粤港澳大湾区不同恢复模式植物群落结构与生态系统健康的动态变化
2021-06-03汪雁佳陆宏芳林永标周丽霞蔡洪月周笛轩
汪雁佳, 陆宏芳, 林永标, 周丽霞, 蔡洪月, 周笛轩
1 中国科学院华南植物园 海岛与海岸带生态修复工程实验室, 广州 510650 2 中国科学院大学, 北京 100049 3 南方海洋科学与工程广东省实验室(广州), 广州 511458
粤港澳大湾区是我国改革开放的前沿,区域人口承载量巨大,也是经济最为活跃的地区之一,在我国经济版图中占有极其重要的地位(占全国 GDP 总量的 12%)。与此同时,城市化和产业化快速发展使大湾区原生生态系统长期遭受严重的人为干扰,形成大面积的丘陵荒坡和退化生态系统,环境污染、水土流失、土地退化和生物多样性锐减等生态问题日益加剧[1]。1985 年广东开展的“五年消灭荒山、十年绿化广东”,以及随后开展的“新一轮绿化广东大行动”,和十八大以后兴起的生态文明建设、美丽中国建设等都促进了区域植被生态恢复工作。经不懈地努力恢复,到 2018 年广东森林覆盖率达 58.59%(广东省林业局),初步在植被恢复、美化绿化方面取得了较好成效,同时在固碳增汇等方面为区域社会经济的发展提供了重要的生态保障[2]。然而,除香港外,大湾区森林植被 58% 以上为人工林,且以针叶林和外来种纯林为主,尤其是桉树人工林,其大规模种植所引起的生态问题争议不断;现存林分普遍存在树种单一、结构简单、天然更新能力较差,生态服务功能弱的问题[3]。“绿色发展,保护生态”是《粤港澳大湾区发展规划纲要》所列基本原则之一;森林作为绿色屏障,在保障大湾区生态环境方面起着至关重要的作用,如何在加快大湾区经济发展的同时牢固树立和践行“绿水青山就是金山银山”的理念,科学有效地恢复和调整区域植被生态系统结构,提升生态系统健康水平和服务功能,推进大湾区经济和生态保护协调发展,是亟待解决的战略问题。
植被恢复目标或成功标准的内核是恢复植被的合理结构、功能和动态过程,进而持续、高效地为人类提供生态系统服务[4]。已有研究表明,相对于外来速生树种,乡土阔叶树种虽然成林较慢,但成林后具有保水固土能力强、物种丰富度高等优势;而物种多样性是生态系统稳定性的基础,因此以乡土树种为主的人工林结构更加稳定[5]。近年来,乡土树种及其混交林的营造在大湾区植被恢复实践中日受青睐[6- 7]。然而,不同物种丰富度的乡土树种人工林植物群落结构、健康与土壤理化性状的发展规律如何?与以桉树为代表的外来种人工纯林以及自然恢复模式相比有何异同?人为启动乡土种人工林的物种丰富度以多少为适宜?一系列问题尚有待长期定位研究的揭示。
针对上述问题,为更好地指导和促进粤港澳大湾区乃至整个华南地区的退化生态系统恢复工作,选取华南地区最具代表性的丘陵荒坡为研究对象,通过大规模森林生态系统野外控制实验和长期定位观测手段,开展生态恢复实验;于 2005 年在鹤山森林生态系统国家野外科学观测研究站人为构建了以红锥纯林(Castanopsishystrixmonoculture,CM)为代表的乡土树种人工纯林、10 种和 30 种两个物种丰富度的乡土树种混交人工林(10 native species mixed forest plantation,10NS;30 native species mixed forest plantation,30NS)、以尾叶桉纯林(Eucalyptusurophyllamonoculture,EU)为代表的外来种人工纯林等植被恢复模式,并以自然恢复灌草坡(Shrub and herb,SH)为对照,就不同恢复模式的植物群落结构、生态系统健康水平和土壤理化性状开展长期定位监测研究。
