正反杂交对两种生态型少花龙葵F1 代生长及硒积累的影响*
2021-06-11黄科文曹艺兰郑阳霞林立金廖明安
黄科文,曹艺兰,郑阳霞,李 良,刘 继,林立金,廖明安
(1.四川农业大学 园艺学院,四川 成都 611130;2.甘孜藏族自治州农技土肥站,四川 康定 626000;3.成都市农林科学院,四川 成都 611130;4.四川农业大学 果蔬研究所,四川 成都 611130)
硒 (selenium,Se) 是一种必需元素,在动物及人体中发挥着重要的生物学作用,具有抗氧化、防癌、增强人体免疫及生殖能力等多种功能[1]。人体硒含量不足易引起GSH-Px 等抗氧化酶活性的降低,从而增加类风湿性关节炎、心肌病和癌症等疾病发生的风险[2]。中国是典型的缺硒国家,自东北(黑龙江)到西南(云南)地区均属于贫硒地带[3],多地曾爆发过大规模的地方性缺硒症状[4]。适当补硒不仅可以有效防治缺硒病症的发生,还能维持人体免疫系统、内分泌系统以及神经系统的稳态,增强人体的抗性[5]。硒在自然界中赋存多种形态,与无机硒相比,硒代半胱氨酸以及硒代蛋氨酸等有机硒可被人体直接吸收利用,被认为是最理想的硒源[6]。在自然环境中,植物不仅在硒的化学循环中扮演着重要角色,更是将无机硒转化为有机硒的主要承担者。尽管通过添加外源硒肥可以有效提高农产品的硒含量,但据不完全统计,目前可面向市场推广的富硒农产品只有40 余种[7],远远达不到人们对硒的需求。因此,筛选及选育优良的富硒品种以及通过农艺措施增强植物对硒的富集能力是富硒农产品的重要研究方向。
杂交选育多指具有不同遗传性状个体间的有性交配,可以通过双亲基因的重新组合进而综合亲本的优良性状,获得具有杂种优势的新品种(系)[8]。优势杂种的目标性状主要通过其亲本传递,选择性状互补或累加的高亲和亲本杂交可以有效提高育种效率,克服双亲的不利性状[9]。因此,在杂交育种中,亲本的正确选择以及合理的组配是成功获得亲本优良性状的重要基础。生态型是指同一物种受生态适应性、种植模式以及地理隔离等因素的影响表现出具有一定结构或功能差异的不同类群[10]。不同生态型的植株在同一生长环境中仍能保持稳定的遗传差异,以其为杂交材料不仅可以增加其后代的遗传变异还能克服当地品种基因贫乏单一的弱点[9]。王坚等[11]以9 个生态类型的59 份粳稻主栽品种为亲本材料,通过双列杂交试验发现华北粳、日本粳以及韩国粳这3 个生态型材料相互结合可有效提高后代稻米的整精米率,为粳稻亲本的选择提供了参考。朱建荣等[12]发现:利用云南不同生态型甘蔗细茎野生种与甘蔗栽培种杂交对后代经济性状的影响存在较大差异,其中云割83-174、瑞割83-160 及云南1 号野生种可作为蔗糖高糖育种的优良种质。