具有多孔结构的球形陶瓷填料(比表面积为90 m2/m3)和菌-藻共生体引入SBR中,构建了序批式菌-藻共生泥膜反应器(A-SBBR),根据填料固定、不固定将反应器分为I型A-SBBR和Ⅱ型A-SBBR,并应用于模拟生活污水处理,研究结果表明,I型TN、TP去除效果分别为65.8%、89.3%;Ⅱ型TN、TP去除效果分别为69.91%、94.78%。据文献报道,Zhang等[10]构建了一种膜曝气菌-藻生物膜反应器,其改变了传统菌-藻生物膜体系的供气方式,利用透气膜组件向反应体系提供O2,成功地在膜片表面培养出菌-藻共生生物膜,其对TN、TP的去除率分别为80.56%、62.54%。
4.2 工业废水
废水中的有毒有害物质对微生物生长繁殖以及生物活性具有强烈的抑制作用,极易影响废水处理效果。相较于传统生物处理技术,菌-藻共生生物膜工艺不仅可以有效去除多种污水中的氮磷等营养元素,同时,藻类具有较强的毒害物质耐受性,对有毒污染物,如重金属、石油烃类等也具有较好的处理效果,因此,也有学者利用菌-藻生物膜处理含毒性物质废水。
重金属污染物是工业废水中常见的难处理污染物,前人广泛开展了微藻对As、Cd、Cr、Pb、Hg等重金属废水处理的研究(如表1所示),证实微藻在耐受重金属毒性并修复重金属污染水体方面具有较大潜力,所以,结合菌-藻共生生物膜在污水处理中的优势,利用菌-藻生物膜法去除水体中的重金属成为一项新兴污水处理技术。菌-藻生物膜法处理重金属污染废水主要依靠吸附作用,大量研究表明,微生物分泌的EPS因含有丰富的含氧官能团,易与金属离子通过络合作用进行吸附。张道勇等[70]研究了丝藻(Ulothrixsp.)-细菌生物膜去除Cd,结果发现,系统中Cd的去除效率与丝藻分泌的EPS相关性良好,此外还发现,丝藻所分泌的EPS为菌-藻共生体提供了一个缓冲Cd毒性的微环境,这使得菌-藻生物膜能在不利的环境中保持较高的活性并能持续有效地去除水体中的Cd。高敏等[71]也探究了菌-藻生物膜对重金属Cd的去除效能,研究发现,Cd的去除效果受温度、pH值以及离子强度影响较大,其中,Cd离子浓度变化范围为2~10 mg/L时,去除率由97.8%增至98.42%,超过10 mg/L时,去除率下降。出现该结果的原因可能是低浓度Cd离子刺激了生物膜分泌EPS,增加了对Cd的吸附,高浓度Cd离子抑制了微生物的活性,降低了EPS的分泌,且存在竞争官能团以及结合位点的现象,故抑制了Cd离子被菌-藻生物膜吸附。梁丽华[72]探究了光照氧化塘菌-藻共生生物膜系统去除煤矿区废水中的Cr(Ⅵ)的潜力,研究结果表明,当Cr(Ⅵ)低于3 mg/L时,去除率大于90%,经过长时间的驯化,在Cr(Ⅵ)进水浓度为20 mg/L的条件下,去除率可稳定在88.2%,证实了菌-藻生物膜对Cr(Ⅵ)具有良好的去除效果。

表1 不同藻种类对重金属的去除效果[69]Table 1 Algal species used in the heavy metal removal
含毒性物质废水中除含有重金属外,还含有难降解有机污染物,有机污染物被微生物矿化降解需要消耗大量O2,传统生物处理工艺需设置曝气装置供氧,不仅能耗高,更易造成有毒物质如酚类污染物、有毒有机溶剂的挥发,产生二次污染,此外,难降解有机污染物易对微生物活性产生抑制效果,降低污水处理效能。近年来,一些学者发现,菌-藻生物膜对工业废水中难降解有机物具有较大去除潜能,并且无须额外供氧,避免二次污染。