1 试验地概况
鹤山森林生态系统国家野外科学观测研究站共和样地(112°50′E,22°34′N),为典型的低山丘陵地貌,海拔在 10—60 m 之间;属南亚热带季风气候,气候温暖多雨,年均气温 21.7 ℃,年均降水量 1700 mm,具有明显的干湿季节交替现象;地带性土壤为砖红壤性赤红壤,土壤质地一般为粘壤土类和壤质粘粒,表层土(0—10 cm)pH 值 3.63—4.35,属强酸性土壤。区域顶级群落是亚热带季风常绿阔叶林,由于早期受人为干扰,植被退化严重,形成大面积的丘陵荒山。于 2005 年在退化荒坡上人工构建了乡土种红锥与外来种尾叶桉两个纯林、10 种与 30 种两个物种丰富度的乡土混交林以及自然恢复灌草坡,运用植被与土壤的长期野外定位监测与室内分析手段,量化研究各植被恢复生态系统结构的发展动态。在该野外控制实验平台选取了上述 5 种植被恢复模式,每种模式设有 3 个重复样地,样地采取完全随机分布模式,每个样地占地约 1 hm2;种植株行距为 2 m×3 m,种植密度约为 1667 株/ hm2(图1)。2018年,5 种恢复模式乔木层植被种类变化情况见表1。13龄时,两个混交林共存的乡土树种(人为种植)有 6 种,分别为华润楠(Machiluschinensis)、枫香(Liquidambarformosana)、阴香(Cinnamomumburmannii)、灰木莲(Manglietiaglauca)、红锥(Castanopsishystrix)、黧蒴锥(Castanopsisfissa),另外 2 种共存的乡土树种深山含笑(Micheliamaudiae)与火力楠(Micheliamacclurei)在 10 树种混交林中能够自然萌生。
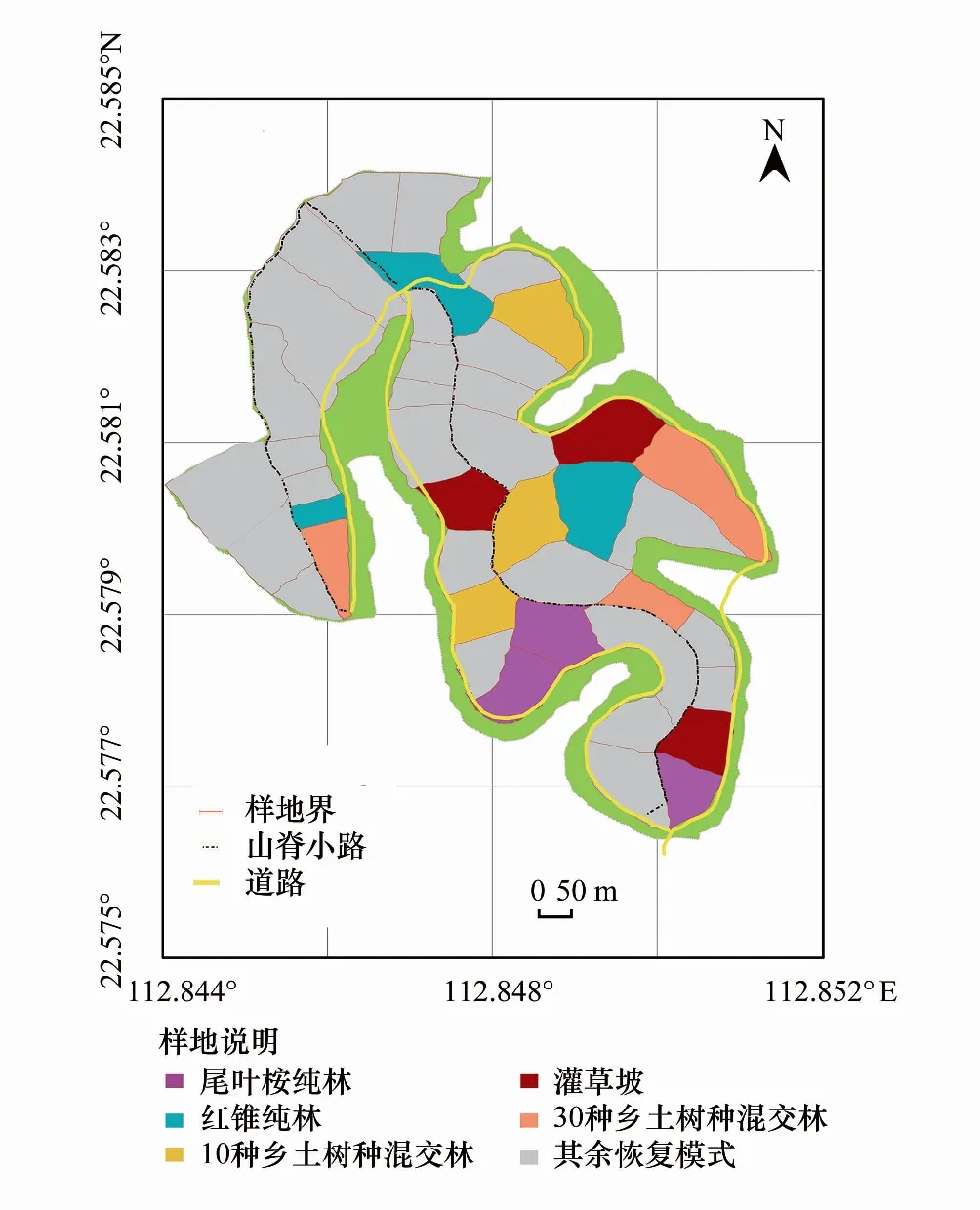
图1 实验样地分布Fig.1 Experimental ground distribution

表1 5种植被恢复模式乔木层植被种类变化
2 研究方法
生物量、生物多样性和土壤理化性状是陆地生态系统的基本特征,被普遍认为是生态系统健康的基础,因此提高植被生物量和物种多样性,进而实现植被与土壤的协同发展,是陆地退化生态系统恢复与重建的重要目标[2]。然而,生物量、物种多样性和土壤理化性状与生态系统健康水平之间的动态量化关系尚有待揭示。针对这一问题,本研究在分析测度各植物群落生物量、物种组成与多样性的同时,引入生态(eco-exergy)概念,量化揭示各植物群落的生物热力学健康水平与发展规律;同时分析植物群落结构和生物热力学健康水平与土壤理化性状间的相关关系,为区域生态恢复实践提供科学依据。
2.1 植被调查与群落结构分析
本研究采用 1 龄(2006年)、4 龄(2009 年)、6 龄(2011 年)和 13 龄(2018 年)的植物群落调查数据。调查方法为相邻格子法,即在每个样地内布设 30 m×30 m 的样地,然后将之平均划分成 9 个 10 m×10 m 网格,调查上述网格内所有乔木的种类、胸径、高度和冠幅;从上述 9 个网格中各随机抽取 1 个 5 m×5 m 网格,共 9 个小网格,调查小网格内所有灌木的种类、基径、高度和冠幅以及草本植物的种类、丛(株)数与盖度。