目前,国内外有关富硒农产品的研究主要集中在富硒农作物的资源筛选,并成功选育了谷丰优8312[13]、晋糯8 号[14]和青杂2 号[15]等具有高富硒潜力的作物良种,但有关不同生态型野菜资源的富硒研究尚未见报道。少花龙葵(Solanum photeinocarpum)又名白花菜,为茄科茄属一年生直立草本植物,可作蔬菜食用,其嫩叶富含粗蛋白、粗纤维、维生素以及多种营养元素,被认为是一种具有较高开发价值的野生植物资源[16]。另外,研究表明:少花龙葵地上部分的硒含量可在外源硒处理下超过富硒植物的临界值并对高浓度硒表现出较强的抗逆性,是一种理想的富硒野菜[17]。本课题组前期研究发现:农田生态型和矿山生态型少花龙葵的生长及抗逆性在相同生长条件下具有较大差异,并通过杂交试验发现少花龙葵F1代对镉胁迫的抗逆性表现出较强的杂种优势,为少花龙葵亲本的选择提供了参考[18]。因此,本试验以农田生态型和矿山生态型少花龙葵为亲本,研究正反杂交对2 种生态型少花龙葵F1代生长及硒积累的影响,以期筛选出既能提高少花龙葵后代硒含量又能促进其生长的杂交组合,为富硒野生蔬菜的杂交选育提供参考。
1 材料与方法
1.1 供试材料
2017 年8 月,分别在四川农业大学雅安校区农场(N30°23′,E103°48′)和四川省汉源县唐家山铅锌矿区(N29°24′,E102°38′)收集少花龙葵的种子保存。
四川农业大学雅安校区农场,平均海拔620 m,属亚热带湿润季风气候区,多年平均气温16.2 ℃,多年平均降雨量1 743.3 mm,多年平均日照时间1 035 h,多年平均蒸发量1 011.2 mm[19]。唐家山铅锌矿区,平均海拔890 m。现保有铅锌矿储量145.8 万 t,年采矿10 万 t,已连续开采15年,零星堆积了60 多万t废矿渣。矿区地处北温带与季风带之间的山地亚热带气候区,具有典型的干热河谷气候特点,多年平均气温17.9 ℃,多年平均降雨量741.8 mm,多年平均日照时间1 475.8 h,多年平均蒸发量1 248.2 mm[19]。
供试土壤取自四川农业大学成都校区周围农田,基本理化性质为:pH 7.09,全氮含量为1.50 g/kg,全磷含量为0.76 g/kg,全钾含量为18.02 g/kg,碱解氮含量为94.82 mg/kg,有效磷含量为6.30 mg/kg,速效钾含量为149.59 mg/kg[20]。将土壤风干、碾碎,称取4.0 kg 装于21 cm×20 cm(高×直径) 的塑料盆内,加入分析纯Na2SeO3溶液,使土壤硒含量为10 mg/kg,保持土壤田间持水量的80%,自然放置平衡4 周,期间不定期翻土混合,使土壤充分混合均匀。
1.2 试验方法