微藻对细菌降解石油烃类污染物具有促进作用,主要体现在微藻可分泌表面活性剂,以有效提高有机污染物的可生化性,同时,微藻细胞分泌的胞外物质不仅促进了石油烃共代谢,还能为共生菌提供营养物质及较稳定着生环境,增强石油烃降解效率[73]。Mukherji等[74]利用生物转盘培养的菌-藻生物膜处理含石油烃废水,研究发现,在高有机负荷条件下(总石油烃,23.9~47.8 g/m3/d),油滴被吸附在生物膜表面且大部分烷烃被微生物矿化,当HRT大于18 h时,石油烃的去除率大于97%。Chavan等[75]研究了苯酚对菌-藻生物膜转盘修复石油污染水体的影响,结果表明,在HRT为21 h,有机污染物负荷为27.33 g/(m3·d)的条件下,当进水中苯酚含量由0.11 g/(m3·d)增加至0.69 g/(m3·d)时,石油烃去除率由99%降低至94%,证实苯酚对微生物活性有一定的抑制作用,但菌-藻生物膜对苯酚仍具有一定的耐受性,因此,污染物降解活性仍然较高,并对苯酚具有一定的去除效果(18.8%~38%)。Muoz等[76]探究了小球藻sorokiniana和菌种Ralstoniabasilensis构建的菌-藻共生生物膜系统对水中的水杨酸(可抑制细菌活性的有机物)的去除效果,结果表明,在ATS反应器中HRT为1.3 d条件下水杨酸去除率可达100%。
4.3 养殖废水
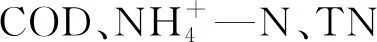
5 结论与展望
菌-藻生物膜既广泛存在于天然水体中的固体介质表面,也可以通过人工强化手段在光生物反应器中培养形成,其中,环境因素、微生物活性以及载体类型是影响菌-藻生物膜形成和脱落的重要条件。相较于传统污水处理工艺,菌-藻生物膜不仅在一定程度上维持高浓度生物量,减少占地面积,同时在污染物去除、节能降耗以及对环境毒性的耐受性等方面也表现出极大的优势。因此,近年来,将菌-藻共生生物膜污水处理技术应用于城镇生活污水、工业废水以及养殖废水的研究方兴未艾。虽然,菌-藻生物膜污水处理体系具有其独特的优势,但在实际工程中的应用仍任重而道远。其中,菌-藻共生生物膜快速形成及高效稳定控制、污染物降解机制及针对特定废水处理的细菌和微藻的富集培养等方面仍有待进一步研究,具体包括:
1)菌-藻生物膜快速形成及稳定高效是完成污水处理的前提和基础,在生物膜形成过程中,细菌及藻类间信号传递,特别是群体感应是影响生物膜形成时效及微生物群落结构的重要因素,而有关如何调控菌-藻间群感效应的研究鲜有报道;另一方面,由于水力剪切力、遮光效应、传质阻力等所导致的菌-藻生物膜非自然脱落,藻细胞生长受限等问题仍未解决。因此,可通过反应器构型设计、运行参数优化等增强藻体光捕捉能力,降低或延缓生物膜脱落频次;同时,将宏观调控与微观的细菌及微藻间信号分子相结合,进一步研究菌-藻共生生物膜快速形成及稳定控制机制。
2)当前有关菌-藻生物膜对毒性物质的耐受机制的理解尚停留在宏观层面,需合理利用基因组学、代谢组学等研究手段,从微观角度剖析菌-藻生物膜在极端环境下的耐受机制和去除机理。
3)目前,有关菌-藻共生生物膜处理普通生活污水的研究报道较多,并有一定的工程应用,而菌-藻生物膜在特种废水中毒害物质的去除方面也展现出巨大潜力,但多处于实验室研究状态。因此,可通过菌种筛选及驯化、微生物富集培养等方式,针对性地投加细菌及藻类来提高菌-藻生物膜在特种废水处理方面的适应能力,并根据微生物及废水特征,合理开发相适应的反应器构型,从而进一步推广菌-藻共生生物膜在特种废水中的工程应用。此外,针对工业废水浊度所造成的藻类太阳能利用率低等问题,可通过结合物化等前处理手段降低进水浊度、色度等,以期减缓水体遮光效应。