乔、灌、草三层植物种类划分主要参照依据为《鹤山植物志》[8]以及《中国植物志》(http://www.iplant.cn/frps)。2018年采用收割法获取草本层主要物种芒萁(Dicranopterisdichotoma)和乌毛蕨(Blechnumorientale)的生物量;乔、灌木优势种和常见种生物量计算主要采用广州鹤山及其他生态恢复相关研究论文[9- 10]中的生物量模型(表2),其他乔木与灌木物种生物量模型来源于广东鼎湖山站(表3),其他灌木层生物量模型来源于云南西双版纳站(表4),蕨类等物种生物量模型主要来源于湖南会同站(表5,各站资料均来源于国家生态科学数据中心:http://rs.cern.ac.cn/)。
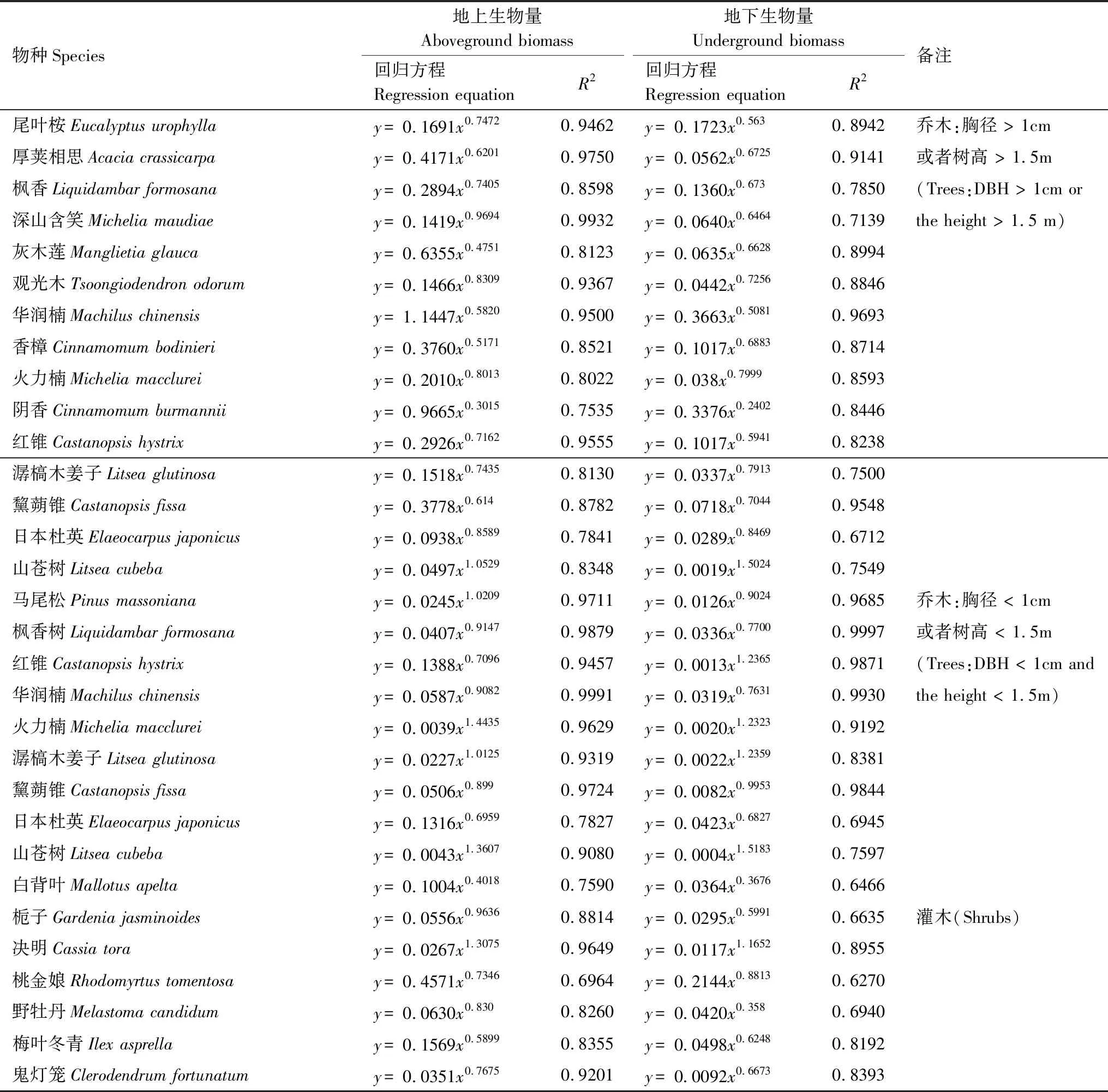
表2 主要乔木及灌木异速生长方程
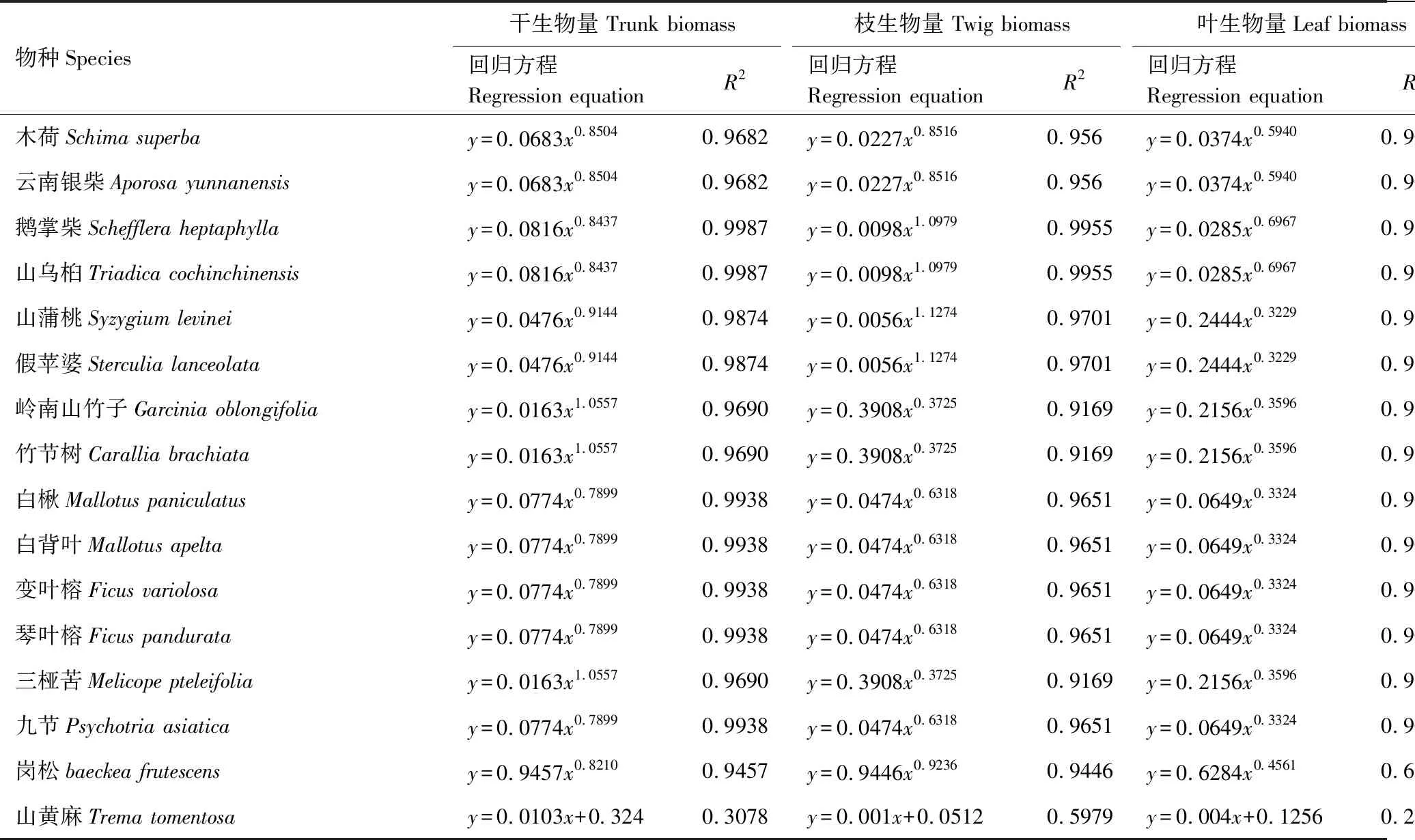
表3 其他乔木及灌木异速生长方程

表4 其他灌木层异速生长方程

表5 蕨类异速生长方程
Shannon-Wiener指数H′(Shannon-Wiener index,SWI)能较好兼容物种丰富度与均匀度,作为物种多样性水平的综合度量指数[11],被广泛运用于植物群落的分析中。除 Shannon-Wiener 指数外,本研究还选取了物种丰富度(species richness,RI)和均匀度指数J(Pielou index,PI)以更全面地分析各植物群落物种多样性。具体计算公式如下:
(1)
J=H′/lnS
(2)
式中,S为样方的植物物种数;Pi为种i的个体数占所有种的个体数的比例。
2.2 群落生物热力学健康指标
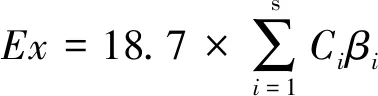
(3)
β=1+ln(201.63×108c)/7.43×105
(4)
其中,c值是生物体单倍体基因组 DNA 的物理总长度(pg)。本研究c值数据来源于英国皇家植物园植物c值数据库(https:// Cvalues.science.kew.org/search);所引用的大部分c值数据精确至属水平,不能精确至属的,则采用同科植物的平均c值。
(5)
2.3 土壤采集和分析
与地上植被调查相对应,各植被恢复模式土壤样品分别采集于 2006 年、2009 年、2011 年和 2018 年的干季。采样方法是分别在上述 15 个实验样地中用直径 3 cm 土钻随机采集 0—10 cm 的土样 5 个,混合后过 10 目土壤筛后装进自封袋中,形成一个混合样;每块样地取 3 个混合样共计 45 个混合样。带回实验室后,一部分湿土用于土壤含水量(SMC)与 pH 的测定,土壤含水量采用烘干法测定,土壤 pH 采用电位法测定;剩余部分风干后研磨,过 100 目土壤筛,用于土壤有机碳(SOC)与土壤全氮(TN)的测定,土壤有机碳采用重铬酸钾氧化-外加热法测定,土壤全氮采用半微量开氏法测定。
2.4 数据处理与分析
数据经过 Excel 2013 软件处理后,在 SPSS 23.0 中采用单因素方差分析(One-way ANOVA)与多重比较(Tukey 检验或 Kruskal-Wallis H 检验,后者在非正态分布或方差不齐情况下使用)分析不同恢复阶段与不同恢复模式植物群落结构特征、生物热力学健康水平和土壤理化性状之间的差异性。图中数据均采用 3 个样地的平均值与标准差表示,显著性水平设定为α=0.05。采用冗余分析(Redundancy analysis,RDA)法[24]分析植物群落结构和生物热力学健康水平与土壤理化性状间的相关关系。其中,RDA 分析使用 R 语言软件完成;其他图件均采用 Origin 2017 软件制作。