2018 年4 月,分别将农田生态型和矿山生态型少花龙葵的种子播种于穴盘中催芽,待少花龙葵幼苗高约5 cm、2 片真叶完全展开时分别移栽至干净土壤中,不定期浇水,保持土壤田间持水量的60%。待少花龙葵进入花蕾期后,选择花冠微露、圆润饱满、色泽泛黄的花苞用镊子去雄后立即套袋,并在去雄的1~3 d 内用蘸取了少量少花龙葵父本花粉的毛笔在去雄母本的柱头上均匀涂抹。杂交后约1 周,当少花龙葵花瓣开始凋谢、幼果成形时去除套袋并做好标记,待果实成熟后及时采收并收集种子保存备用。农田生态型少花龙葵(♀)× 矿山生态型少花龙葵(♂) 的后代记为正交F1代;矿山生态型少花龙葵(♀)× 农田生态型少花龙葵(♂)的后代记为反交F1代;农田生态型少花龙葵自交的后代记为农田生态型F1代;矿山生态型少花龙葵自交的后代记为矿山生态型F1代。
2018 年7 月,分别将少花龙葵正交F1代、反交F1代、农田生态型F1代和矿山生态型F1代的种子播种于穴盘中催芽,待少花龙葵幼苗高约5 cm、2 片真叶完全展开时,分别选择长势一致的幼苗移栽至准备好的含硒塑料盆中,每盆种植少花龙葵4 株,每个处理重复3 次。不定期浇水,保持土壤田间持水量的60%。整个生长过程中不定期交换盆与盆的位置以减弱边际效应的影响,及时去除杂草并防治病虫害,待少花龙葵果实成熟后收获材料,测定相关指标。
1.3 样品收集与测定分析
2018 年8 月,收集少花龙葵从上往下第3~4片功能叶,采用1∶1 乙醇丙酮浸泡法测定叶绿素a、叶绿素b 以及类胡萝卜素含量,采用氮蓝四唑法测定超氧化物歧化酶(SOD)活性,采用愈创木酚法测定过氧化物酶(POD)活性,采用高锰酸钾滴定法测定过氧化氢酶(CAT)活性,采用考马斯亮蓝染色法测定可溶性蛋白含量[21]。之后整株收获少花龙葵并拆分成根、茎、叶,用自来水洗净后再用去离子水冲洗3 次,于110 ℃杀青15 min,80 ℃条件下烘干至恒质量,用电子天平称量、粉碎、过0.15 mm 筛。称取0.50 g 烘干样品,经硝酸—高氯酸溶液消煮至透明后定容,采用氢化物原子荧光光谱法分别测定少花龙葵根、茎、叶的硒含量[22]。
1.4 数据处理
数据采用DPS 进行方差分析(Duncan 新复极差法进行多重比较),并用“平均值±标准差”表示;采用以下公式进行计算:
转运系数=植物地上部分硒含量/根系硒含量;
转运量系数=植物地上部分硒积累量/根系硒积累量;
硒积累量=植物硒含量×植物生物量。
2 结果与分析
2.1 正反杂交对少花龙葵F1 代生物量的影响
由表1 可知:在外源硒处理下,少花龙葵的根冠比及各部位生物量从大到小的顺序均为:正交F1代>反交F1代>农田生态型F1代>矿山生态型F1代。少花龙葵正反杂交F1代根、茎及地上部分的生物量均显著高于2 种生态型少花龙葵(P<0.05)。少花龙葵正交F1代根、茎、叶及地上部分的生物量在所有处理中最大,分别较矿山生态型F1代少花龙葵显著提高了29.84%、37.99%、11.97%和24.96% (P<0.05)。另外,少花龙葵正反杂交F1代的根冠比均高于亲本,说明杂交对少花龙葵F1代根系发育的促进作用更显著。