3 结果与分析
3.1 5 种恢复模式植物群落生物量动态与比较
3.1.15 种恢复模式植物群落生物量发展动态
研究期间,自然恢复灌草坡(SH)植物群落与各层生物量均无显著变化(图2);4 种人工林中,尾叶桉纯林(EU) 前期生物量增长最快,在 6 龄时已显著高于其1龄时的水平,6龄后其生物量增长放缓;两种乡土混交林(10NS、30NS)前期(6龄前)生物量增长较慢,但发展到13龄时两种乡土混交林生物量也已显著高于其各自1龄时的水平;红锥纯林(CM)生物量增长速度最慢,截止13龄时仍与其1龄时的水平无显著差异。4种人工林乔木层生物量增加趋势与群落总体生物量增加趋势一致,且13龄的EU、10NS与30NS乔木层生物量显著高于其各自1龄时的水平。4种人工林灌木层生物量表现为逐渐减少的趋势,13龄的CM和10NS灌木层生物量分别显著低于其各自1龄与4龄时的水平;而草本层生物量经过13年的发展则与其各自1龄时的水平无显著差异。
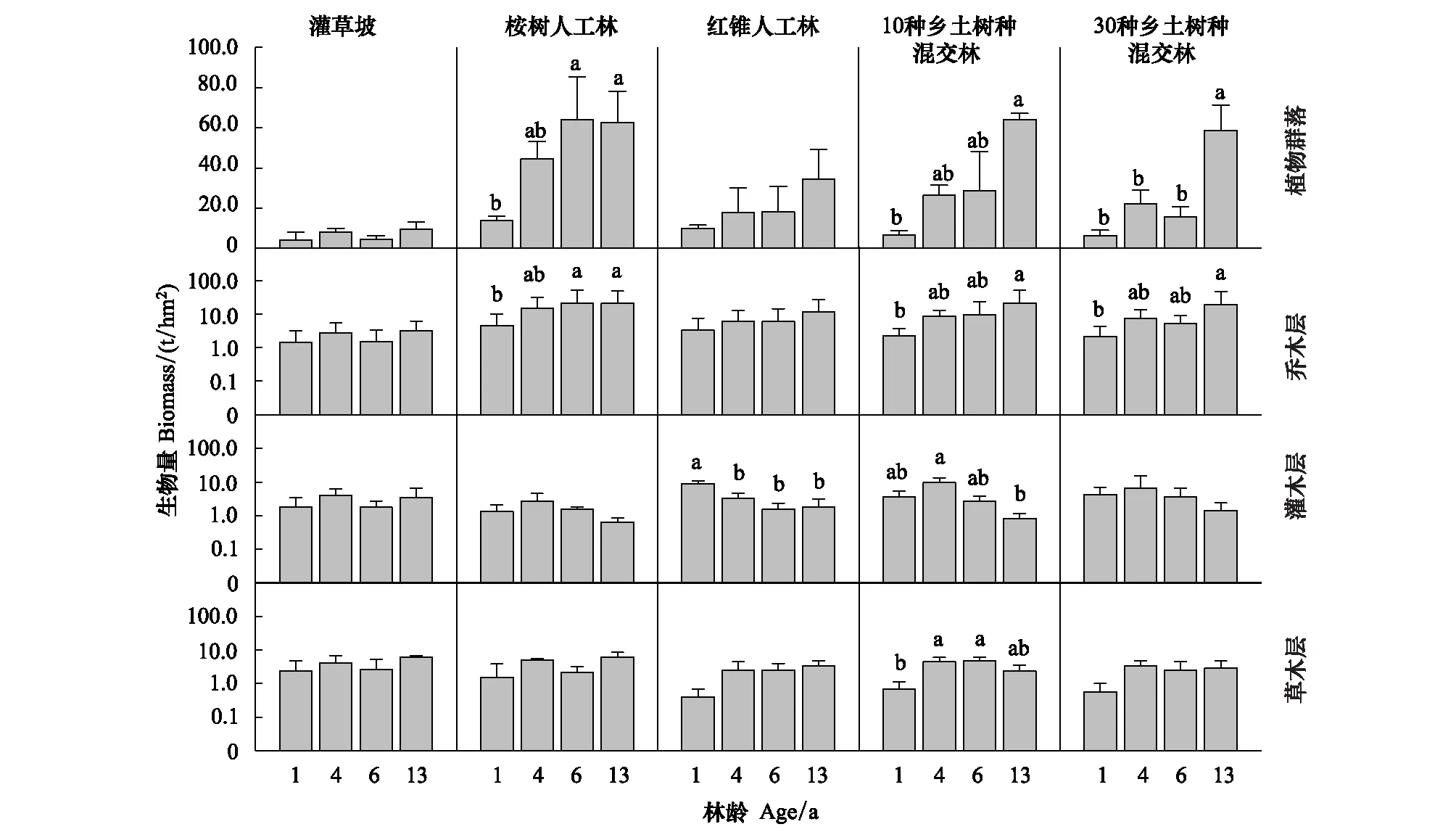
图2 5 种恢复模式植物群落生物量发展动态Fig.2 Dynamics of plant community biomass in the five restoration modes图中小写字母表示数据之间的差异达到显著性(P<0.05);图中数据为平均值±标准差(n=3)
3.1.25 种恢复模式植物群落间生物量动态比较
13龄时,EU 与两种乡土树种混交林植物群落生物量均显著高于自然恢复灌草坡;其中 EU 植被生物量在 1—6 龄时始终保持相对高位,而两个混交林后期发展速度加快,至 13 龄时,已与 EU 无显著差异,且10NS 生物量略高于 EU(图3)。分层结果显示,1—13 龄期间,5 种恢复模式灌木层和草本层生物量均无显著差异。整个研究期间,4 种人工林乔木层生物量显著高于自然恢复灌草坡;EU 乔木层生物量只在 1 龄时显著高于 3 种乡土种人工林;13 龄时 EU 乔木层生物量略高于 CM 和 30NS,略低于 10NS(图3)。
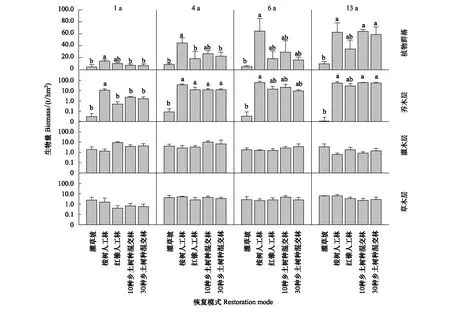
图3 5 种恢复模式间植物群落生物量的动态比较Fig.3 Dynamic comparison of plant community biomass among the five restoration modes
3.2 5 种恢复模式植物群落物种多样性发展动态与比较
3.2.15 种恢复模式植物群落物种多样性发展动态
相较于1 龄时的物种丰富度水平,5 种恢复模式的物种丰富度在 13 年后均有所下降,但只有 30NS 的物种丰富度下降达到显著性水平(图4)。分层结果显示,30NS 乔木层与草本层物种丰富度随着林龄的增加呈下降趋势,且在 13 龄时的物种丰富度显著低于其 1 龄时的水平。各个恢复模式灌木层物种丰富度在 13 年间均无显著变化。
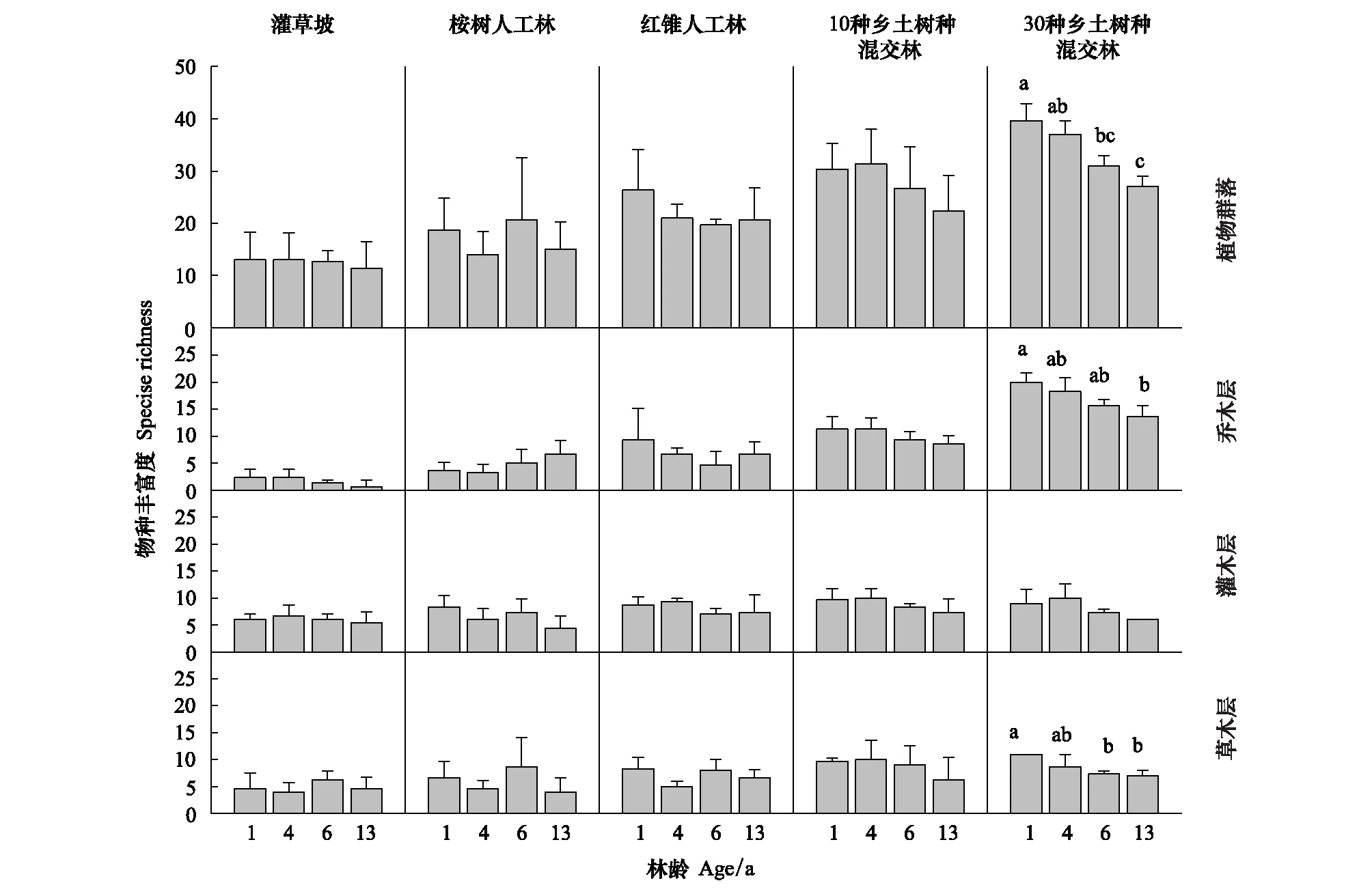
图4 5 种恢复模式植物群落物种丰富度发展动态Fig.4 Dynamic of plant community species richness in the five restoration modes