2.2 正反杂交对少花龙葵F1 代叶片光合色素含量的影响
由表2 可知:在外源硒处理下,少花龙葵叶片的叶绿素a、叶绿素b 以及类胡萝卜素含量从大到小的顺序均为:矿山生态型F1代>正交F1代>农田生态型F1代>反交F1代。矿山生态型F1代叶片的叶绿素a、类胡萝卜素以及叶绿素总量分别较少花龙葵反交F1代显著提高了11.19%、16.19%和10.48% (P<0.05)。各处理少花龙葵叶片的叶绿素b 含量无显著差异(P>0.05)。少花龙葵的叶绿素a/b 从大到小的顺序为:正交F1代>矿山生态型F1代>农田生态型F1代>反交F1代,表明不同杂交组合对少花龙葵后代耐阳性的影响存在差异。
2.3 正反杂交对少花龙葵F1 代抗氧化酶活性及可溶性蛋白含量的影响
由表3 可知:在外源硒处理下,少花龙葵叶片的SOD、POD 和CAT 活性以及可溶性蛋白含量从大到小的顺序均为:正交F1代>反交F1代>农田生态型F1代>矿山生态型F1代。其中,农田生态型F1代少花龙葵的SOD 及CAT 活性均显著高于矿山生态型F1代少花龙葵(P<0.05)。与矿山生态型F1代少花龙葵相比,少花龙葵正交F1代的SOD、POD 和CAT 活性以及可溶性蛋白含量分别显著提高了24.95%、53.09%、35.49%和44.07% (P<0.05)。
2.4 正反杂交对少花龙葵F1 代硒含量的影响
由表4 可知:各处理下少花龙葵根系的硒含量无显著差异(P>0.05)。少花龙葵茎、叶及地上部分的硒含量从大到小的顺序均为:正交F1代>反交F1代>矿山生态型F1代>农田生态型F1代。与农田生态型F1代少花龙葵相比,少花龙葵正交F1代茎、叶以及地上部分的硒含量分别显著提高了24.37%、31.61% 和24.86% (P<0.05)。另外,少花龙葵的转运系数从大到小的顺序为:正交F1代>反交F1代>矿山生态型F1代>农田生态型F1代,表明少花龙葵正反杂交F1代将硒从根系转运到地上部分的能力均强于2 种生态型少花龙葵的F1代。
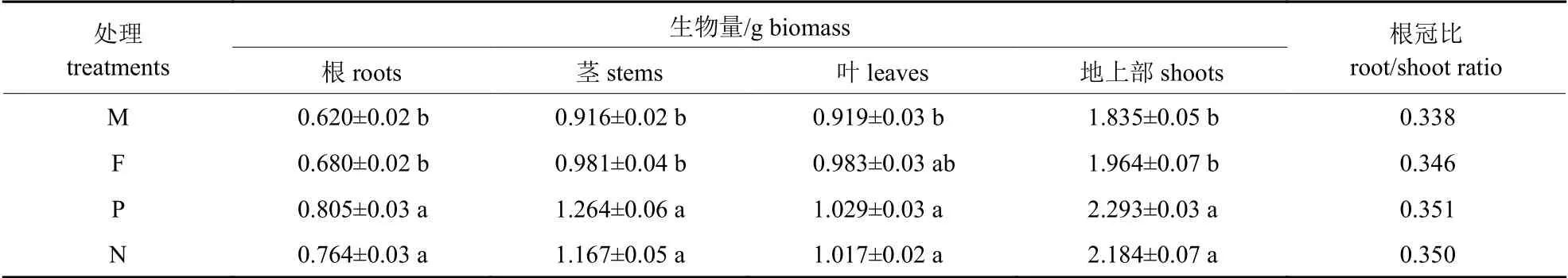
表1 正反杂交对少花龙葵F1 代单株生物量和根冠比的影响Tab.1 Effects of positive and negative hybridization on the biomass per plant in F1 generation and root/shoot ratio of S.photeinocarpum
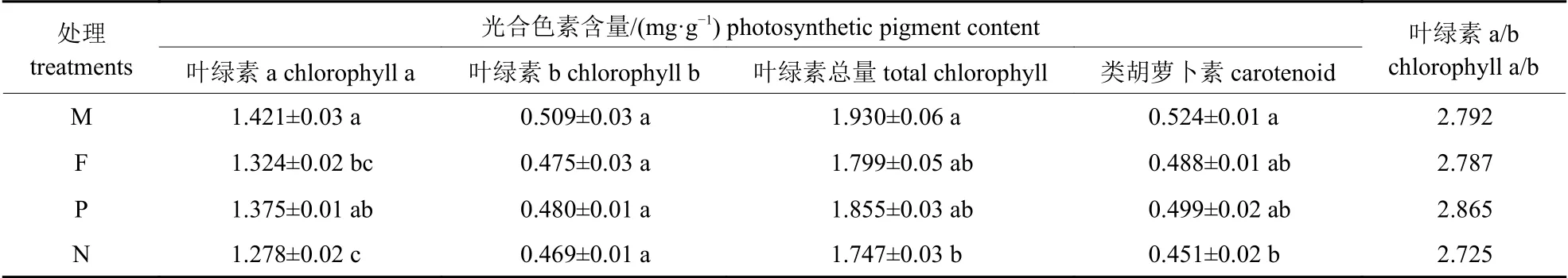