在 6—13 龄间,5 种恢复模式植物群落 SWI 与 PI 均呈下降趋势;其中 30NS 的降幅最大, 13 龄 时其物种多样性水平(SWI 与 PI)均显著低于其 1 龄与 6 龄时的水平。分层结果显示,13 龄时 30NS 乔木层 SWI 显著低于其 1 龄时的水平(图5)。研究期间,5 种恢复模式灌木层与草本层植被 SWI 和 PI均无显著变化。

图5 5 种恢复模式植物群落物种多样性发展动态 Fig.5 Dynamics of plant community species diversity in the five restoration modes
3.2.25 种恢复模式间植物群落物种多样性动态比较
13 年间,5 种恢复模式植物群落物种丰富度主要表现为混交林>纯林>对照灌草坡。1 龄时,30NS 群落物种丰富度显著高于 EU 和 SH;发展到 13 龄时,30NS 植物群落的物种丰富度显著高于 SH;6 龄以后,纯桉林与 3 种乡土种人工林在物种丰富度上已无显著差异。乔木层结果显示,整个研究期间两种乡土混交林物种丰富度均显著高于 SH;两种人工纯林间则无显著差异;发展至 13 龄时,灌草坡显著低于 4 种人工林,两种纯林显著低于 30NS;两种混交林间亦已无显著差异。整个研究期间,灌木层物种丰富度无显著变化。1 龄时 30NS 草本层物种丰富度显著高于 SH,但 6 龄以后不同恢复模式草本层物种丰富度无显著差异。
与物种丰富度一致,13 年间,5 种恢复模式植物群落 SWI 和 PI 也主要表现为混交林>纯林>对照灌草坡(图7)。单因素方差分析结果显示,灌草坡植物群落的 SWI 在 1 龄时显著低于 30NS,但与其他 3 种恢复模式无显著差异,6 龄以后显著低于两种混交林;10NS 植物群落在 13 龄时超越 30NS,成为 5 种恢复模式中植物群落 SWI 和 PI 最高的模式。除在 4 龄时 EU 的植物 SWI 显著低于30NS 外,此后 4 种人工林恢复模式植物群落 SWI 均无显著差异。1—6 龄间,5 种植被恢复模式的群落 PI 均无显著差异;发展到 13 龄时,灌草坡的 PI 与纯桉林无显著差异,但显著低于 3 种乡土树种人工林;且 10NS 显著高于 EU。
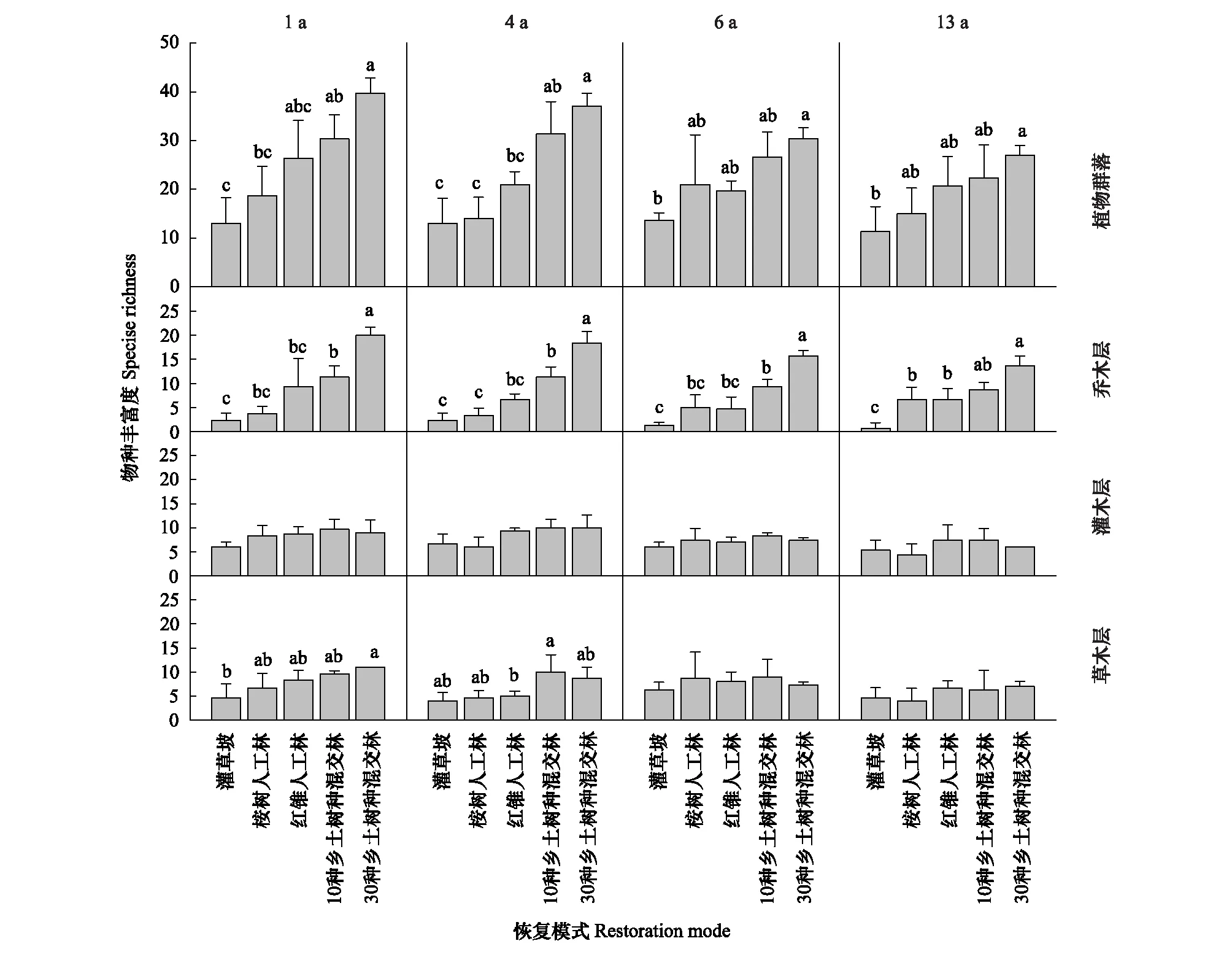
图6 5 种恢复模式间植物群落物种丰富度的动态比较Fig.6 Dynamic comparison of plant community species richness among the five restoration modes

图7 5 种植被恢复模式间植物群落物种多样性的动态比较Fig.7 Dynamic comparison of plant community species diversity among the five restoration modes
分层结果显示,1 龄时只有 30NS 乔木层的 SWI 显著高于 EU;而发展到 13 龄时,两种混交林的乔木层 SWI 均显著高于其他三种恢复模式;各恢复模式乔木层 PI 均无显著差异。研究期间 5 种恢复模式间灌木层和草本层植物群落 SWI 和 PI 均无显著差异。
3.3 5 种恢复模式植物群落生物热力学健康水平发展动态与比较
3.3.15 种恢复模式植物群落生物热力学健康水平发展动态
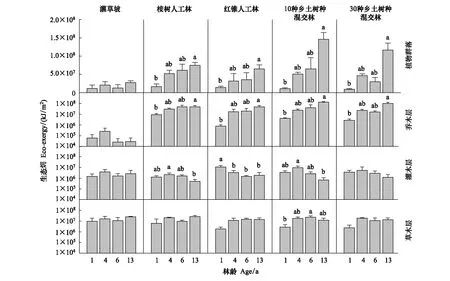
图8 5 种植被恢复模式植物群落生态发展动态Fig.8 Dynamics of plant community eco-exergy in the five restoration modes
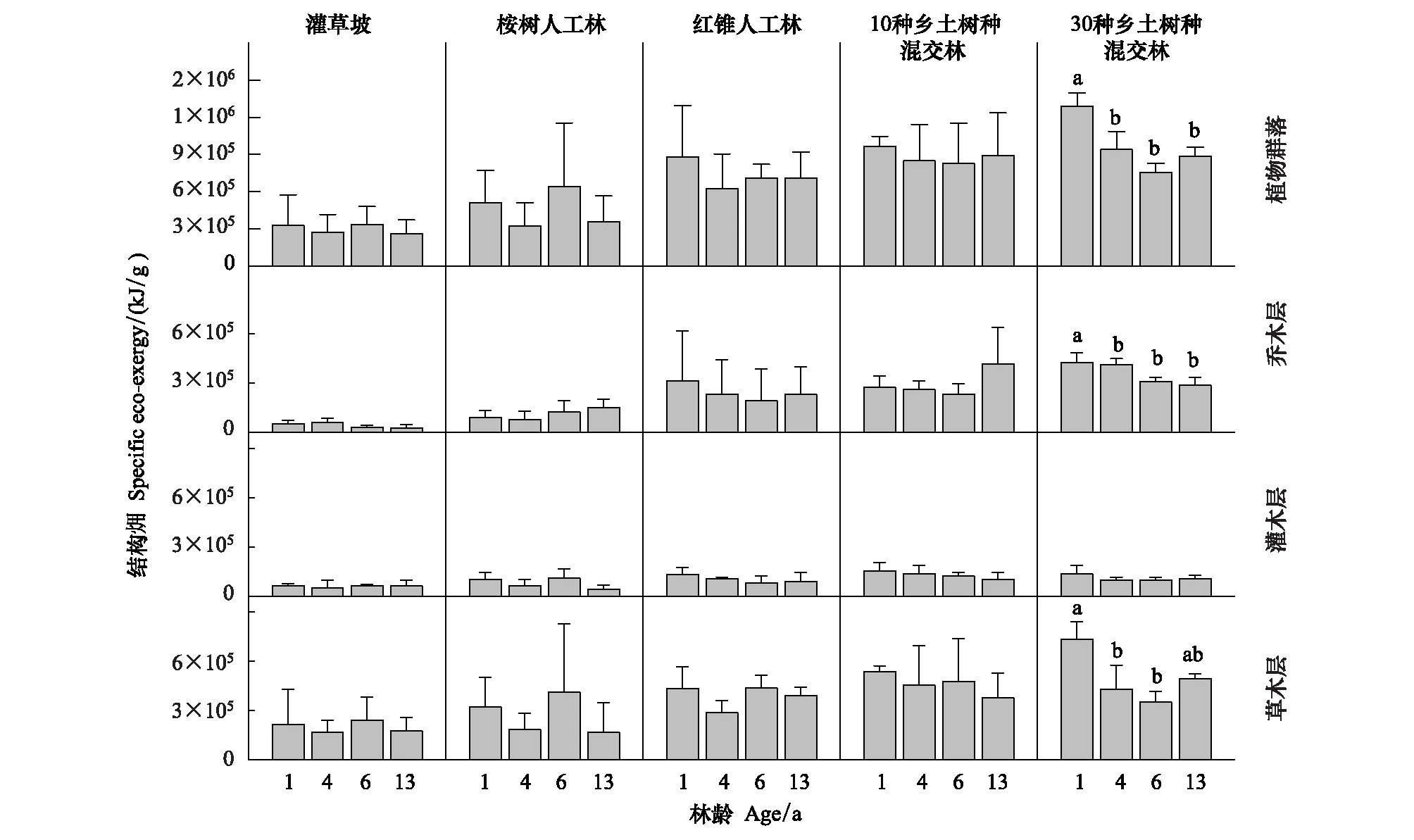
图9 5 种恢复模式植物群落结构发展动态Fig.9 Dynamics of plant community specific eco-exergy in the five restoration modes
3.3.25 种恢复模式间植物群落生物热力学健康水平的动态比较
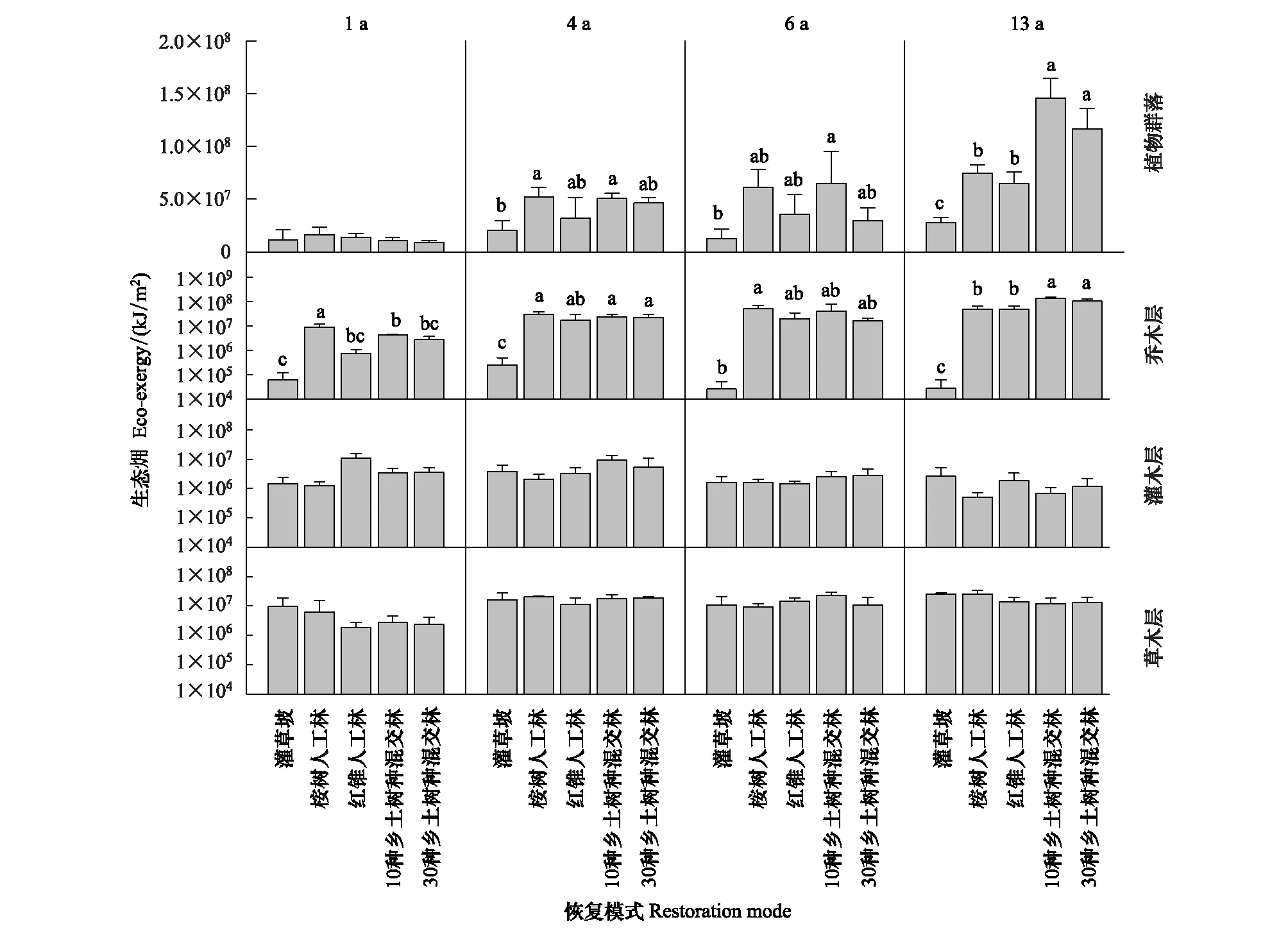
图10 5 种恢复模式间植物群落生态动态比较Fig.10 Dynamic comparison of plant community eco-exergy among the five restoration modes
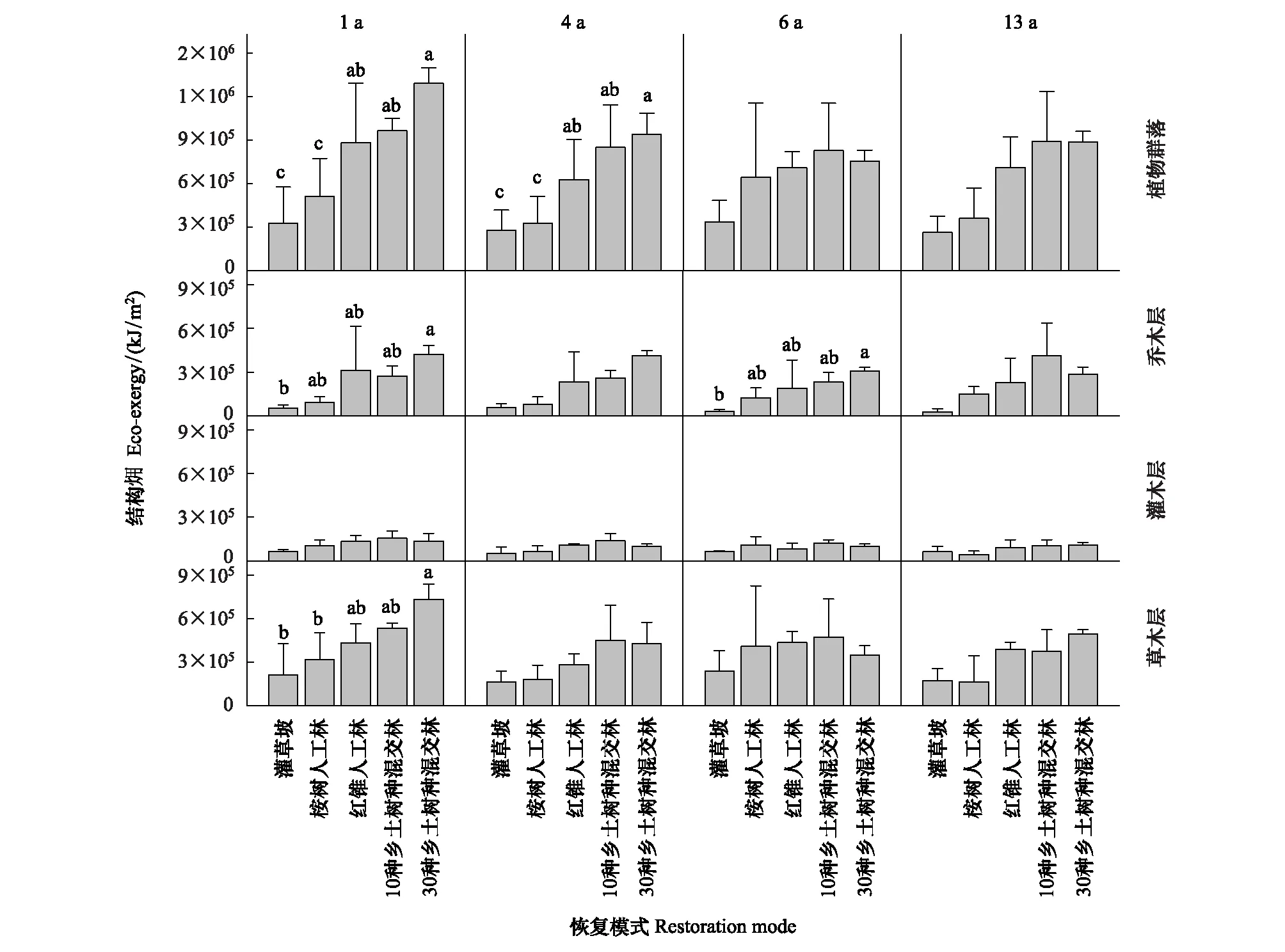
图11 5 种恢复模式间植物群落结构动态比较Fig.11 Dynamic comparison of plant community specific eco-exergy among the five restoration modes
3.4 5 种恢复模式土壤理化性状发展动态与比较
研究期间,5 种恢复模式土壤酸碱度(pH)、含水量(SMC)、有机碳含量(SOC)、全氮含量(TN)和碳/氮比(C/N)均呈现(波动)上升趋势。13 龄时,5 种恢复模式的 SMC 均显著高于其各自在1龄时的水平;两个混交林和自然恢复灌草坡的土壤pH显著高于其各自在1龄时的水平。5 种恢复模式的 SOC 均在 1—4 龄期间略有下降,而后快速提升;6 龄时除 EU 外的 4 种恢复模式 SOC 均显著高于其各自在 4 龄时的水平;之后 SOC 的增长速度放慢甚至下降;13 龄时 CM 的 SOC 已与其 1—4 龄的水平无显著差异,EU 的 SOC 呈现出与两种混交林相似的变化趋势,但差异不显著。研究期间,只有 30NS 的土壤 TN 在 13 龄时显著高于其 4 龄的水平,只有 CM 的土壤 C/N 在13 龄时显著高于其 1—6 龄的水平;其他模式的土壤 TN 和 C/N 均无显著变化。红锥纯林的土壤 C/N 在 13 龄时显著高于其 1—6 龄时的水平。整个研究期间,5 种恢复模式间土壤 pH、含水量、有机碳与碳氮比均无显著差异。13 龄时,只有 10NS 的土壤全氮(TN)显著高于 SH 和 CM。
3.5 植物群落结构与生物热力学健康水平和土壤因子间的相关关系
RDA 分析结果显示,在1—6 龄时各层植物群落结构和生物热力学健康水平与土壤环境因子间无显著相关关系。但随着时间的推移,土壤环境数据的解释度由 1 龄时的 73.3% 上升至 13 龄时的 82.0%,植被与土壤间的相关关系逐步建立(图13)。13 龄时的冗余分析结果表明,各层植物群落结构和生物热力学健康水平与土壤环境因子接近显著相关关系(P<0.1),且第一轴解释了54.7% 的方差变异,第二轴解释了 27.3% 的方差变异;进一步分析结果表明,SOC 对地上群落结构方差变化的贡献最大且达到显著水平(F=2.713,P=0.042)。