表2 正反杂交对少花龙葵F1 代光合色素含量的影响Tab.2 Effects of positive and negative hybridization on the photosynthetic pigment contents in F1 generation of S.photeinocarpum
2.5 正反杂交对少花龙葵F1 代硒积累量的影响
由表5 可知:少花龙葵茎、叶以及地上部分的硒积累量从大到小的顺序均为:正交F1代>反交F1代>矿山生态型F1代>农田生态型F1代,根系的硒积累量从大到小的顺序为:正交F1代>反交F1代>农田生态型F1代>矿山生态型F1代。少花龙葵正反杂交F1代各部位的硒积累量均显著高于2 种生态型少花龙葵的F1代(P<0.05)。与农田生态型F1代少花龙葵相比,少花龙葵正交F1代茎、叶以及地上部分的硒积累量显著提高了60.28%、37.75% 和45.76% (P<0.05)。少花龙葵正交F1代的转运量系数达到最大值,而农田生态型F1代少花龙葵的转运量系数最低。
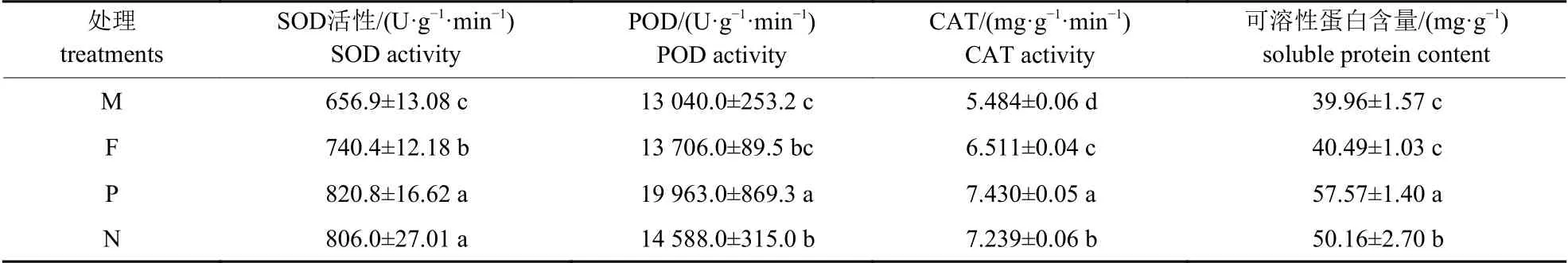
表3 正反杂交对少花龙葵F1 代抗氧化酶活性及可溶性蛋白含量的影响Tab.3 Effects of positive and negative hybridization on the antioxidant enzyme activity and soluble protein content in F1 generation of S.photeinocarpum
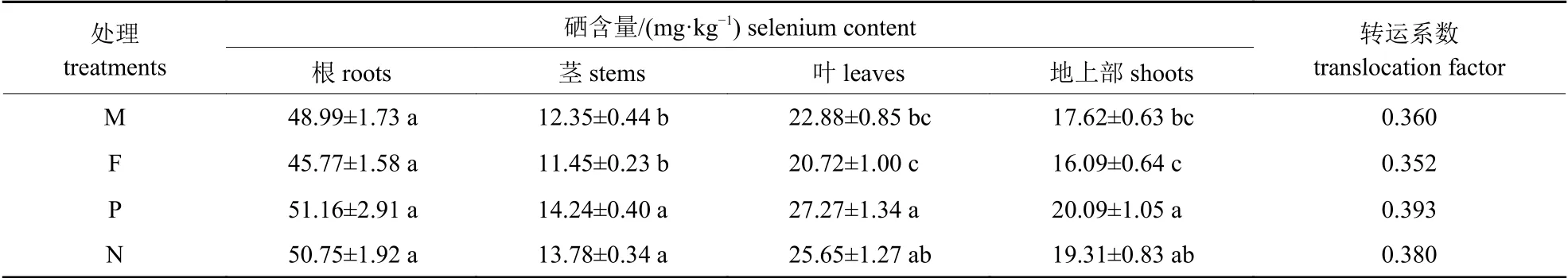
表4 正反杂交对少花龙葵F1 代硒含量和转运系数的影响Tab.4 Effects of positive and negative hybridization on the selenium content in F1 generation and translocation factor of S.photeinocarpum
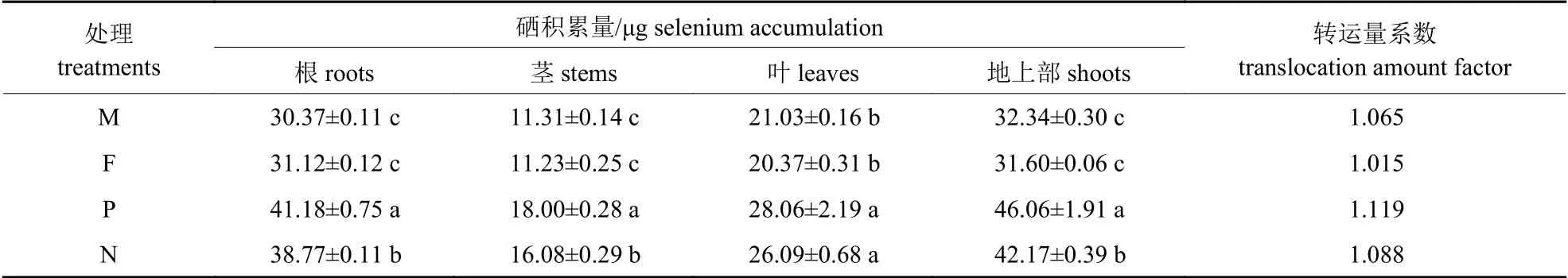
表5 正反杂交对少花龙葵F1 代单株硒积累量和转运量系数的影响Tab.5 Effects of positive and negative hybridization on the selenium accumulation in F1 generation per plant and translocation amount factor of S.photeinocarpum
3 讨论