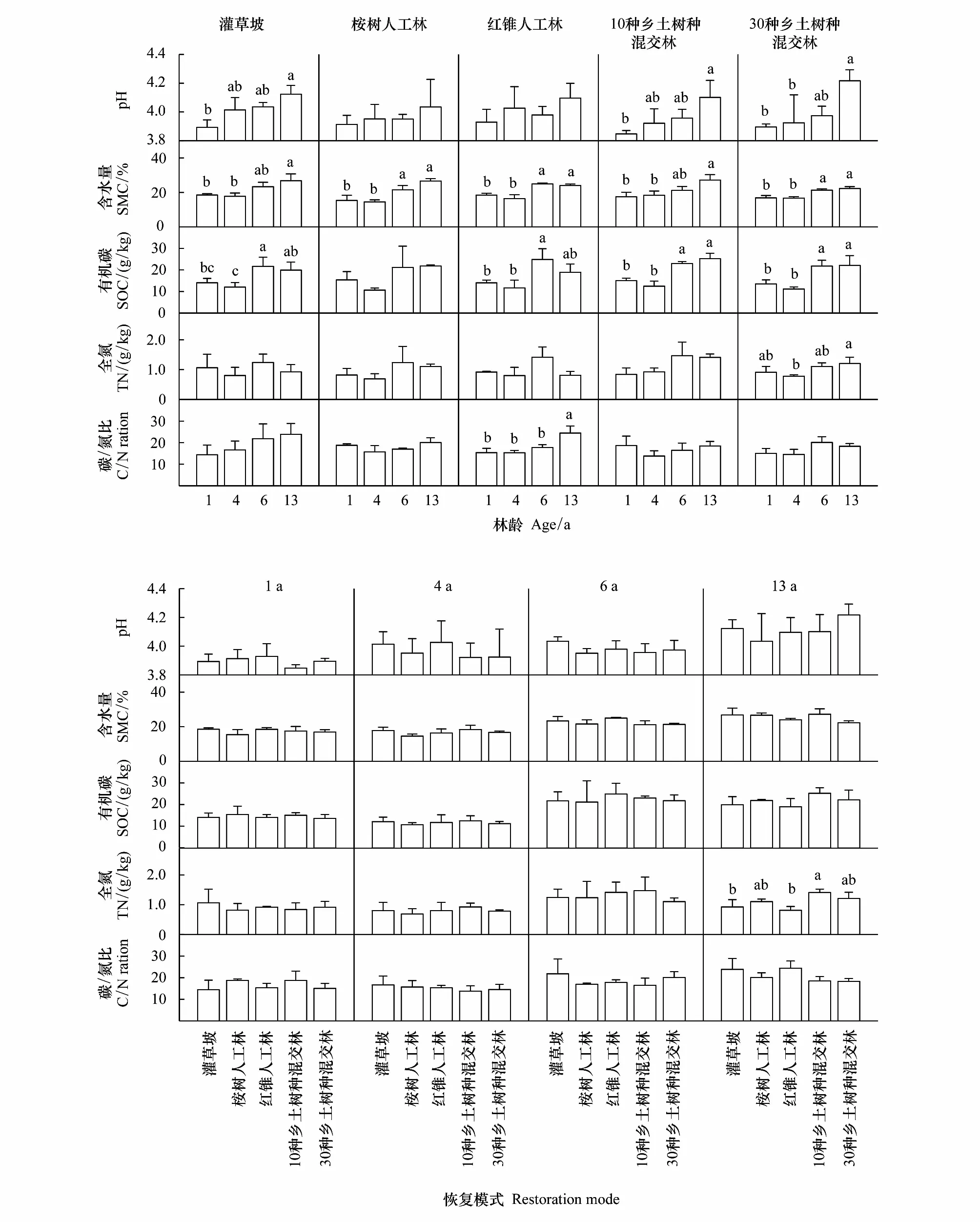
图12 5 种恢复模式土壤理化性质Fig.12 Soil properties of the five restoration modes

图13 不同林龄植物群落结构和生物热力学健康水平与土壤理化因子间的冗余分析(RDA)Fig.13 Redundancy analysis results of plant community structures, bio-thermodynamic health states and soil properties at different forest ageRI: 丰富度指数species richness;PI: Pielou 指数Pielou index;SWI: Shannon-Wiener 指数Shannon-Weiner index;Bio: 生物量biomass,Ex: 生态eco-exergy,Exsp: 结构specific eco-exergy,SMC: 土壤含水量soil moisture content;SOC: 土壤有机碳soil organic carbon;TN: 全氮total nitrogen content
4 讨论
退化生态系统自然恢复是一种被动的恢复策略[25],通常需要经过漫长的发展历程,而利用人工林恢复模式则可在较短时间内促进植被生长与生物量累积[26- 27]。本研究结果表明,研究期间 5 种植被恢复模式的生物量均呈波动增长趋势(图2);且相对于自然恢复模式灌草坡,4 种人工林均可有效加速植被的恢复进程。纯桉林在生长初期生物量累积方面的优势更为明显,在 4 龄时显著高于同龄的 30 种乡土树种混交林与红锥纯林。有研究表明,以桉树为代表的速生先锋种在酸性或不肥沃土壤上具有高度适应性,可在人为启动后快速恢复地上生物量[28],但随着林龄的增加具有明显的自疏现象,这将导致其生物量在后期发展过程中增长放缓,甚至可能出现负增长[29];而相比纯林,混交林也可产生相似或更高的生物量[30- 31]。本研究中外来先锋种尾叶桉的速生性使其生物量在 6 龄时已显著高于其自身 1 龄时的水平,但后劲不足,在 6—13 龄间增长速度放缓;相反,乡土种人工林生物量增长表现出前慢后快的特征,且两个混交林优于红锥纯林(图2)。到 13 龄时,两个乡土树种混交林生物量已与外来种桉树纯林无显著差异,且 10NS 已超越 EU(图3),由此说明多树种混交有助于提高人工林发展后期的及生物量。
大量研究表明,混交林营造被认为是加速生物多样性、系统稳定性恢复的有效手段[32- 35]。与前人的研究结果一致,本研究发现 5 种恢复模式在 13 年间植物群落物种多样性整体表现为混交林>纯林>对照灌草坡的规律(图6,图7)。但是,与生物量的正向发展趋势不同,5 种恢复模式植物群落物种多样性(物种丰富度、Shannon-Wiener 指数、Pielou 均匀度指数)在研究期间均呈波动下降趋势。
1 龄时,30 NS 物种多样性在所有人工林最高,但发展到13 龄时,30NS物种多样性不仅显著低于其在 1 龄时的水平(图4,图5),而且在 Shannon-Wiener 指数、均匀度指数方面略低于10NS;除此以外,10NS 的 Shannon-Wiener 指数、Pielou 均匀度指数均显著高于 SH,且其 Pielou 均匀度显著高于 EU(图7)。Yu 等在广东鹤山的长期定位研究发现,除物种丰富度外,遗传多样性也与混交林群落生物量稳定性显著正相关;而且优势种的特性对于系统的稳定性有着重要影响[35]。因此,营造乡土树种混交林是加速植被恢复的有效措施,但并不是起始物种丰富度越高越好[36],混交林遗传多样性和优势树种的选取亦是需要谨慎考虑的问题。