杂交能够积累亲本中控制同一性状的不同微效基因,可使杂交种在产量、品质以及抗逆性方面表现出较强的杂种优势,培育出生长更旺、代谢更快和抗性更强的杂交后代[23-24]。本试验发现少花龙葵正反杂交F1代根和地上部分的生物量均显著高于其亲本,在生物量上具有显著的杂交优势,与前人的结果[18]相似。光合作用是植物碳同化及干物质积累的基础,而光合色素是限制高等植物光合作用的关键因素[25]。余显权等[26]指出杂交水稻的叶绿素含量与其产量呈正相关关系,暗示了光合色素含量是影响植物杂交后代生长的因素之一。本试验结果表明:2 种生态型少花龙葵的F1代与其正交F1代的光合色素含量均无显著差异,而少花龙葵反交F1代的叶绿素总量和类胡萝卜素含量都低于其他处理,与前人的研究结果[27]不同,可能与植株的遗传背景存在差异有关。
虽然适宜浓度的外源硒可以增强植株的抗逆性[27],但长期暴露在高硒环境下的植株不仅会产生明显的脂质过氧化反应[28],其渗透调节物质的形成也会受到抑制[29]。因此,抗氧化系统和渗透调节物质既是植物抵御高硒环境的基础,也是影响植物生长和硒积累的重要因素。孟杰等[30]发现:耐低锌基因型水稻和锌敏感基因型水稻的杂交后代对低锌环境表现出更强的抗逆性,与其亲本相比,杂交水稻的SOD 活性降幅更小,POD、CAT 及APX 活性显著提高。丁在松等[31]的研究也表明:杂交可以通过提高旱稻后代的抗氧化酶活性来改善植株对渗透胁迫的抗逆性。本试验发现:2 种生态型少花龙葵的正反杂交F1代在抗逆性状中均表现出了超亲优势。另外,农田生态型F1代和正交F1代少花龙葵的抗氧化酶活性和可溶性蛋白含量分别高于矿山生态型F1代及反交F1代。这表明与矿山生态型少花龙葵相比,农田生态型少花龙葵对硒的抗逆性更强,并且以其作为母本杂交对少花龙葵F1代抗逆性的促进作用更好,这可能是因为细胞质遗传效应在杂交过程中发挥了一定作用,使其表现出一定的母系遗传倾向。农田生态型和矿山生态型F1代少花龙葵的各部分硒含量均无显著差异,而正反杂交F1代地上部分的硒含量及转运系数均高于亲本。这表明杂交有利于少花龙葵F1代对土壤中硒的吸收和转运,其相关机制有待进一步深入研究。硒的积累量不仅可以真实反映植物对土壤中硒的富集能力,也是评价富硒农产品的重要参数。本试验结果表明:少花龙葵正反杂交F1代根、茎、叶的硒积累量均显著高于亲本,其中少花龙葵正交F1代地上部分的单株硒积累量最大,达到46.06 μg。说明以2 种生态型少花龙葵为亲本杂交对其F1代的富硒能力具有明显的杂交优势,且以农田生态型少花龙葵为母本的杂交组合对其F1代富硒能力的提升效果更好。
4 结论
在硒含量为10 mg/kg 的土壤中,以农田生态型和矿山生态型少花龙葵为亲本杂交使其正反杂交F1代在生长、抗逆性以及硒积累方面均表现出了超亲优势。少花龙葵正交F1代的生物量、光合色素含量、抗氧化酶活性、可溶性蛋白含量和硒含量均不同程度高于少花龙葵反交F1代,表明以农田生态型少花龙葵为母本、矿山生态型少花龙葵为父本的杂交组合对其F1代的生长及富硒能力具有更强的提升作用,可为高富硒少花龙葵的选育工作提供理论依据。