地上生物量与地下土壤库的互作与非同步发展是植被恢复与演替过程中的常态[9,37- 38]。有研究发现,20 龄时南亚热带自然恢复灌草坡土壤中的 SOC、水解氮和有效钾等营养元素已基本恢复到同地带顶级森林群落的水平,而地上植被仍处于草本和灌丛占优势的低生物多样性阶段[39]。另外,在长时期尺度上混交乡土人工林的土壤全碳、全氮、全磷以及速效磷含量显著高于纯桉林与裸荒地[40]。本研究结果显示,研究期间 5 种恢复模式土壤 pH、SMC、SOC、TN、C/N 均呈波动上升趋势,至 13 龄时 5 种恢复模式的 SMC,两个混交林的 SOC 和 pH,自然恢复灌草坡的土壤 pH,以及红锥纯林的土壤 C/N 均已显著高于其各自 1 龄时的水平(图12)。而 5 种恢复模式相比,13 龄时,只有 10NS土壤全氮显著高于 CM 与 SH,5 种植被恢复模式的土壤 pH、SMC、SOC 与土壤 C/N 均无显著差异(图12)。RDA 结果显示,研究期间土壤理化性状对地上植被结构与健康水平的解释度由 1 龄时的 73.3% 上升至 13 龄时的 82.0%;13 龄时,只有 SOC 与地上植物群落结构与生物热力学健康指标间的相关关系已达到显著水平(图13)。综合上述研究结果,本研究认为植被与土壤间互作关系的建立是一个长期的过程;植物群落结构改变对土壤理化性状的差异性影响难以短期内有所显现。
5 结论
研究量化分析了 3 种乡土树种人工林植物群落结构、健康水平与土壤理化性状在 1—13 龄间的发展规律,并与外来种桉树人工纯林与自然恢复灌草坡进行对比,旨在为粤港澳大湾区退化生态系统的修复及管理提出建议。主要结论如下:
(1)1—13 龄间,3 种乡土树种人工林与尾叶桉纯林和自然恢复灌草坡的植物群落结构、生物热力学健康及植被与土壤间的关系均遵循相似的动态规律,但在节率上有所差异。研究期间,5 种恢复模式植物群落的生物量和生态呈波动上升趋势,但物种多样性与结构则呈(波动)下降趋势,表明各群落发展进入物种更新的瓶颈期,应考虑在林分改造中适当引入遗传信息含量较高的森林演替中晚期物种,促进群落的正向演替与发展。
(2)相对而言,混交林在生物量、物种多样性和群落生物热力学健康水平的恢复方面均优于纯林,而纯林又优于自然恢复灌草坡;但乡土树种人工林起始物种丰富度并不是越高越好。13 龄时,10 种乡土树种混交林的生物量、群落多样性水平(Shannon-Wiener 指数与 Pielou 均匀度指数)以及生物热力学健康水平(生态)和土壤总氮含量均优于 30 种乡土树种混交林。
(3)植被与土壤间互作关系的建立是一个长期的过程;人工林恢复模式可有效加速植物群落的发展,但对土壤理化性状的差异性影响难以短期内有所显现。研究期间植被与土壤间的相关关系逐步建立,但13 龄时,只有土壤有机碳含量与地上植被结构与生物热力学健康指标建立起了显著的相关关系。除了 13 龄时 10 种乡土树种混交林的土壤全氮含量显著高于红锥纯林与自然恢复灌草坡外,整个研究期间,5 种恢复模式间的土壤 pH、含水量、有机碳与碳氮比均无显著差异。
